

Tracking the Spatio-Temporal Distribution of Organic Particles to Predict Macroaggregation in the Northern Adriatic Sea



Abstract
:1. Introduction
2. Materials and Methods
2.1. Seawater Sampling
2.2. Oceanographic Measurements
2.3. Model Particles
2.4. Electrochemical Method
2.5. Electrochemical Measurements
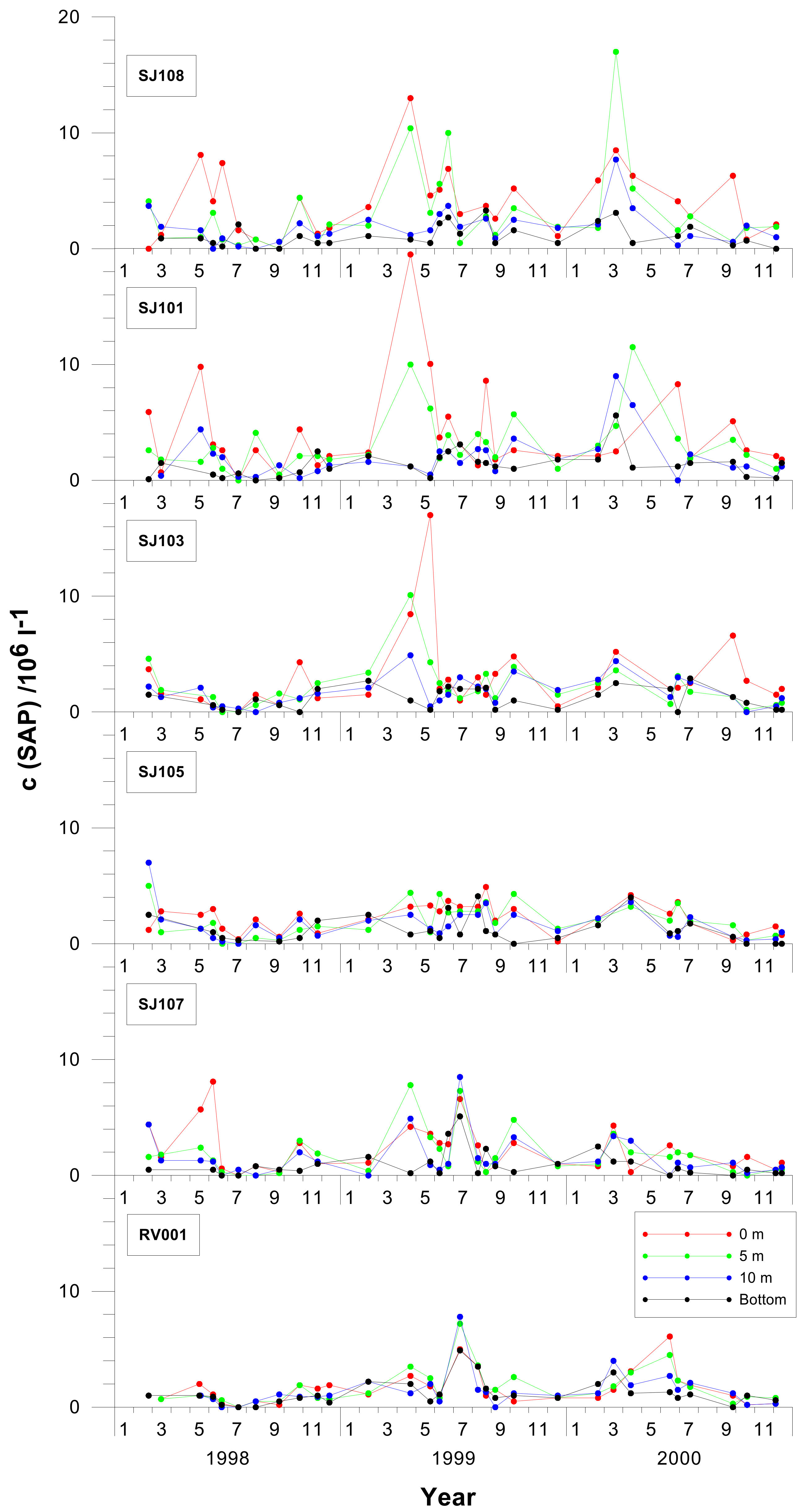
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Templado, J. Future Trends of Mediterranean Biodiversity. In The Mediterranean Sea, 1st ed.; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 1, pp. 479–498. [Google Scholar] [CrossRef]
- Marbá, N.; Jordá, G.; Agustí, S.; Girard, C.; Duarte, C.M. Footprints of Climate Change on Mediterranean Sea Biota. Front. Mar. Sci. 2015, 2, 56. [Google Scholar] [CrossRef]
- Schroeder, K.; Chiggiato, J.; Bryden, H.L.; Borghini, M.; Ben Ismail, S. Abrupt Climate Shift in the Western Mediterranean Sea. Sci. Rep. 2016, 6, 23009. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Yáñez, M.; García, M.J.; Salat, J.; García-Martínez, M.; Pascual, J.; Moya, F. Warming Trends and Decadal Variability in the Western Mediterranean Shelf. Glob. Planet Chang. 2008, 63, 177–184. [Google Scholar] [CrossRef]
- Budiša, A.; Paliaga, P.; Juretić, T.; Lučić, D.; Supić, N.; Pasarić, Z.; Đakovac, T.; Mladinić, M.; Dadić, V.; Tičina, V. Distribution, diet and relationships of the invasive ctenophore Mnemiopsis leidyi with anchovies and zooplankton, in the northeastern Adriatic Sea. Medit. Mar. Sci. 2021, 22, 827–842. [Google Scholar] [CrossRef]
- Savun-Hekimoğlu, B.; Gazioğlu, C. Mucilage Problem in the Semi-Enclosed Seas: Recent outburst in the Sea of Marmara. Int. J. Environ. Geoinform. 2021, 8, 402–413. [Google Scholar] [CrossRef]
- Aktan, Y.; Dede, A.; Ciftci, P.S. Mucilage Event Associated with Diatoms and Dinoflagellates in Sea of Marmara, Turkey. In Harmful Algae News; The Intergovernmental Oceanographic Commission of UNESCO; UNESCO: Paris, France, 2008; Volume 36, pp. 1–3. [Google Scholar]
- Tüfekçi, V.; Balkıs, N.; Polat Beken, Ç.; Ediger, D.; Mantıkçı, M. Phytoplankton Composition and Environmental Conditions of a Mucilage Event in the Sea of Marmara. Turk. J. Biol. 2010, 34, 199–210. [Google Scholar] [CrossRef]
- İşinibilir Okyar, M.; Üstün, F.; Orun, D.A. Changes in Abundance and Community Structure of the Zooplankton Population during the 2008 Mucilage Event in the Northeastern Marmara Sea. Turk. J. Zool. 2015, 39, 28–38. [Google Scholar] [CrossRef]
- Özalp, H.B. First Massive Mucilage Event Observed in Deep Waters of Çanakkale Strait (Dardanelles), Turkey. J. Black Sea Mediterr. Environ. 2021, 27, 49–66. [Google Scholar]
- Genitsaris, S.; Stefanidou, N.; Sommer, U.; Moustaka-Gouni, M. Phytoplankton blooms, red tides and mucilaginous aggregates in the urban Thessaloniki Bay, Eastern Mediterranean. Diversity 2019, 11, 136. [Google Scholar] [CrossRef]
- Danovaro, R.; Fonda Umani, S.; Pusceddu, A.M. Climate Change and the Potential Spreading of Marine Mucilage and Microbial Pathogens in the Mediterranean Sea. PLoS ONE 2009, 4, e7006. [Google Scholar] [CrossRef]
- Rouaud, V.; Susperregui, N.; Fahy, A.; Guyoneaud, R.; Bichon, S.; Lienart, C.; Del Amo, Y.; Savoye, N.; Gaudin, P.; Duran, R.; et al. Dynamics of microbial communities across the three domains of life over an annual cycle with emphasis on marine mucilage in the Southern Bay of Biscay resolved by microbial fingerprinting. Cont. Shelf Res. 2019, 186, 127–137. [Google Scholar] [CrossRef]
- Karadurmuş, U.; Sari, M. Marine mucilage in the Sea of Marmara and Its Effects on the Marine Ecosystem: Mass Deaths. Turk. J. Zool. 2022, 46, 93–102. [Google Scholar] [CrossRef]
- Degobbis, D.; Malej, A.; Fonda-Umani, S. The mucilage phenomenon in the northern Adriatic: A critical review of the present scientific hypotheses. Ann. Ist. Super Sanità 1999, 35, 373–381. [Google Scholar]
- Precali, R.; Giani, M.; Marini, M.; Grilli, F.; Ferrari, C.R.; Pecar, O.; Paschini, E. Mucilaginous aggregates in the northern Adriatic in the period 1999-2002: Typology and distribution. Sci. Total Environ. 2005, 353, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Tejedor, M.F.; Ismael, A.; Montresor, M.; Mozetič, P.; Taş, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2021, 102, 101843. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, R.A.; Rinaldi, A. (Eds.) Marine Mucilages. Special issue. Sci. Total Environ. 1995, 165, 1–235. [Google Scholar]
- Funari, E.; Azam, F.; Fonda Umani, S.; Pagnotta, R. (Eds.) State of the art and new scientific hypotheses on the phenomenon of mucilages in the Adriatic Sea. Special issue. Ann. Ist. Super Sanità 1999, 35, 353–476. [Google Scholar]
- Žutić, V.; Svetličić, V. Interfacial processes. In The Handbook of Environmental Chemistry; Wangersky, P., Ed.; Part D. Marine Chemistry; Springer-Verlag: Berlin/Heidelberg, Germany, 2000; Volume 5, pp. 149–165. [Google Scholar]
- Giani, M.; Degobbis, D.; Rinaldi, A. (Eds.) Mucilages in the Adriatic and Tyrrhenian Seas. Special issue. Sci. Total Environ. 2005, 353, 1–380. [Google Scholar] [CrossRef]
- Svetličić, V.; Žutić, V.; Zimmermann, A.H. Biophysical scenario of giant gel formation in the Northern Adriatic Sea. Ann. N. Y. Acad. Sci. 2005, 1048, 524–527. [Google Scholar] [CrossRef]
- Svetličić, V.; Balnois, E.; Žutić, V.; Chevalet, J.; Hozić Zimmermann, A.; Kovač, S.; Vdović, N. Electrochemical detection of gel microparticles in seawater. Croat. Chem. Acta 2006, 79, 107–113. [Google Scholar]
- Pletikapić, G.; Ivošević DeNardis, N. Application of surface analytical methods for hazardous situation in the Adriatic Sea: Monitoring of organic matter dynamics and oil pollution. Nat. Hazards Earth Syst. Sci. 2017, 17, 31–44. [Google Scholar] [CrossRef]
- Novosel, N.; Mišić Radić, T.; Zemla, J.; Lekka, M.; Čačković, A.; Kasum, D.; Legović, T.; Žutinić, P.; Gligora Udovič, M.; Ivošević DeNardis, N. Temperature-induced response in algal cell surface properties and behaviour: An experimental approach. J. Appl. Phycol. 2022, 34, 243–259. [Google Scholar] [CrossRef]
- Novosel, N.; Mišić Radić, T.; Levak Zorinc, M.; Zemla, J.; Lekka, M.; Vrana, I.; Gašparović, B.; Horvat, L.; Kasum, D.; Legović, T.; et al. Salinity induced chemical, mechanical and behavioral changes in marine microalgae. J. Appl. Phycol. 2022, 34, 1293–1309. [Google Scholar] [CrossRef]
- Pletikapić, G.; Mišić Radić, T.; Hozić Zimmermann, A.; Svetličić, V.; Pfannkuchen, M.; Marić, D.; Godrijan, J.; Žutić, V. AFM imaging of extracellular polymer release by marine diatom Cylindrotheca closterium (Ehrenberg) Reiman & J.C. Lewin. J. Mol. Recognit. 2011, 24, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Novosel, N.; Ivošević DeNardis, N. Structural Features of the Algal Cell Determine Adhesion Behavior at a Charged Interface. Electroanal 2021, 33, 1436–1443. [Google Scholar] [CrossRef]
- Žutić, V.; Svetličić, V.; Ivošević, N.; Hozić, A.; Pečar, O. Northern Adriatic mesocosm experiment Rovinj 2003: Dynamics of organic microparticles studied by the electrochemical technique. Period Biol. 2004, 106, 67–74. [Google Scholar]
- Ivošević DeNardis, N.; Šegota, S.; Svetličić, V.; Castelli, A.; Kljajić, Z. Characterization of marine organic matter in the bay of Boka Kotorska by electrochemical and atomic force microscopy imaging. Stud. Mar. 2013, 26, 5–21. [Google Scholar]
- Kraus, R.; Supić, N.; Precali, R. Factors favoring phytoplankton blooms in the northern Adriatic: Towards the northern Adriatic empirical ecological model. Ocean Sci. 2016, 12, 19–37. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis, 2nd ed.; Pergamon Press: New York, NY, USA, 1985; pp. 1–173. [Google Scholar]
- Utermöhl, H. Zur Verfollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Verein. Theor. Angew. Limnol. 1958, 17, 47–71. [Google Scholar]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrates Animals; Smith, M.L., Chanley, M.H., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar] [CrossRef]
- Ivošević, N.; Žutić, V. Polarography of marine particles: A model study. Croat. Chem. Acta 1997, 70, 167–178. [Google Scholar]
- Baldi, F.; Ivošević, N.; Minacci, A.; Pepi, M.; Fani, R.; Svetličić, V.; Žutić, V. Adhesion of Acinetobacter venetianus to diesel fuel roplets studied with in situ electrochemical and molecular probes. Appl. Environ. Microbiol. 1999, 65, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Žutić, V.; Ivošević, N.; Svetličić, V.; Long, R.A.; Azam, F. Film formation by marine bacteria at a model fluid interface. Aquat. Microb. Ecol. 1999, 17, 231–238. [Google Scholar] [CrossRef]
- Kovač, S.; Kraus, R.; Geček, S.; Žutić, V. Cell suspension as a model system for electrochemical analysis. Croat. Chem. Acta 2000, 73, 279–291. [Google Scholar]
- Poulain, P.M.; Kourafalou, V.H.; Cushman-Roisin, B. Northern Adriatic Sea. In Physical Oceanography of the Adriatic Sea; Cushman-Roisin, B., Gačić, M., Poulain, P.M., Artegiani, A., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 143–165. [Google Scholar] [CrossRef]
- Supić, N.; Orlić, M.; Degobbis, D. Istrian Coastal Countercurrent and its Year-to-Year Variability. Estuar. Coast. Shelf Sci. 2000, 51, 385–397. [Google Scholar] [CrossRef]
- Supić, N.; Kraus, R.; Kuzmić, M.; Paschini, E.; Precali, R.; Russo, A.; Vilibić, I. Predictability of northern Adriatic winter conditions. J. Mar. Syst. 2012, 90, 42–57. [Google Scholar] [CrossRef]
- Sournia, A. La production primaire planctonique en Mediterranée. Essai de mise a jour. Bull Etude Comm. Méditerr. Num. Spéc. 1973, 5, 1–128. [Google Scholar]
- Fonda Umani, S.; Milani, L.; Borme, D.; de Olazabal, A.; Parlato, S.; Precali, R.; Kraus, R.; Lucić, D.; Njire, J.; Totti, C.; et al. Inter-annual variations of planktonic food webs in the northern Adriatic Sea. Sci. Total Environ. 2005, 353, 218–231. [Google Scholar] [CrossRef]
- Fonda Umani, S.; Del Negro, P.; Larato, C.; De Vittor, C.; Cabrini, M.; Celio, M.; Falconi, C.; Tamberlich, F.; Azam, F. Major inter-annual variations in microbial dynamics in the Gulf of Trieste (northern Adriatic Sea) and their ecosystem implications. Aquat. Microb. Ecol. 2007, 46, 163–175. [Google Scholar] [CrossRef]
- Solidoro, C.; Bastianini, M.; Bandelj, V.; Codermatz, R.; Cossarini, G.; Canu, D.M.; Ravagnan, E.; Salon, S.; Trevisani, S. Current state, scales of variability, and trends of biogeochemical properties in the northern Adriatic Sea. J. Geophys. Res. 2009, 114, 7–91. [Google Scholar] [CrossRef]
- Mozetič, P.; Solidoro, C.; Cossarini, G.; Socal, G.; Precali, R.; Francé, J.; Bianchi, F.; Vittor, C.D.; Smodlaka, N.; Fonda Umani, S. Recent Trends Towards Oligotrophication of the Northern Adriatic: Evidence from Chlorophyll a Time Series. Estuaries Coast. 2010, 33, 362–375. [Google Scholar] [CrossRef]
- Totti, C.; Cangini, M.; Ferrari, C.; Kraus, R.; Pompei, M.; Pugnetti, A.; Romagnoli, T.; Vanucci, S.; Socal, G. Phytoplankton size-distribution and community structure in relation to mucilage occurrence in the northern Adriatic Sea. Sci. Total Environ. 2005, 353, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Varezić Bojanić, D.B.; Vidjak, O.; Kraus, R.; Precali, R. Regulating mechanisms of calanoid copepods variability in the northern Adriatic Sea: Testing the roles of west-east salinity and phytoplankton gradients. Estuar. Coast. Shelf Sci. 2015, 164, 288–300. [Google Scholar] [CrossRef]
- Dautović, J.; Vojvodić, V.; Tepić, N.; Ćosović, B.; Ciglenečki, I. Dissolved organic carbon as potential indicator of global change: A long-term investigation in the northern Adriatic. Sci. Total Environ. 2017, 587–588, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Ciglenečki, I.; Vilibić, I.; Dautović, J.; Vojvodić, V.; Ćosović, B.; Zemunik, P.; Dunić, N.; Mihanović, H. Dissolved organic carbon and surface active substances in the northern Adriatic Sea: Long-term trends, variability and drivers. Sci. Total Environ. 2020, 730, 139104. [Google Scholar] [CrossRef] [PubMed]
- Gašparović, B.; Ćosović, B. Surface-active properties of organic matter in the North Adriatic Sea. Estuar. Coast. Shelf Sci. 2003, 58, 555–566. [Google Scholar] [CrossRef]
- Revelante, N.; Gilmartin, M. Temporal succession of phytoplankton in the northern Adriatic. Neth. J. Sea Res. 1976, 10, 377–396. [Google Scholar] [CrossRef]
- Revelante, N.; Gilmartin, M. The lateral advection of particulate organic matter from the Po Delta region during summer stratification, and its implications for the northern Adriatic. Estuar. Coast. Shelf Sci. 1992, 35, 191–212. [Google Scholar] [CrossRef]
- Kraus, R.; Supić, N. Impact of circulation on high phytoplankton blooms and fish catch in the northern Adriatic (1990–2004). Estuar. Coast. Shelf Sci. 2011, 91, 198–210. [Google Scholar] [CrossRef]
- Marić, D.; Kraus, R.; Godrijan, J.; Supić, N.; Đakovac, T.; Precali, R. Phytoplankton response to climatic and anthropogenic influences in the north-eastern Adriatic during the last four decades. Estuar. Coast. Shelf Sci. 2012, 115, 98–112. [Google Scholar] [CrossRef]
- Kraus, R.; Supić, N.; Lučić, D.; Njire, J. Impact of winter oceanographic conditions on zooplankton abundance in northern Adriatic with implications on Adriatic anchovy stock prognosis. Estuar. Coast. Shelf Sci. 2015, 167, 56–66. [Google Scholar] [CrossRef]
- Russo, A.; Maccaferri, S.; Djakovac, T.; Precali, R.; Degobbis, D.; Deserti, M.; Paschini, E.; Lyons, D.M. Meteorological and oceanographic conditions in the northern Adriatic Sea during the period June 1999–July 2002: Influence on the mucilage phenomenon. Sci. Total Environ. 2005, 353, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Degobbis, D.; Precali, R.; Ferrari, C.R.; Djakovac, T.; Rinaldi, A.; Ivančić, I.; Gismondi, M.; Smodlaka, N. Changes in nutrient concentrations and ratios during mucilage events in the period 1999–2002. Sci. Total Environ. 2005, 353, 103–114. [Google Scholar] [CrossRef]
- Volf, G.; Atanasova, N.; Kompare, B.; Precali, R.; Ožanić, N. Study of the impact of TIN/PO4 ratio on mucilage formation in the northern Adriatic using regression trees. Acta Adriat. 2015, 56, 207–222. [Google Scholar]
- Kraus, R.; Supić, N. Sea dynamics impacts on the macroaggregates: A case study of the 1997 mucilage event in the northern Adriatic. Prog. Oceanogr. 2015, 138, 249–267. [Google Scholar] [CrossRef]
- Alldredge, A.L. The potential role of particulate diatom exudates in forming nuisance mucilaginous scums. Ann. Ist. Super Sanità 1999, 35, 397–400. [Google Scholar]
- Myklestad, S.M. Phytoplankton extracellular production and leakage with consideration on the polysaccharide accumulation. Ann. Ist. Super Sanità 1999, 35, 411–419. [Google Scholar]
- Kraus, R.; Ivančić, I.; Đakovac, T.; Smodlaka, N. Northern Adriatic mesocosm experiment Rovinj 2003: Changes in the microphytoplankton community. Period Biol. 2004, 106, 31–37. [Google Scholar]
- Najdek, M.; Blažina, M.; Đakovac, T.; Kraus, R. The role of the diatom Cylindrotheca closterium in a mucilage event in the northern Adriatic Sea: Coupling with high salinity water intrusions. J. Plankton Res. 2005, 27, 851–862. [Google Scholar] [CrossRef]
- Degobbis, D.; Fonda-Umani, S.; Franco, P.; Malej, A.; Precali, R.; Smodlaka, N. Changes in the northern Adriatic ecosystem and the hypertrophic appearance of gelatinous aggregates. Sci. Total Environ. 1995, 165, 43–58. [Google Scholar] [CrossRef]
- Grilli, F.; Paschini, E.; Precali, R.; Russo, A.; Supić, N. Circulation and horizontal fluxes in the northern Adriatic Sea in the period June 1999–July 2002. Part I: Geostrophic circulation and current measurement. Sci. Total Environ. 2005, 353, 57–67. [Google Scholar] [CrossRef]
- Giani, M.; Savelli, F.; Berto, D.; Zangrando, V.; Ćosović, B.; Vojvodić, V. Temporal dynamics of dissolved and particulate organic carbon in the northern Adriatic Sea in relation to the mucilage events. Sci. Total Environ. 2005, 353, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Ćosović, B.; Vojvodić, V. Mucilage events in the Northern Adriatic: Search for tools for early warning. Period Biol. 2000, 102, 255–259. [Google Scholar]
- Pettine, M.; Capri, S.; Manganelli, M.; Patrolecco, L.; Puddu, A.; Zoppini, A. The dynamics of DOM in the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2001, 52, 471–489. [Google Scholar] [CrossRef]
- Giani, M.; Berto, D.; Zangrando, V.; Castelli, S.; Sist, P.; Urbani, R. Chemical characterization of different typologies of mucilaginous aggregates in the Northern Adriatic Sea. Sci. Total Environ. 2005, 353, 232–246. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.G.; Lewin, J.C.; Johnson, P. Marine Polymers. IV Diatom Polysaccharides. Bot. Mar. 1972, 15, 102–108. [Google Scholar] [CrossRef]
- Haug, A.; Myklestad, S.M. Polysaccharides of marine diatoms with special reference to Chaetoceros species. Mar. Biol. 1976, 34, 217–222. [Google Scholar] [CrossRef]
- Urbani, R.; Magaletti, E.; Sist, P.; Cicero, A.M. Extracellular carbohydrates released by the marine diatoms Cylindrotheca closterium, Thalassiosira pseudonana and Skeletonema costatum: Effect of P-depletion and growth status. Sci. Total Environ. 2005, 353, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Marty, J.C.; Žutić, V.; Precali, R.; Saliot, A.; Ćosović, B.; Smodlaka, N.; Cauwet, G. Organic matter characterization in the Northern Adriatic sea with special reference to the sea surface microlayer. Mar. Chem. 1988, 25, 243–263. [Google Scholar] [CrossRef]
- Malfatti, F.; Turk, V.; Tinta, T.; Mozetič, P.; Manganelli, M.; Samoa, T.J.; Ugalde, J.A.; Kovač, N.; Stefanelli, M.; Antonioli, M.; et al. Microbial mechanisms coupling carbon and phosphorus cycles in phosphorus-limited northern Adriatic Sea. Sci. Total Environ. 2014, 470/471, 1173–1183. [Google Scholar] [CrossRef]
- Totti, C.; Cucchiari, E.M.; Romagnoli, T. Intra and interannual variability of phytoplankton in coastal area of Senigallia (northern Adriatic Sea) from 1988 to 2000. Biol. Mar. Mediterr. 2002, 9, 391–399. [Google Scholar]
- Bernardi Aubry, F.; Berton, A.; Bastianini, M.; Socal, G.; Acri, F. Phytoplankton succession in a coastal area of the NW Adriatic, over a 10-year sampling period (1990–1999). Cont. Shelf Res. 2004, 24, 97–115. [Google Scholar] [CrossRef]
- Cerino, F.; Fornasaro, D.; Kralj, M.; Giani, M.; Cabrini, M. Phytoplankton temporal dynamics in the coastal waters of the north-eastern Adriatic Sea (Mediterranean Sea) from 2010 to 2017. Nat. Conserv. 2019, 34, 343–372. [Google Scholar] [CrossRef]
- Totti, C.; Romagnoli, T.; Accoroni, S.; Coluccelli, A.; Pellegrini, M.O.; Campanelli, A.; Grilli, F.; Marini, M. Phytoplankton communities in the northwestern Adriatic Sea: Interdecadal variability over a 30-years period (1988–2016) and relationships with meteoclimatic drivers. J. Mar. Syst. 2019, 193, 137–153. [Google Scholar] [CrossRef]
- Albay, M. Marmara Denizi’nin Müsilaj Sorunu (The Mucilage Problem of the Marmara Sea, in Turkish), 1st ed.; Istanbul University Press: Istanbul, Turkey, 2023; pp. 1–405. [Google Scholar] [CrossRef]
- Yagci, A.L.; Colkesen, I.; Kavzoglu, T.; Sefercik, U.G. Daily monitoring of marine mucilage using the MODIS products: A case study of 2021 mucilage bloom in the Sea of Marmara, Turkey. Environ. Monit. Assess 2022, 194, 170. [Google Scholar] [CrossRef] [PubMed]
- Kavzoglu, T.; Goral, M. Google Earth Engine for Monitoring Marine Mucilage: Izmit Bay in Spring 2021. Hydrology 2022, 9, 135. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Li, L.; Hussain, S.; Lee, J.L.; Mumtaz, F.; Elbeltagi, A.; Waqas, M.S.; Dilawar, A. Analysis of Seasonal Variations in Surface Water Quality over Wet and Dry Regions. Water 2022, 14, 1058. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Hussain, S.; Cheema, M.J.M.; Lee, J.L.; Waqas, M.S.; Aslam, M.A. Seasonal effect of agricultural pollutants on coastline environment: A case study of the southern estuarine water ecosystem of the boseong county Korea. Pak. J. Agric. Sci. 2022, 59, 117–124. [Google Scholar] [CrossRef]
- Rizvi, F.F.; Ahmed, B.; Hussain, S.; Khan, A.; Raza, M.; Shahid, M. Assessment of climate extremes from historical data (1960-2013) of Soan River Basin in Pakistan. Int. J. Glob. Warm. 2021, 25, 1–37. [Google Scholar] [CrossRef]
- Fitzenreiter, K.; Mao, M.; Xia, M. Characteristics of Surface Currents in a Shallow Lagoon–Inlet–Coastal Ocean System Revealed by Surface Drifter Observations. Estuaries Coasts 2022, 45, 2327–2344. [Google Scholar] [CrossRef]
- Jiang, L.; Xia, M. Modeling investigation of the nutrient and phytoplankton variability in the Chesapeake Bay outflow plume. Prog. Oceanogr. 2018, 162, 290–302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Date | Depth | Species | Species Abundance | Total Abundance | Species Contribution | Cumulative (%) |
|---|---|---|---|---|---|---|---|
| SJ101 | 25 Feb 1998 | 0 | Skeletonema marinoi | 748,140 | 1,146,260 | 65.3 | 65.3 |
| Nitzschia delicatissima complex | 290,820 | 25.4 | 90.6 | ||||
| 5 | Nitzschia delicatissima complex | 928,700 | 1,735,300 | 53.5 | 53.5 | ||
| Skeletonema marinoi | 715,580 | 41.2 | 94.8 | ||||
| 10 | Nitzschia delicatissima complex | 583,120 | 768,120 | 75.9 | 75.9 | ||
| Skeletonema marinoi | 170,200 | 22.2 | 98.1 | ||||
| SJ101 | 17 Mar 1998 | 0 | Nitzschia delicatissima complex | 59,570 | 61,050 | 97.6 | 97.6 |
| 5 | Nitzschia delicatissima complex | 39,590 | 41,810 | 94.7 | 94.7 | ||
| 10 | Nitzschia delicatissima complex | 38,110 | 39,590 | 96.3 | 96.3 | ||
| SJ101 | 15 Feb 1999 | 0 | Skeletonema marinoi | 85,470 | 125,800 | 67.9 | 67.9 |
| Nitzschia delicatissima complex | 28,120 | 22.4 | 90.3 | ||||
| 5 | Skeletonema marinoi | 87,690 | 128,020 | 68.5 | 68.5 | ||
| Nitzschia delicatissima complex | 29,970 | 23.4 | 91.9 | ||||
| 10 | Skeletonema marinoi | 67,340 | 105,450 | 63.9 | 63.9 | ||
| Nitzschia delicatissima complex | 29,600 | 28.1 | 91.9 | ||||
| SJ101 | 24 Apr 1999 | 0 | Skeletonema marinoi | 6,926,400 | 15,258,800 | 45.4 | 45.4 |
| Nitzschia delicatissima complex | 5,220,700 | 34.2 | 79.6 | ||||
| Chaetoceros sp. | 1,713,100 | 11.2 | 90.8 | ||||
| 5 | Nitzschia delicatissima complex | 657,860 | 990,120 | 66.4 | 66.4 | ||
| Skeletonema marinoi | 227,920 | 23.0 | 89.5 | ||||
| 10 | Skeletonema marinoi | 14,430 | 31,080 | 46.4 | 46.4 | ||
| Nitzschia delicatissima complex | 13,690 | 44.0 | 90.5 | ||||
| SJ101 | 21 Feb 2000 | 0 | Skeletonema marinoi | 1,266,880 | 1,381,580 | 91.7 | 91.7 |
| 10 | Skeletonema marinoi | 201,280 | 248,640 | 81.0 | 81.0 | ||
| SJ101 | 21 Mar 2000 | 0 | Chaetoceros sp. | 10,730 | 21,460 | 50.0 | 50.0 |
| Skeletonema marinoi | 4440 | 20.7 | 70.7 | ||||
| 5 | Chaetoceros sp. | 14,060 | 38,110 | 36.9 | 36.9 | ||
| Skeletonema marinoi | 8510 | 22.3 | 59.2 | ||||
| Nitzschia delicatissima complex | 7400 | 19.4 | 78.6 | ||||
| Chaetoceros curvisetus | 6660 | 17.5 | 96.1 | ||||
| 10 | Nitzschia delicatissima complex | 29,230 | 48,840 | 59.8 | 59.8 | ||
| Skeletonema marinoi | 10,360 | 21.2 | 81.1 | ||||
| Chaetoceros curvisetus | 5180 | 10.6 | 91.7 | ||||
| SJ101 | 17 Apr 2000 | 0 | Nitzschia delicatissima complex | 102,490 | 113,960 | 89.9 | 89.9 |
| 5 | Nitzschia delicatissima complex | 156,140 | 163,540 | 95.5 | 95.5 | ||
| 10 | Nitzschia delicatissima complex | 33,670 | 39,220 | 85.8 | 85.8 |
| Station | Date | Depth | Species | Species Abundance | Sample Abundance | Species Contribution | Cumulative (%) |
|---|---|---|---|---|---|---|---|
| SJ107 | 25 Feb 1998 | 0 | Nitzschia delicatissima complex | 93,240 | 124,320 | 75.0 | 75.0 |
| Rhizosolenia fragilissima | 13,320 | 10.7 | 85.7 | ||||
| 5 | Nitzschia delicatissima complex | 386,280 | 481,000 | 80.3 | 80.3 | ||
| Rhizosolenia fragilissima | 86,580 | 18.0 | 98.3 | ||||
| 10 | Nitzschia delicatissima complex | 123,210 | 140,970 | 87.4 | 87.4 | ||
| SJ107 | 17 Mar 1998 | 0 | Nitzschia delicatissima complex | 123,580 | 127,280 | 97.1 | 97.1 |
| 5 | Nitzschia delicatissima complex | 66,970 | 71,780 | 93.3 | 93.3 | ||
| 10 | Nitzschia delicatissima complex | 94,720 | 99,160 | 95.5 | 95.5 | ||
| SJ107 | 15 Feb 1999 | 0 | Skeletonema marinoi | 3700 | 12,210 | 30.3 | 30.3 |
| Chaetoceros affinis | 1480 | 12.1 | 42.4 | ||||
| Chaetoceros curvisetus | 1480 | 12.1 | 54.5 | ||||
| Nitzschia tenuirostris | 1480 | 12.1 | 66.7 | ||||
| 5 | Skeletonema marinoi | 8880 | 13,320 | 66.7 | 66.7 | ||
| Nitzschia tenuirostris | 2220 | 16.7 | 83.3 | ||||
| 10 | Skeletonema marinoi | 14,060 | 17,390 | 80.9 | 80.9 | ||
| SJ107 | 24 Apr 1999 | 0 | Chaetoceros sp. | 364,450 | 836,200 | 43.6 | 43.6 |
| Nitzschia delicatissima complex | 223,850 | 26.8 | 70.4 | ||||
| 5 | Nitzschia delicatissima complex | 246,420 | 695,600 | 35.4 | 35.4 | ||
| Skeletonema marinoi | 155,400 | 22.3 | 57.8 | ||||
| Chaetoceros socialis | 138,380 | 19.9 | 77.7 | ||||
| Chaetoceros curvisetus | 76,220 | 11.0 | 88.6 | ||||
| 10 | Nitzschia delicatissima complex | 109,520 | 204,240 | 53.6 | 53.6 | ||
| Skeletonema marinoi | 47,360 | 23.2 | 76.8 | ||||
| SJ107 | 21 Feb 2000 | 0 | Chaetoceros curvisetus | 492 | 1476 | 33.3 | 33.3 |
| Nitzschia delicatissima complex | 246 | 16.7 | 50.0 | ||||
| Nitzschia tenuirostris | 246 | 16.7 | 66.7 | ||||
| Prorocentrum micans | 246 | 16.7 | 83.3 | ||||
| 5 | Nitzschia tenuirostris | 555 | 1850 | 30.0 | 30.0 | ||
| Melosira dubia | 370 | 20.0 | 50.0 | ||||
| Nitzschia delicatissima complex | 370 | 20.0 | 70.0 | ||||
| Achnanthes sp. | 185 | 10.0 | 80.0 | ||||
| 10 | Nitzschia tenuirostris | 246 | 861 | 28.6 | 28.6 | ||
| Achnanthes sp. | 123 | 14.3 | 42.9 | ||||
| Mastogloia sp. | 123 | 14.3 | 57.1 | ||||
| Navicula sp. | 123 | 14.3 | 71.4 | ||||
| Nitzschia delicatissima complex | 123 | 14.3 | 85.7 | ||||
| SJ107 | 17 Mar 2000 | 0 | Skeletonema marinoi | 75,110 | 105,080 | 71.5 | 71.5 |
| Chaetoceros sp. | 17,760 | 16.9 | 88.4 | ||||
| 5 | Skeletonema marinoi | 81,770 | 92,500 | 88.4 | 88.4 | ||
| 10 | Skeletonema marinoi | 36,260 | 45,140 | 80.3 | 80.3 | ||
| SJ107 | 14 Apr 2000 | 0 | Nitzschia delicatissima complex | 555 | 1110 | 50.0 | 50.0 |
| Prorocentrum micans | 370 | 33.3 | 83.3 | ||||
| 5 | Nitzschia delicatissima complex | 1353 | 1353 | 100.0 | 100.0 | ||
| 10 | Nitzschia delicatissima complex | 1110 | 1665 | 66.7 | 66.7 | ||
| Glenodinium sp. | 185 | 11.1 | 77.8 | ||||
| Gymnodinium sp. | 185 | 11.1 | 88.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraus, R.; Ivošević DeNardis, N. Tracking the Spatio-Temporal Distribution of Organic Particles to Predict Macroaggregation in the Northern Adriatic Sea. Water 2023, 15, 1665. https://doi.org/10.3390/w15091665
Kraus R, Ivošević DeNardis N. Tracking the Spatio-Temporal Distribution of Organic Particles to Predict Macroaggregation in the Northern Adriatic Sea. Water. 2023; 15(9):1665. https://doi.org/10.3390/w15091665
Chicago/Turabian StyleKraus, Romina, and Nadica Ivošević DeNardis. 2023. "Tracking the Spatio-Temporal Distribution of Organic Particles to Predict Macroaggregation in the Northern Adriatic Sea" Water 15, no. 9: 1665. https://doi.org/10.3390/w15091665
APA StyleKraus, R., & Ivošević DeNardis, N. (2023). Tracking the Spatio-Temporal Distribution of Organic Particles to Predict Macroaggregation in the Northern Adriatic Sea. Water, 15(9), 1665. https://doi.org/10.3390/w15091665