

Cyanide Biodegradation by a Native Bacterial Consortium and Its Potential for Goldmine Tailing Biotreatment

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Native Consortium Sampling
2.2. Bacterial Isolation
2.3. DNA Extraction and 16S rRNA Amplification
2.4. Sequencing and Analyses
2.5. Mass Spectrometry Proteomic Fingerprint
2.6. Cyanide Biodegradation
2.7. Kinetic Biodegradation Models
3. Results
3.1. Bacterial Isolation and Identification of Native Cyanotrophic Consortium
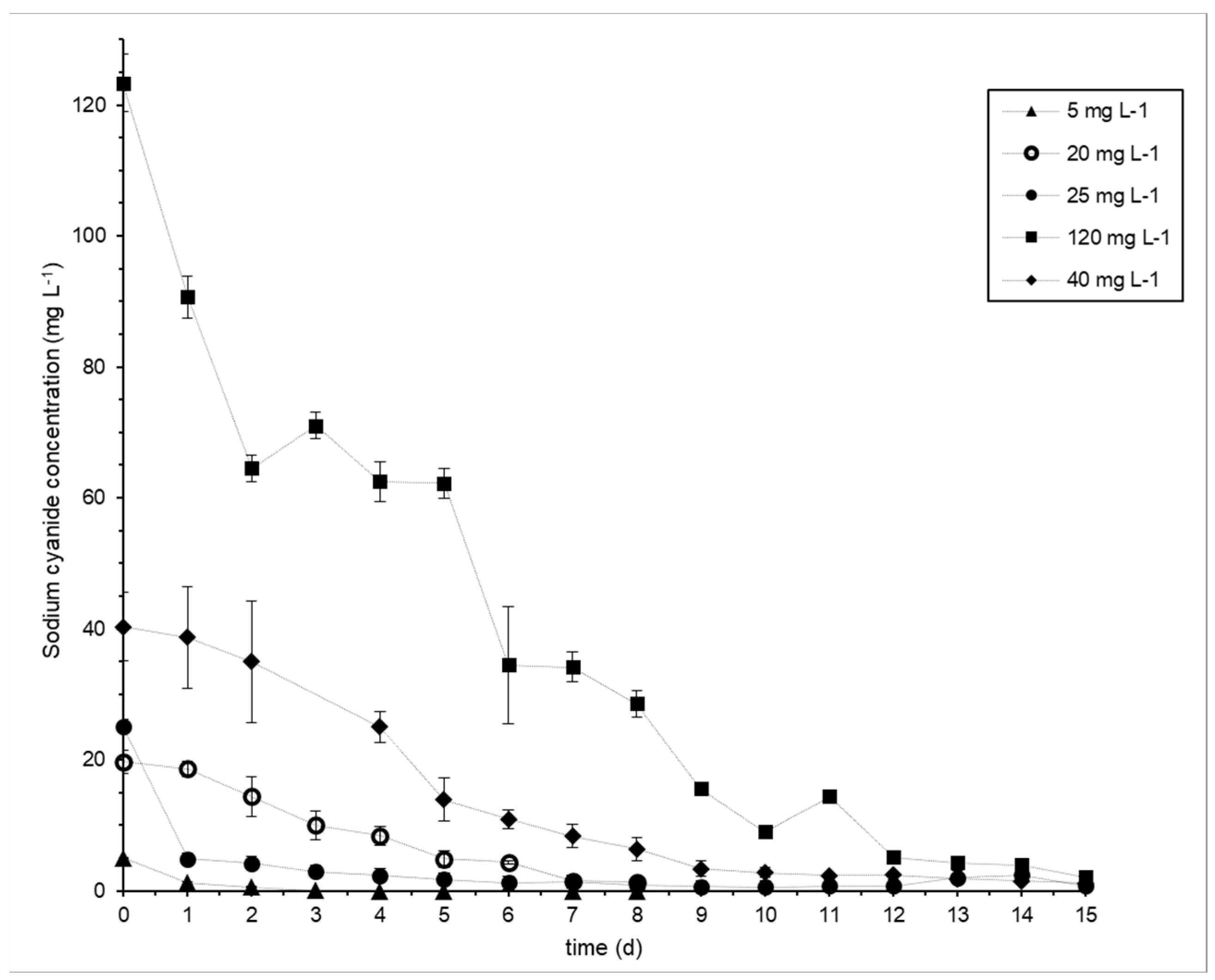
3.2. Cyanide Biodegradation
3.3. Kinetic Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, M.; Akhter, Y.; Chatterjee, S. A review on remediation of cyanide containing industrial wastes using biological systems with special reference to enzymatic degradation. World J. Microbiol. Biotechnol. 2019, 35, 70. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Sewell, B.T.; Benedik, M.J. Cyanide bioremediation: The potential of engineered nitrilases. Appl. Microbiol. Biotechnol. 2017, 101, 3029–3042. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, S.; Dananjeyan, B.; Krishnamurthy, K.; Benckiser, G. Aerobic cyanide degradation by bacterial isolates from cassava factory wastewater. Braz. J. Microbiol. 2015, 46, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, M.A.; Paksirajan, K.; Lens, P.N.L. Assessment of the effluent quality from a gold mining industry in Ghana. Environ. Sci. Pollut. Res. 2012, 20, 3799–3811. [Google Scholar] [CrossRef] [PubMed]
- Alvillo-Rivera, A.; Garrido-Hoyos, S.; Buitrón, G.; Thangarasu-Sarasvathi, P.; Rosano-Ortega, G. Biological treatment for the degradation of cyanide: A review. J. Mater. Res. Technol. 2021, 12, 1418–1433. [Google Scholar] [CrossRef]
- Cyanide in Drinking-Water. Background Document for Development of WHO Guidelines for Drinking-Water Quality. World Health Organization. Available online: https://cdn.who.int/media/docs/default-source/wash-documents/wash-chemicals/cyanide-background.pdf?sfvrsn=29c5b9f4_4 (accessed on 29 January 2022).
- US-EPA. Cyanide Clarification of Free and Total Cyanide Analysis for Safe Drinking Water Act (SDWA) Compliance, Revision 1.0. Available online: https://www.epa.gov/sites/production/files/2016-08/documents/cyanide-clarification-free-and-total-cyanide-analysis-safe-drinking-water.pdf (accessed on 30 January 2023).
- Ganji, S.M.S.A.; Hayati, M. Selecting an appropriate method to remove cyanide from the wastewater of Moteh gold mine using a mathematical approach. Environ. Sci. Pollut. Res. 2018, 25, 23357–23369. [Google Scholar] [CrossRef]
- Kumar, R.; Saha, S.; Dhaka, S.; Kurade, M.B.; Kang, C.U.; Baek, S.H.; Jeon, B.-H. Remediation of cyanide-contaminated environments through microbes and plants: A review of current knowledge and future perspectives. Geosystem Eng. 2016, 20, 28–40. [Google Scholar] [CrossRef]
- Cabello, P.; Luque-Almagro, V.M.; Olaya-Abril, A.; Sáez, L.P.; Moreno-Vivián, C.; Roldán, M.D. Assimilation of cyanide and cyano-derivatives by Pseudomonas pseudoalcaligenes CECT5344: From omic approaches to biotechnological applications. FEMS Microbiol. Lett. 2018, 365, fny032. [Google Scholar] [CrossRef]
- Mekuto, L.; Ntwampe, S.; Akcil, A. An integrated biological approach for treatment of cyanidation wastewater. Sci. Total Environ. 2016, 571, 711–720. [Google Scholar] [CrossRef]
- Mahendran, R.; Bs, S.; Thandeeswaran, M.; Kg, K.; Vijayasarathy, M.; Angayarkanni, J.; Muthusamy, G. Microbial (Enzymatic) Degradation of Cyanide to Produce Pterins as Cofactors. Curr. Microbiol. 2019, 77, 578–587. [Google Scholar] [CrossRef]
- Ibrahim, K.K.; Syed, M.A.; Shukor, M.Y.; Ahmad, S.A. Biological Remediation of Cyanide: A Review. Biotropia 2015, 22, 151–163. [Google Scholar] [CrossRef]
- Lovasoa, C.R.; Hela, K.; Harinaivo, A.A.; Hamma, Y. Bioremediation of soil and water polluted by cyanide: A review. Afr. J. Environ. Sci. Technol. 2017, 11, 272–291. [Google Scholar] [CrossRef]
- Luque-Almagro, V.M.; Moreno-Vivián, C.; Roldán, M.D. Biodegradation of cyanide wastes from mining and jewellery industries. Curr. Opin. Biotechnol. 2016, 38, 9–13. [Google Scholar] [CrossRef]
- Gupta, N.; Balomajumder, C.; Agarwal, V.K. Enzymatic mechanism and biochemistry for cyanide degradation: A review. J. Hazard. Mater. 2010, 176, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Luque-Almagro, V.M.; Cabello, P.; Sáez, L.P.; Olaya-Abril, A.; Moreno-Vivián, C.; Roldán, M.D. Exploring anaerobic environments for cyanide and cyano-derivatives microbial degradation. Appl. Microbiol. Biotechnol. 2017, 102, 1067–1074. [Google Scholar] [CrossRef]
- Moradkhani, M.; Yaghmaei, S.; Nejad, Z.G. Biodegradation of Cyanide under Alkaline Conditions by a Strain of Pseudomonas putida Isolated from Gold Mine Soil and Optimization of Process Variables through Response Surface Methodology (RSM). Period. Polytech. Chem. Eng. 2017, 62, 265–273. [Google Scholar] [CrossRef]
- Willey, J.; Sandman, K.; Wood, D.H. Prescott’s Microbiology, 12th ed.; McGraw Hill: New York, NY, USA, 2022. [Google Scholar]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S rRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Madigan, M.T.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D. Brock Biology of Microorganisms, 15th ed.; Pearson: Upper Saddle River, NJ, USA, 2017. [Google Scholar]
- Singh, N.; Balomajumder, C. Batch growth kinetic studies for elimination of phenol and cyanide using mixed microbial culture. J. Water Process Eng. 2016, 11, 130–137. [Google Scholar] [CrossRef]
- Huertas, M.; Sáez, L.; Roldán, M.D.; Luque-Almagro, V.M.; Martínez-Luque, M.; Blasco, R.; Castillo, F.; Moreno-Vivián, C.; Garcia-Garcia, I. Alkaline cyanide degradation by Pseudomonas pseudoalcaligenes CECT5344 in a batch reactor. Influence of pH. J. Hazard. Mater. 2010, 179, 72–78. [Google Scholar] [CrossRef]
- Safa, Z.J.; Aminzadeh, S.; Zamani, M.; Motallebi, M. Significant increase in cyanide degradation by Bacillus sp. M01 PTCC 1908 with response surface methodology optimization. AMB Express 2017, 7, 200. [Google Scholar] [CrossRef]
- Razanamahandry, L.C.; Onwordi, C.T.; Saban, W.; Bashir, A.K.H.; Mekuto, L.; Malenga, E.; Manikandan, E.; Fosso-Kankeu, E.; Maaza, M.; Ntwampe, S.K.O. Performance of various cyanide degrading bacteria on the biodegradation of free cyanide in water. J. Hazard. Mater. 2019, 380, 120900. [Google Scholar] [CrossRef] [PubMed]
- Roldán, M.D.; Olaya-Abril, A.; Sáez, L.P.; Cabello, P.; Luque-Almagro, V.M.; Moreno-Vivián, C. Bioremediation of cyanide-containing wastes. EMBO Rep. 2021, 22, e53720. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Jin, X.; Wang, F.; Liu, Y.; Chen, X. Successful startup of a full-scale acrylonitrile wastewater biological treatment plant (ACN-WWTP) by eliminating the inhibitory effects of toxic compounds on nitrification. Water Sci. Technol. 2013, 69, 553–559. [Google Scholar] [CrossRef]
- Sankaranarayanan, A.; Gowthami, M. Cyanide Degradation by Consortium of Bacterial Species Isolated from Sago Industry Effluenty. J. Environ. Treat. Tech. 2014, 3, 41–46. [Google Scholar]
- Dwivedi, N.; Balomajumder, C.; Mondal, P. Comparative evaluation of cyanide removal by adsorption, biodegradation, and simultaneous adsorption and biodegradation (SAB) process using Bacillus cereus and almond shell. J. Environ. Biol. 2016, 37, 551–556. [Google Scholar] [PubMed]
- He, Y.-C.; Pan, X.-H.; Xu, X.-F.; Wang, L.-Q. Biosynthesis of Benzoylformic Acid from Benzoyl Cyanide with a New Bacterial Isolate of Brevibacterium sp. CCZU12-1. Appl. Biochem. Biotechnol. 2014, 172, 3223–3233. [Google Scholar] [CrossRef]
- Zhao, J.; Tang, X.; Wang, D.; Gan, X.; Hua, R. Isolation and identification of malathion-degrading strain of Brevundimonas diminuta. In Proceedings of the 2011 International Conference on Electrical and Control Engineering, Yichang, China, 16–18 September 2011. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Tyagi, S.P.; Kaushik, R.; Saxena, A.K. Culturable diversity and functional annotation of psychrotrophic bacteria from cold desert of Leh Ladakh (India). World J. Microbiol. Biotechnol. 2014, 31, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Steinbock, B.; Bechtold, R.; Sevigny, J.L.; Thomas, D.; Thomas, W.K.; Ghosh, A. Draft Genome Sequences of 10 Bacterial Strains Isolated from an Abandoned Coal Mine in Southeast Kansas. Genome Announc. 2019, 8, e01001-19. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, P.; Sánchez-Castro, I.; Descostes, M.; Merroun, M.L. Draft genome sequence data of Microbacterium sp. strain Be9 isolated from uranium-mill tailings porewaters. Data Brief 2020, 31, 105732. [Google Scholar] [CrossRef]
- Román-Ponce, B.; Ramos-Garza, J.; Arroyo-Herrera, I.; Maldonado-Hernández, J.; Bahena-Osorio, Y.; Vásquez-Murrieta, M.S.; Wang, E.T. Mechanism of arsenic resistance in endophytic bacteria isolated from endemic plant of mine tailings and their arsenophore production. Arch. Microbiol. 2018, 200, 883–895. [Google Scholar] [CrossRef]
- Irawati, W.; Patricia; Soraya, Y.; Baskoro, A.H. A Study on Mercury-Resistant Bacteria Isolated from a Gold Mine in Pongkor Village, Bogor, Indonesia. HAYATI J. Biosci. 2012, 19, 197–200. [Google Scholar] [CrossRef]
- Román-Ponce, B.; Li, Y.H.; Murrieta, M.S.V.; Sui, X.H.; Chen, W.F.; Santos, P.E.D.L.; Wang, E.T. Brevibacterium metallicus sp. nov., an endophytic bacterium isolated from roots of Prosopis laegivata grown at the edge of a mine tailing in Mexico. Arch. Microbiol. 2015, 197, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Khamar, Z.; Makhdoumi-Kakhki, A.; Gharaie, M.M. Remediation of cyanide from the gold mine tailing pond by a novel bacterial co-culture. Int. Biodeterior. Biodegradation 2015, 99, 123–128. [Google Scholar] [CrossRef]
- Brüger, A.; Fafilek, G.; Restrepo, B.O.J.; Rojas-Mendoza, L. On the volatilisation and decomposition of cyanide contaminations from gold mining. Sci. Total Environ. 2018, 627, 1167–1173. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef]
- Ojha, S.; Mishra, S.; Kapoor, S.; Chand, S. Synthesis of hexyl α-glucoside and α-polyglucosides by a novel Microbacterium isolate. Appl. Microbiol. Biotechnol. 2013, 97, 5293–5301. [Google Scholar] [CrossRef]
- Buczolits, S.; Schumann, P.; Valens, M.; Rosselló-Mora, R.; Busse, H.-J. Identification of a bacterial strain isolated from the liver of a laboratory mouse as Microbacterium paraoxydans and emended description of the species Microbacterium paraoxydans Laffineur et al 2003. Indian J. Microbiol. 2008, 48, 243–251. [Google Scholar] [CrossRef]
- Bacillus Cereus and Other Bacillus Species, in Bad Bug Book, Foodborne Pathogenic Microorganisms and Natural Toxins. Food and Drug Administration. Available online: https://www.fda.gov/food/foodborne-pathogens/bad-bug-book-second-edition (accessed on 9 August 2022).
- Karthik, C.; Ramkumar, V.S.; Pugazhendhi, A.; Gopalakrishnan, K.; Arulselvi, P.I. Biosorption and biotransformation of Cr(VI) by novel Cellulosimicrobium funkei strain AR6. J. Taiwan Inst. Chem. Eng. 2017, 70, 282–290. [Google Scholar] [CrossRef]
- Bhati, T.; Gupta, R.; Yadav, N.; Singh, R.; Fuloria, A.; Waziri, A.; Chatterjee, S.; Purty, R. Assessment of Bioremediation Potential of Cellulosimicrobium sp. for Treatment of Multiple Heavy Metals. Microbiol. Biotechnol. Lett. 2019, 47, 269–277. [Google Scholar] [CrossRef]
- Flora, S.; Kaushik, P.; Mathur, M.; Bhatnagar, P.; Rawat, N.; Raghuvanshi, P.; Swarnkar, H. Arsenic hyper-tolerance in four Microbacterium species isolated from soil contaminated with textile effluent. Toxicol. Int. 2012, 19, 188–194. [Google Scholar] [CrossRef]
- Sathyavathi, S.; Manjula, A.; Rajendhran, J.; Gunasekaran, P. Extracellular synthesis and characterization of nickel oxide nanoparticles from Microbacterium sp. MRS-1 towards bioremediation of nickel electroplating industrial effluent. Bioresour. Technol. 2014, 165, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, H.; Yang, K.; Ji, B.; Chen, D.; Zhang, H.; Sun, Y.; Tian, J. Autotrophic denitrification by nitrate-dependent Fe(II) oxidation in a continuous up-flow biofilter. Bioprocess Biosyst. Eng. 2015, 39, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, S.; Irudayarajan, L.; Khambhaty, Y.; Palanivel, S. Batch experiments towards remediation of phenolic syntan using individual as well as co-culture of Bacillus cereus and Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2019, 35, 137. [Google Scholar] [CrossRef]
- Shah, S.; Damare, S. Cellular response of Brevibacterium casei #NIOSBA88 to arsenic and chromium—A proteomic approach. Braz. J. Microbiol. 2020, 51, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Rathi, M.; Yogalakshmi, K.N. Brevundimonas diminuta MYS6 associated Helianthus annuus L. for enhanced copper phytoremediation. Chemosphere 2020, 263, 128195. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Huang, P.; Dong, C.; Deng, X. Nickel bioaccumulation by a marine bacterium Brevibacterium sp. (X6) isolated from Shenzhen Bay, China. Mar. Pollut. Bull. 2021, 170, 112656. [Google Scholar] [CrossRef] [PubMed]
- Saggu, S.K.; Jha, G.; Mishra, P.C. Enzymatic Degradation of Biofilm by Metalloprotease From Microbacterium sp. SKS10. Front. Bioeng. Biotechnol. 2019, 7, 192. [Google Scholar] [CrossRef]
- Ojha, S.; Kapoor, S.; Mishra, S. Carotenoid Production by a Novel Isolate of Microbacterium paraoxydans. Indian J. Microbiol. 2017, 58, 118–122. [Google Scholar] [CrossRef]
- Silva, J.A.; Tobella, L.M.; Becerra, J.; Godoy, F.; Martínez, M.A. Biosynthesis of poly-β-hydroxyalkanoate by Brevundimonas vesicularis LMG P-23615 and Sphingopyxis macrogoltabida LMG 17324 using acid-hydrolyzed sawdust as carbon source. J. Biosci. Bioeng. 2007, 103, 542–546. [Google Scholar] [CrossRef]
- Ferracini-Santos, L.; Sato, H.H. Production of alkaline protease from Cellulosimicrobium cellulans. Braz. J. Microbiol. 2009, 40, 54–60. [Google Scholar] [CrossRef]


{kind=link}
| Colonies | Microbiological Characteristics | 16S rRNA Identification | MALDI-TOF MS 4 Fingerprint | |||||
|---|---|---|---|---|---|---|---|---|
| Appearance in Agar R2A 1 | Gram Staining | Spore Staining | Species/Genus | IP 2 (%) | Sim 3 (%) | Species | Max. Factor 5 | |
| C1, C2, C3, C4, C5, C9, C11, C13, C14, C15 | Transparent yellowish, creamy | Positive | Positive | Bacillus cereus | 100 | 100 | Bacillus cereus | 2.441 |
| C6, C7, C8, C18, C20, C21, C23 | Transparent yellow, smooth | Positive | Positive | Microbacterium paraoxydans | 100 | 99.61 | Microbacterium paraoxydans | 2.248 |
| C10, C12, C17, C19 | Transparent pink to brownish, smooth | Negative | Negative | Brevundimonas vesicularis | 99.87 | 100 | Brevundimonas vesicularis | 2.063 |
| C16 | Yellowish, smooth | Positive | Negative | Cellulosimicrobium sp. | 99.61 | 99.61 | Cellulosimicrobium cellulans | 1.693 |
| C22 | Whitish and sticky | Positive | Negative | Brevibacterium casei | 100 | 100 | Brevibacterium casei | 1.714 |
| F-CN0 1 (mg L−1) | PF-CN 2 (mg L−1 OD540nm−1 d−1) | BE 3 (%) | Kinetic Model | ||||||
|---|---|---|---|---|---|---|---|---|---|
| OD540nm 4 | S05 (mg L−1) | First-Order Kinetic Model | Second-Order Kinetic Model | ||||||
| k1 6 (d−1) | R2 7 | k2 8 (mg.mg−1 d−1) | kS 9 (mg L−1) | R2 7 | |||||
| 5 | 0.05 | 99.8 | 0.301 | 5.92 | 0.1151 | 0.9773 | 0.2314 | 307.796 | 0.9058 |
| 20 | * | 93.2 | 0.241 | 19.68 | 0.133 | 0.9795 | 0.0319 | 31.879 | 0.8987 |
| 25 | 0.19 | 97 | 0.355 | 25.13 | 0.109 | 0.9898 | 0.1378 | 19.002 | 0.8913 |
| 40 | * | 97.15 | 0.379 | 40.35 | 0.1126 | 0.9924 | 0.0545 | 54.792 | 0.8415 |
| 120 | * | 98.3 | 0.414 | 123.37 | 0.1344 | 0.988 | 0.0283 | 137.129 | 0.7826 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado-López, M.J.; Garrido-Hoyos, S.E.; Raynal-Gutiérrez, M.E.; El-Kassis, E.G.; Luque-Almagro, V.M.; Rosano-Ortega, G. Cyanide Biodegradation by a Native Bacterial Consortium and Its Potential for Goldmine Tailing Biotreatment. Water 2023, 15, 1595. https://doi.org/10.3390/w15081595
Alvarado-López MJ, Garrido-Hoyos SE, Raynal-Gutiérrez ME, El-Kassis EG, Luque-Almagro VM, Rosano-Ortega G. Cyanide Biodegradation by a Native Bacterial Consortium and Its Potential for Goldmine Tailing Biotreatment. Water. 2023; 15(8):1595. https://doi.org/10.3390/w15081595
Chicago/Turabian StyleAlvarado-López, María José, Sofía E. Garrido-Hoyos, María Elena Raynal-Gutiérrez, Elie G. El-Kassis, Víctor M. Luque-Almagro, and Genoveva Rosano-Ortega. 2023. "Cyanide Biodegradation by a Native Bacterial Consortium and Its Potential for Goldmine Tailing Biotreatment" Water 15, no. 8: 1595. https://doi.org/10.3390/w15081595
APA StyleAlvarado-López, M. J., Garrido-Hoyos, S. E., Raynal-Gutiérrez, M. E., El-Kassis, E. G., Luque-Almagro, V. M., & Rosano-Ortega, G. (2023). Cyanide Biodegradation by a Native Bacterial Consortium and Its Potential for Goldmine Tailing Biotreatment. Water, 15(8), 1595. https://doi.org/10.3390/w15081595