
Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Chemicals and Samples
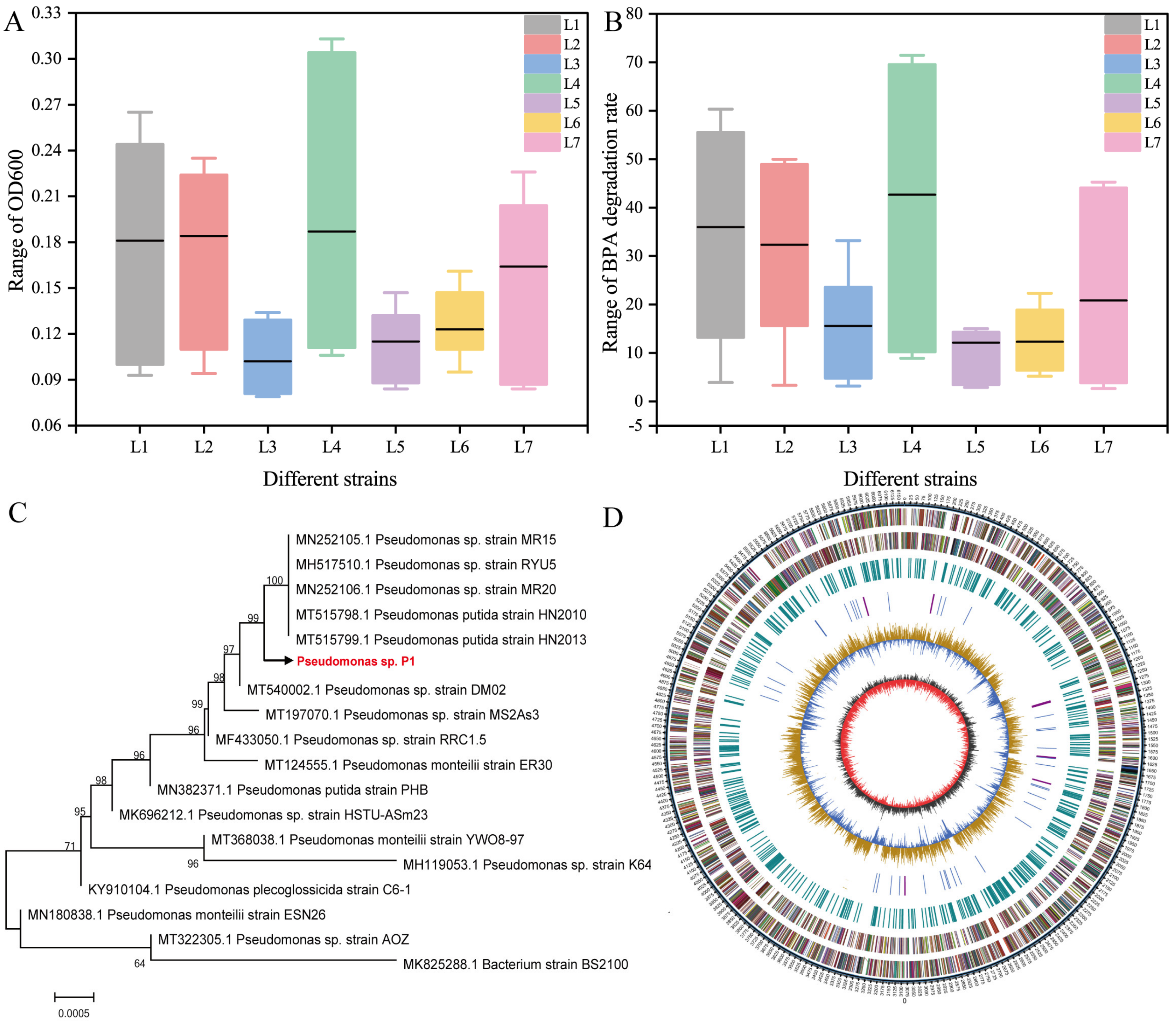
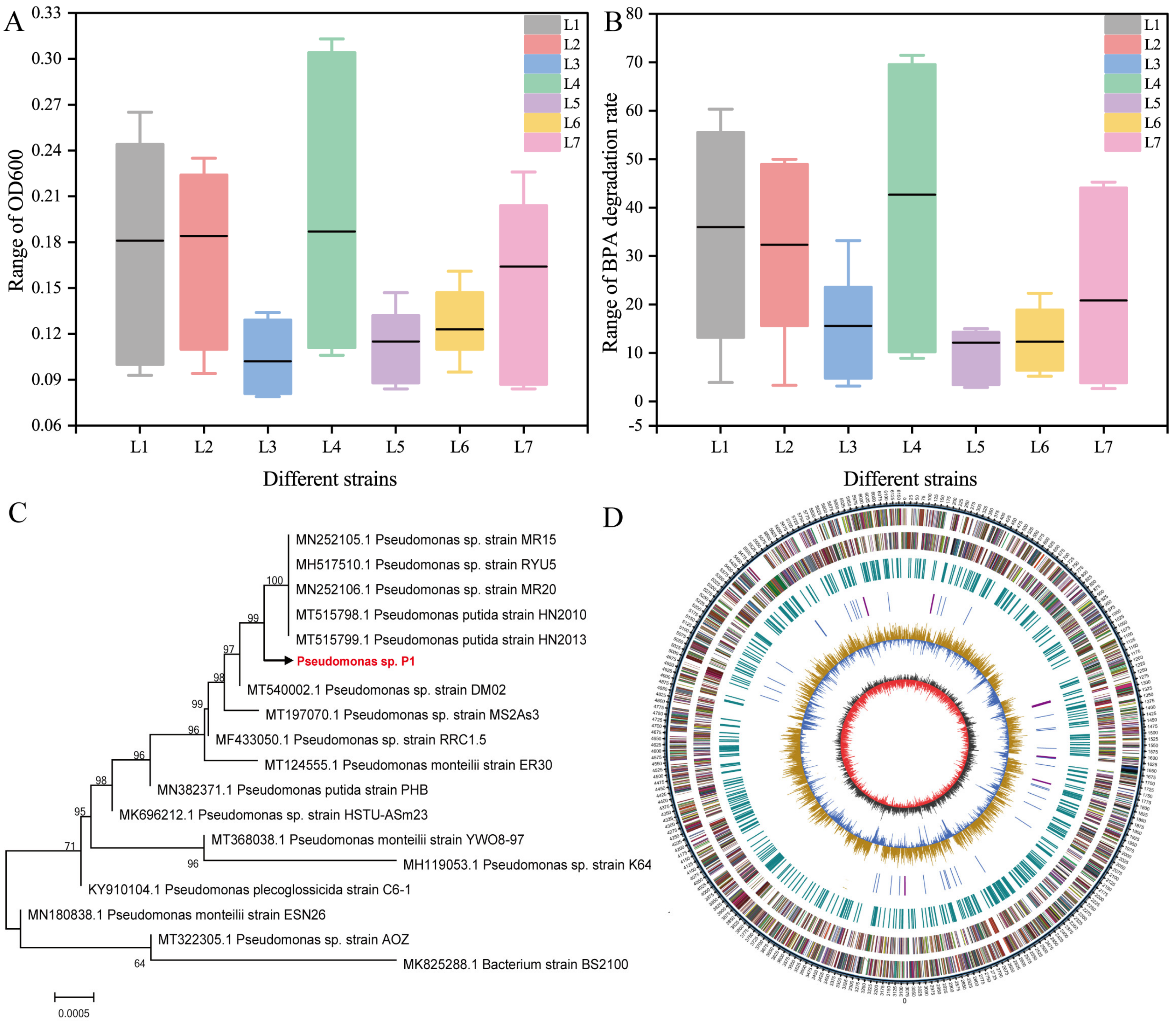
2.2. Isolation and Identification of Strains
2.3. Degradation of BPA by Strain P1
2.4. Genome Sequencing and Functional Annotation
2.5. Amino Acid Sequence Homology Analysis
2.6. PCR, RT-PCR, and qPCR Amplification of Target Genes
3. Results and Discussion
3.1. Screening and Genome Sequencing of BPA-Degrading Bacteria
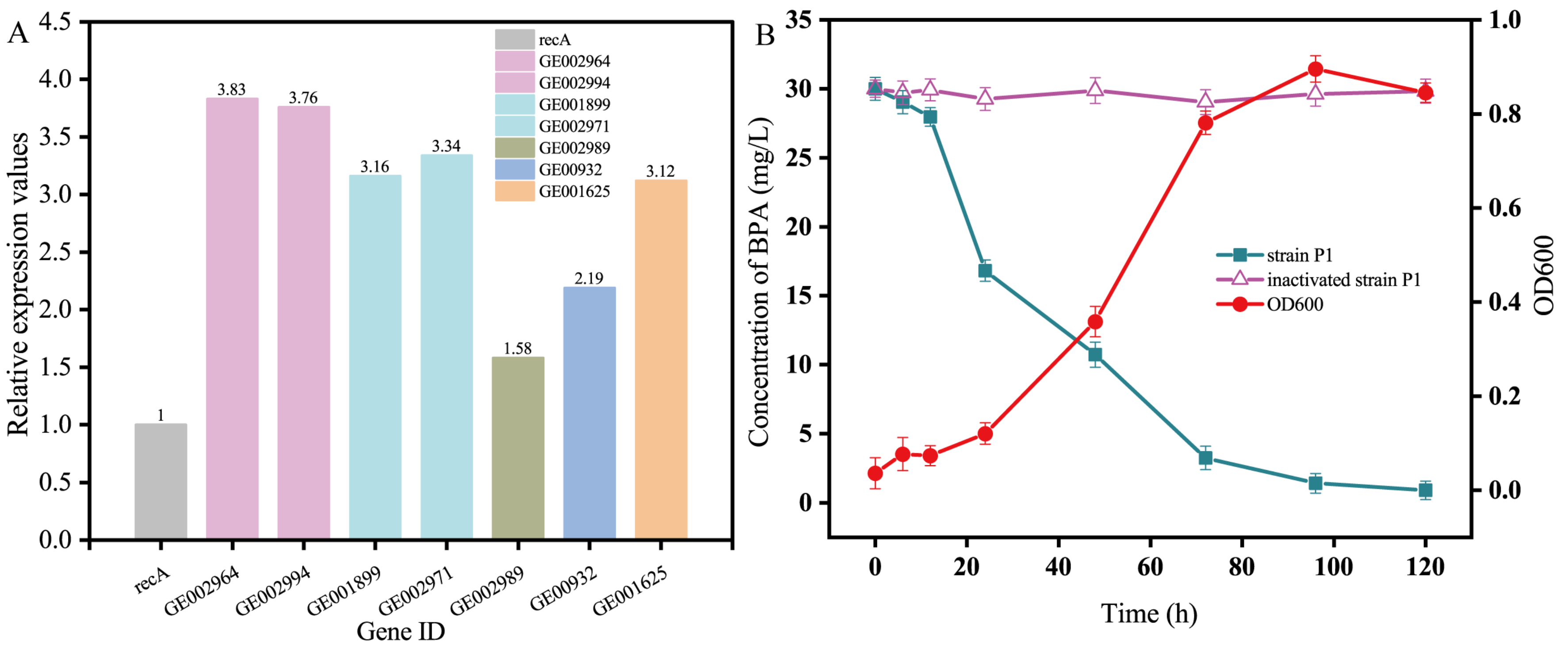
3.2. Screening of BPA Degradation Genes in Pseudomonas sp. P1
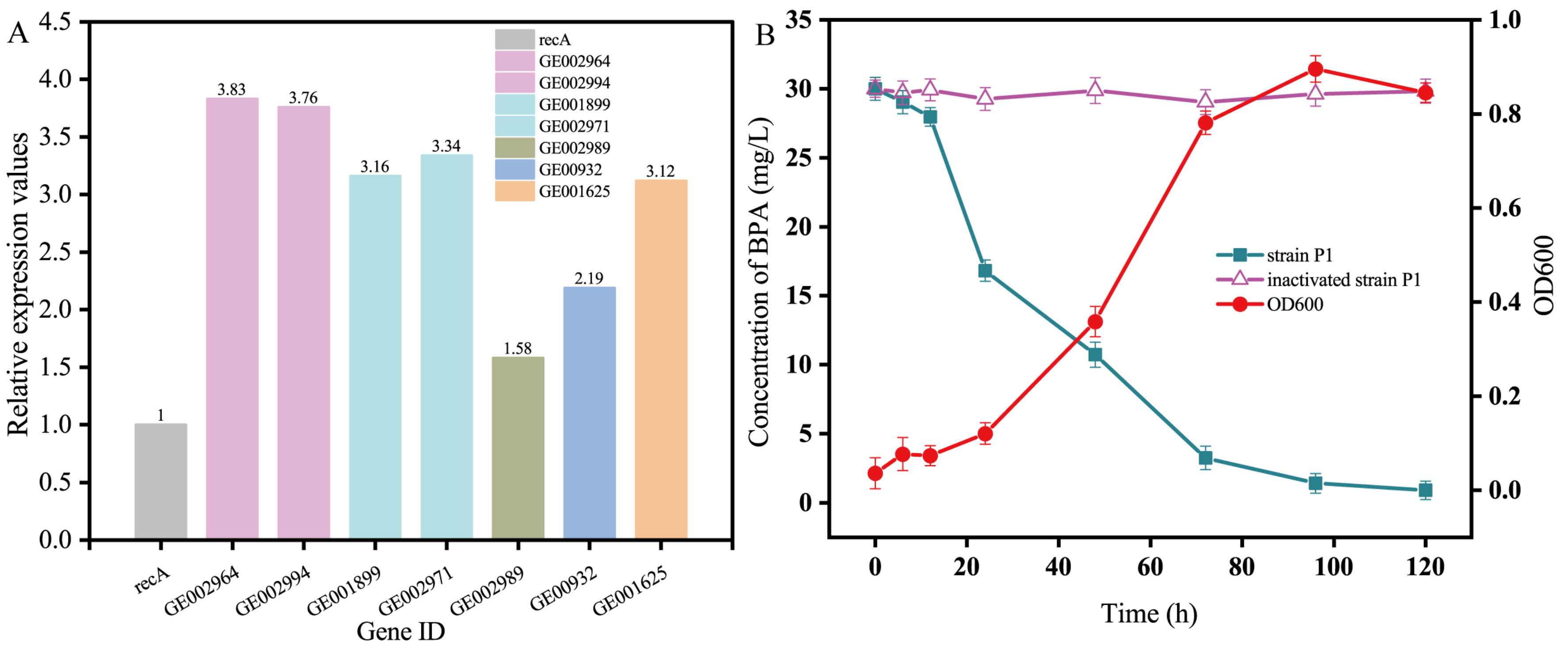
3.3. Expression of the BPA Degradation Gene in Pseudomonas sp. P1
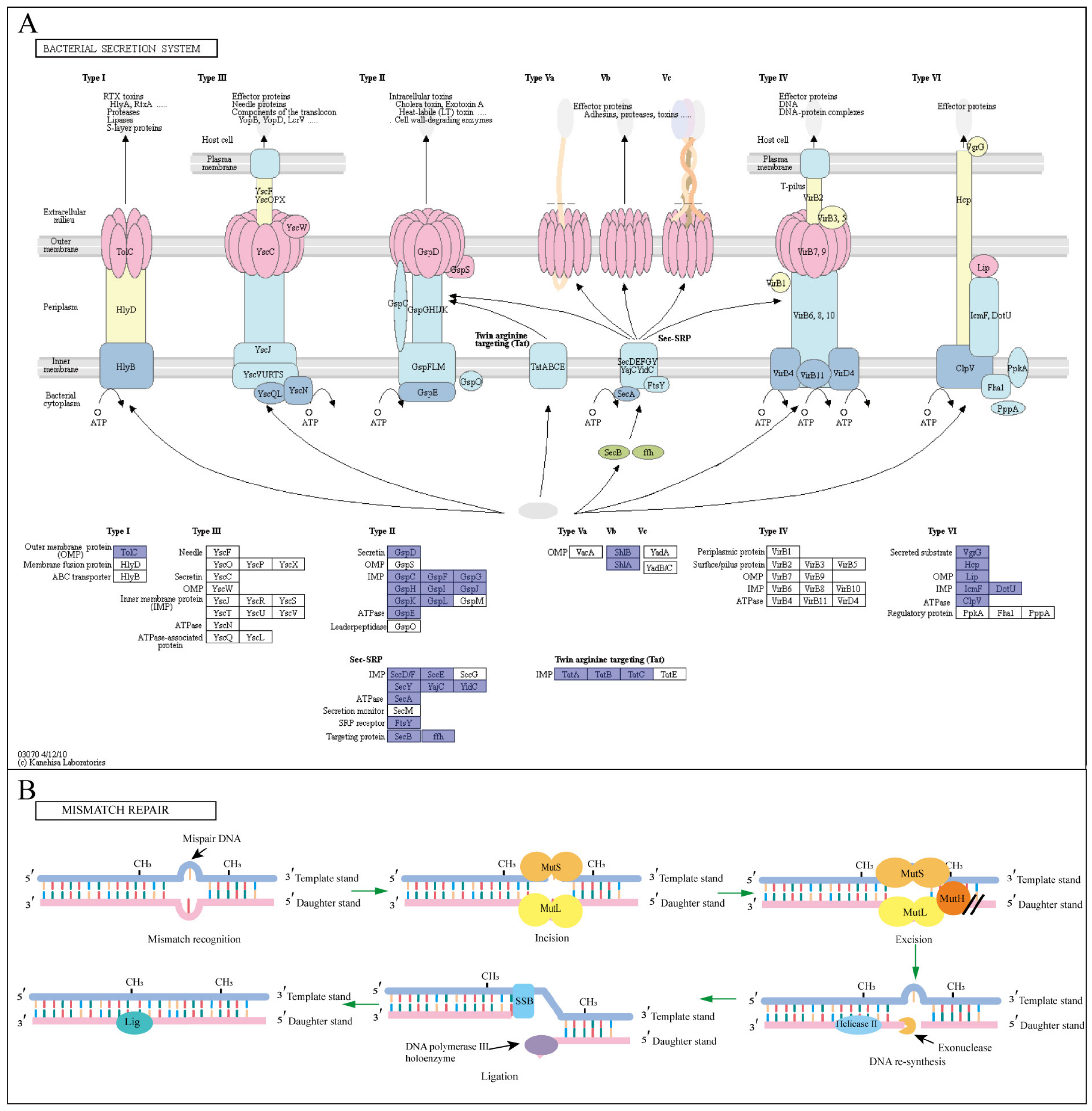
3.4. The Defense Mechanism of Pseudomonas sp. P1 against BPA
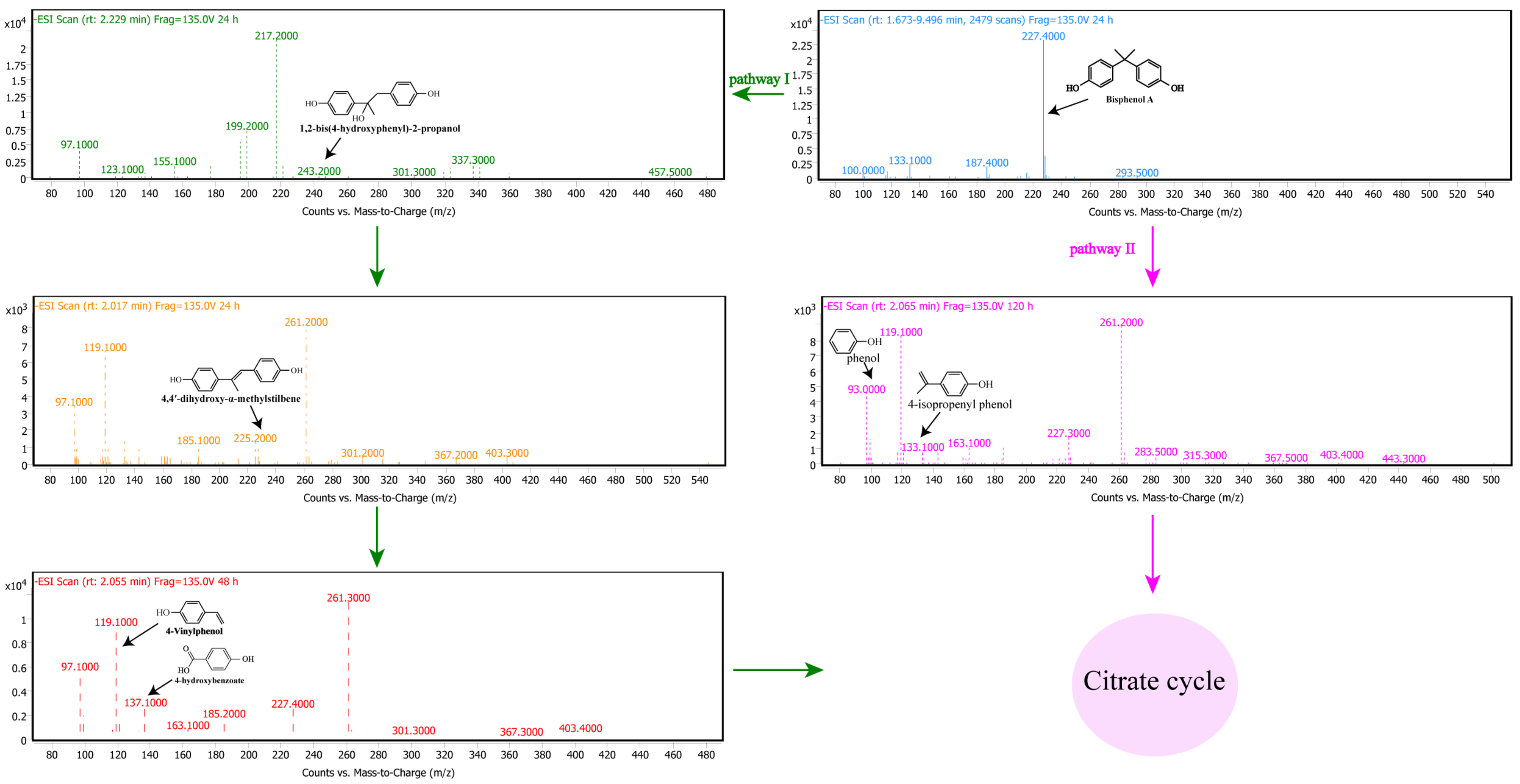
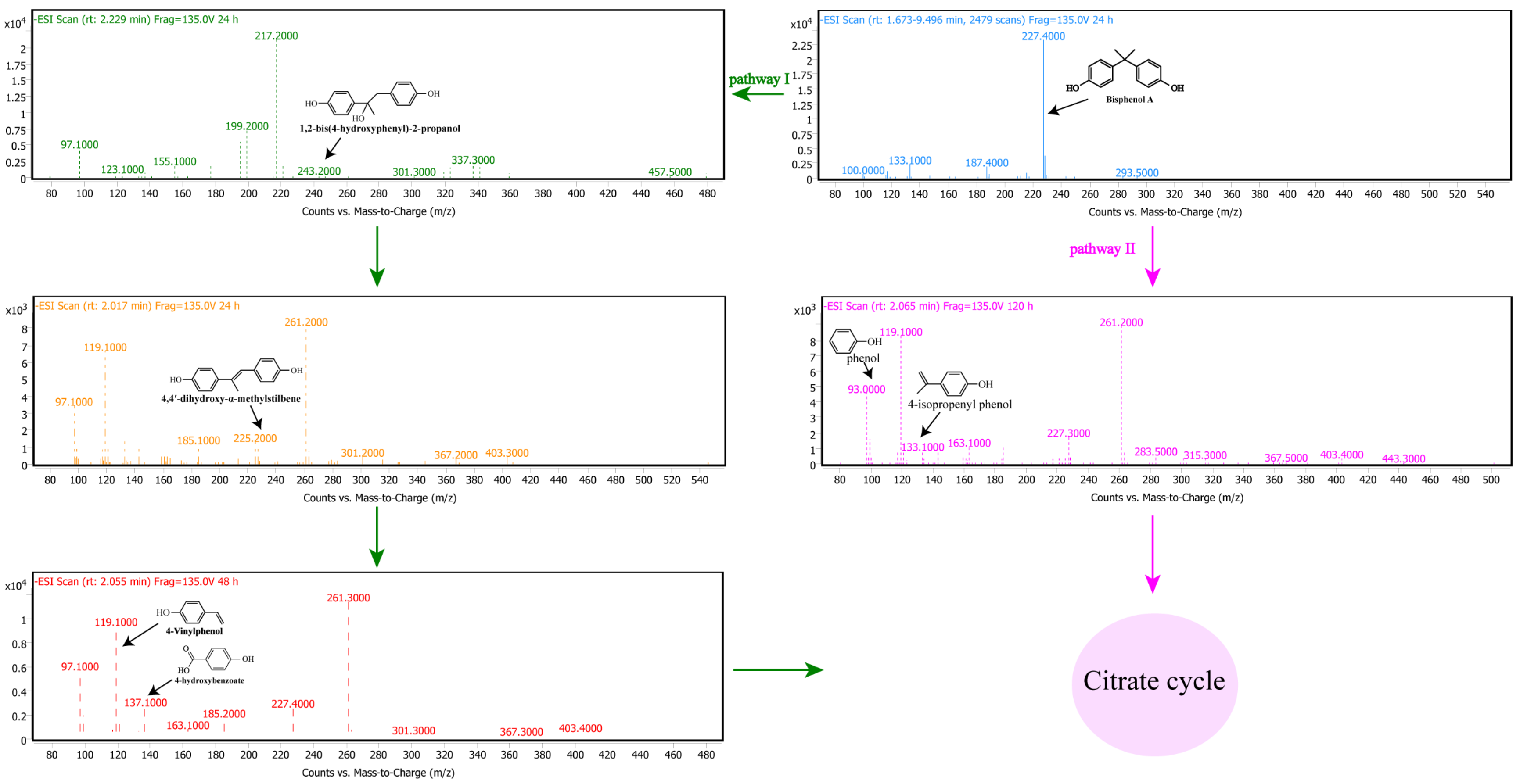
3.5. Pathway of BPA Degradation by Pseudomonas sp. P1
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michalowicz, J. Bisphenol A—Sources, toxicity and biotransformation. Environ. Toxicol. Phar. 2014, 37, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Cantonwine, D.E.; Hauser, R.; Meeker, J.D. Bisphenol A and Human Reproductive Health. Obstet. Gynecol. 2013, 8, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Wazir, U.; Mokbel, K. Bisphenol A: A Concise Review of Literature and a Discussion of Health and Regulatory Implications. In Vivo 2019, 33, 1421–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleme, J.J.; Fan, Y.V.; Tan, R.R.; Jiang, P. Minimising the present and future plastic waste, energy and environmental footprints related to COVID-19. Renew. Sust. Energ. Rev. 2020, 127, 109883. [Google Scholar] [CrossRef]
- Kalina, M.; Tilley, E. “This is our next problem”: Cleaning up from the COVID-19 response. Waste Manag. 2020, 108, 202–205. [Google Scholar] [CrossRef]
- Haldar, S.; Muralidaran, Y.; Miguez, D.; Mulla, S.I.; Mishra, P. Eco-toxicity of nano-plastics and its implication on human metabolism: Current and future perspective. Sci. Total Environ. 2023, 861, 160571. [Google Scholar] [CrossRef]
- Arnold, S.M.; Clark, K.E.; Staples, C.A.; Klecka, G.M.; Dimond, S.S.; Caspers, N.; Hentges, S.G. Relevance of drinking water as a source of human exposure to bisphenol A. J. Expo. Sci. Env. Epidemiol. 2013, 23, 137–144. [Google Scholar] [CrossRef]
- Olatunji, O.S.; Fatoki, O.S.; Opeolu, B.O.; Ximba, B.J.; Chitongo, R. Determination of selected steroid hormones in some surface water around animal farms in Cape Town using HPLC-DAD. Environ. Monit. Assess. 2017, 189, 363. [Google Scholar] [CrossRef]
- Wanda, E.M.M.; Mamba, B.B.; Msagati, T.A.M. Hydrochemical modelling of water quality in terms of emerging micropollutants in Mpumalanga, Gauteng and North West Provinces. Phys. Chem. Earth 2017, 100, 143–157. [Google Scholar] [CrossRef]
- Wanda, E.M.M.; Nyoni, H.; Mamba, B.B.; Msagati, T.A.M. Occurrence of Emerging Micropollutants in Water Systems in Gauteng, Mpumalanga, and North West Provinces, South Africa. Int. J. Env. Pub. Health 2017, 14, 79. [Google Scholar] [CrossRef] [Green Version]
- Maduka Ignatius, C.; Ezeonu Francis, E.; Neboh Emeka, E.; Shu Elvis, N.; Ikekpeazu Ebele, J. BPA and Environmental Estrogen in Potable Water Sources in Enugu Municipality, South-East, Nigeria. Bull. Environ. Contam. Toxicol. 2010, 85, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhao, J.L.; Zhang, C.Y.; Rao, W.L.; Liang, G.H.; Zhang, H.; Liu, Y.H.; Guan, Y.F.; Zhang, H.Y.; Ying, G.G. Profile and removal of bisphenol analogues in hospital wastewater, landfill leachate, and municipal wastewater in South China. Sci. Total Environ. 2021, 790, 148269. [Google Scholar] [CrossRef] [PubMed]
- Andujar, N.; Galvez-Ontiveros, Y.; Zafra-Gomez, A.; Rodrigo, L.; Jesus Alvarez-Cubero, M.; Aguilera, M.; Monteagudo, C.; Rivas, A. Bisphenol A Analogues in Food and Their Hormonal and Obesogenic Effects: A Review. Nutrients 2019, 11, 2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narevski, A.C.; Novakovic, M.I.; Petrovic, M.Z.; Mihajlovic, I.J.; Maodus, N.B.; Vujic, G.V. Occurrence of bisphenol A and microplastics in landfill leachate: Lessons from South East Europe. Environ. Sci. Pollut. Res. 2021, 28, 42196–42203. [Google Scholar] [CrossRef] [PubMed]
- Greca, S.-C.d.A.; Kyrou, I.; Pink, R.; Randeva, H.; Grammatopoulos, D.; Silva, E.; Karteris, E. Involvement of the Endocrine-Disrupting Chemical Bisphenol A (BPA) in Human Placentation. J. Clin. Med. 2020, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Ejaredar, M.; Lee, Y.; Roberts, D.J.; Sauve, R.; Dewey, D. Bisphenol A exposure and children’s behavior: A systematic review. J. Expo. Sci. Environ. Epidemiol. 2017, 27, 175–183. [Google Scholar] [CrossRef]
- Ashfaq, M.; Khan, K.N.; Rehman, M.S.U.; Mustafa, G.; Nazar, M.F.; Sun, Q.; Iqbal, J.; Mulla, S.I.; Yu, C.P. Ecological risk assessment of pharmaceuticals in the receiving environment of pharmaceutical wastewater in Pakistan. Ecotoxicol. Environ. Saf. 2017, 136, 31–39. [Google Scholar] [CrossRef]
- Mokra, K.; Wozniak, K.; Bukowska, B.; Sicinska, P.; Michalowicz, J. Low-concentration exposure to BPA, BPF and BPAF induces oxidative DNA bases lesions in human peripheral blood mononuclear cells. Chemosphere 2018, 201, 119–126. [Google Scholar] [CrossRef]
- Naveira, C.; Rodrigues, N.; Santos, F.S.; Santos, L.N.; Neves, R.A.F. Acute toxicity of Bisphenol A (BPA) to tropical marine and estuarine species from different trophic groups. Environ. Pollut. 2021, 268, 115911. [Google Scholar] [CrossRef]
- Leech, D.M.; Snyder, M.T.; Wetzel, R.G. Natural organic matter and sunlight accelerate the degradation of 17 beta-estradiol in water. Sci. Total Environ. 2009, 407, 2087–2092. [Google Scholar] [CrossRef]
- Sun, Z.; Zhao, L.; Liu, C.; Zhen, Y.; Ma, J. Fast adsorption of BPA with high capacity based on pi-pi electron donoracceptor and hydrophobicity mechanism using an in-situ sp(2) C dominant N-doped carbon. Chem. Eng. J. 2020, 381, 122510. [Google Scholar] [CrossRef]
- Hou, Z.; Wen, Z.; Wang, D.; Wang, J.; Francois-Xavier, C.P.; Wintgens, T. Bipolar jet electrospinning bi-functional nanofibrous membrane for simultaneous and sequential filtration of Cd2+ and BPA from water: Competition and synergistic effect. Chem. Eng. J. 2018, 332, 118–130. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, S.; Liu, X.; Sun, M.; Cao, J.; Wang, J.; Wang, S.; Wen, T.; Wang, X. Oxygen defects-induced charge transfer in Bi7O9I3 for enhancing oxygen activation and visible-light degradation of BPA. Chemosphere 2022, 286, 131783. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.; Franke, M.; Stelter, M.; Braeutigam, P. Degradation of endocrine disruptor bisphenol A by ultrasound-assisted electrochemical oxidation in water. Ultrason. Sonochemistry 2017, 39, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Bharagava, R.N.; Kishor, R.; Purchase, D.; Ferreira, L.; Bilal, M. Environmental and Health Hazards of Textile Industry Wastewater Pollutants and Its Treatment Approaches; Springer: Cham, Switzerland, 2021; pp. 1–24. [Google Scholar] [CrossRef]
- Peng, Y.-H.; Chen, Y.J.; Chang, Y.J.; Shih, Y.H. Biodegradation of bisphenol A with diverse microorganisms from river sediment. J. Hazard. Mater. 2015, 286, 285–290. [Google Scholar] [CrossRef]
- Ben Ouada, S.; Ben Ali, R.; Leboulanger, C.; Ben Ouada, H.; Sayadi, S. Effect of Bisphenol A on the extremophilic microalgal strain Picocystis sp. (Chlorophyta) and its high BPA removal ability. Ecotoxicol. Environ. Saf. 2018, 158, 1–8. [Google Scholar] [CrossRef]
- Li, G.; Xiong, J.; Wong, P.K.; An, T. Enhancing tetrabromobisphenol A biodegradation in river sediment microcosms and understanding the corresponding microbial community. Environ. Pollut. 2016, 208, 796–802. [Google Scholar] [CrossRef]
- Das, R.; Li, G.; Mai, B.; An, T. Spore cells from BPA degrading bacteria Bacillus sp. GZB displaying high laccase activity and stability for BPA degradation. Sci. Total Environ. 2018, 640, 798–806. [Google Scholar] [CrossRef]
- Sakai, K.; Yamanaka, H.; Moriyoshi, K.; Ohmoto, T.; Ohe, T. Biodegradation of bisphenol A and related compounds by Sphingomonas sp. strain BP-7 isolated from seawater. Biosci. Biotech. Biochem. 2007, 71, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Cydzik-Kwiatkowska, A.; Zielinska, M.; Bernat, K.; Bulkowska, K.; Wojnowska-Baryla, I. Insights into mechanisms of bisphenol A biodegradation in aerobic granular sludge. Bioresour. Technol. 2020, 315, 123806. [Google Scholar] [CrossRef]
- Yu, Y.; Yin, H.; Peng, H.; Lu, G.; Dang, Z. Proteomic mechanism of decabromodiphenyl ether (BDE-209) biodegradation by Microbacterium Y2 and its potential in remediation of BDE-209 contaminated water-sediment system. J. Hazard. Mater. 2020, 387, 121708. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, Y.; Mao, Y.; Zhang, T. Partnership of Arthrobacter and Pimelobacter in Aerobic Degradation of Sulfadiazine Revealed by Metagenomics Analysis and Isolation. Environ. Sci. Technol. 2018, 52, 2963–2972. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Yin, C.F.; Sun, L.; Zhang, J.; Xu, Y.; Zhou, N.Y. Biodegradation of bisphenol-A polycarbonate plastic by Pseudoxanthomonas sp. strain NyZ600. J. Hazard. Mater. 2021, 416, 125775. [Google Scholar] [CrossRef]
- Kyrila, G.; Katsoulas, A.; Schoretsaniti, V.; Rigopoulos, A.; Rizou, E.; Doulgeridou, S.; Sarli, V.; Samanidou, V.; Touraki, M. Bisphenol A removal and degradation pathways in microorganisms with probiotic properties. J. Hazard. Mater. 2021, 413, 125363. [Google Scholar] [CrossRef] [PubMed]
- Shobnam, N.; Sun, Y.; Mahmood, M.; Loffler, F.E.; Im, J. Biologically mediated abiotic degradation (BMAD) of bisphenol A by manganese-oxidizing bacteria. J. Hazard. Mater. 2021, 417, 125987. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, K.; Chen, L. Bacteria-mediated bisphenol A degradation. Appl. Microbiol. Biotechnol. 2013, 97, 5681–5689. [Google Scholar] [CrossRef]
- Qiu, Q.; Wang, P.; Kang, H.; Wang, Y.; Tian, K.; Huo, H. Genomic Analysis of a New Estrogen-Degrading Bacterial Strain, Acinetobacter sp. DSSKY-A-001. Int. J. Genomics. 2019, 2019, 2804134. [Google Scholar] [CrossRef] [Green Version]
- Tian, K.; Yu, Y.; Qiu, Q.; Sun, X.; Meng, F.; Bi, Y.; Gu, J.; Wang, Y.; Zhang, F.; Huo, H. Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi. Microorganisms 2023, 11, 67. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Xu, X.; Tian, K.; Zhou, D.; Meng, F.; Zhang, H.; Huo, H. Genomics analysis of the steroid estrogen-degrading bacterium Serratia nematodiphila DH-S01. Biotechnol. Biotechnol. Equip. 2020, 34, 430–440. [Google Scholar] [CrossRef]
- Tian, K.; Meng, F.; Meng, Q.; Gao, Y.; Zhang, L.; Wang, L.; Wang, Y.; Li, X.; Huo, H. The Analysis of Estrogen-Degrading and Functional Metabolism Genes inRhodococcus equi DSSKP-R-001. Int. J. Genom. 2020, 2020, 9369182. [Google Scholar] [CrossRef]
- De Morais Farias, J.; Krepsky, N. Data for: Bacterial degradation of bisphenol analogues: An overview. Environ. Sci. Pollut. 2022, 29, 76543–76564. [Google Scholar] [CrossRef] [PubMed]
- Eltoukhy, A.; Jia, Y.; Nahurira, R.; Abo-Kadoum, M.A.; Khokhar, I.; Wang, J.; Yan, Y. Biodegradation of endocrine disruptor Bisphenol A by Pseudomonas putida strain YC-AE1 isolated from polluted soil, Guangdong, China. BMC Microbiol. 2020, 20, 11. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Liang, J.; Liu, M.; Rui, J.; Shi, J.; Yu, Y.; Zhang, X. Aerobic degradation of bisphenol A by Pseudomonas sp. LM-1: Characteristic and pathway. Biodegradation 2023, 34, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Thathola, P.; Agnihotri, V.; Pandey, A.; Upadhyay, S.K. Biodegradation of bisphenol A using psychrotolerant bacterial strain Pseudomonas palleroniana GBPI_508. Arch. Microbiol. 2022, 204, 272. [Google Scholar] [CrossRef]
- Vijayalakshmi, V.; Senthilkumar, P.; Mophin-Kani, K.; Sivamani, S.; Sivarajasekar, N.; Vasantharaj, S. Bio-degradation of Bisphenol A by Pseudomonas aeruginosa PAb1 isolated from effluent of thermal paper industry: Kinetic modeling and process optimization. J. Radiat. Res. Appl. Sci. 2018, 11, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Akahira, A.; Oshiman, K.I.; Tsuchido, T.; Matsumura, Y. Purification of cytochrome P450 and ferredoxin, involved in bisphenol A degradation, from Sphingomonas sp. strain AO1. Appl. Environ. Microb. 2005, 71, 8024–8030. [Google Scholar] [CrossRef] [Green Version]
- Tong, T.; Li, R.; Chai, M.; Wang, Q.; Yang, Y.; Xie, S. Metagenomic analysis of microbial communities continuously exposed to Bisphenol A in mangrove rhizosphere and non-rhizosphere soils. Sci. Total Environ. 2021, 792, 148486. [Google Scholar] [CrossRef]
- Mandal, K.; Singh, B.; Jariyal, M.; Gupta, V.K. Microbial degradation of fipronil by Bacillus thuringiensis. Ecotoxicol. Environ. Saf. 2013, 93, 87–92. [Google Scholar] [CrossRef]
- Surhio, M.A.; Talpur, F.N.; Nizamani, S.M.; Amin, F.; Bong, C.W.; Lee, C.W.; Ashraf, M.A.; Shah, M.R. Complete degradation of dimethyl phthalate by biochemical cooperation of the Bacillus thuringiensis strain isolated from cotton field soil. Rsc. Adv. 2014, 4, 55960–55966. [Google Scholar] [CrossRef]
- Bathelt, C.M.; Ridder, L.; Mulholland, A.J.; Harvey, J.N. Aromatic hydroxylation by cytochrome P450: Model calculations of mechanism and substituent effects. J. Am. Chem. Soc. 2003, 125, 15004–15005. [Google Scholar] [CrossRef] [PubMed]
- Rittle, J.; Green, M.T. Cytochrome P450 Compound I: Capture, Characterization, and C-H Bond Activation Kinetics. Science 2010, 330, 933–937. [Google Scholar] [CrossRef] [Green Version]
- Degtyarenko, K.N. Structural domains of P450-containing monooxygenase systems. Protein Eng. Des. Sel. 1995, 8, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Zhao, H.; Yin, H.; Peng, H.; Tang, L.; Gao, J.; Ma, Y. Triphenyltin biodegradation and intracellular material release by Brevibacillus brevis. Chemosphere 2014, 105, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Bi, X.; Peng, Y.; Bai, M. Research advances of the phosphorus-accumulating organisms of Candidatus Accumulibacter, Dechloromonas and Tetrasphaera: Metabolic mechanisms, applications and influencing factors. Chemosphere 2022, 307, 135675. [Google Scholar] [CrossRef]
- Hui, X.; Chen, Z.; Zhang, J.; Lu, M.; Cai, X.; Deng, Y.; Hu, Y.; Wang, Y. Computational prediction of secreted proteins in gram-negative bacteria. Comput. Struct. Biotechnol. J. 2021, 19, 1806–1828. [Google Scholar] [CrossRef] [PubMed]
- Kudva, R.; Denks, K.; Kuhn, P.; Vogt, A.; Mueller, M.; Koch, H.G. Protein translocation across the inner membrane of Gram-negative bacteria: The Sec and Tat dependent protein transport pathways. Res. Microbiol. 2013, 164, 505–534. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.; Sargent, F.; Berks, B.C. The Tat Protein Export Pathway. EcoSal Plus 2010, 4. [Google Scholar] [CrossRef]
- Prajapati, B.; Bernal-Cabas, M.; Lopez-Alvarez, M.; Schaffer, M.; Bartel, J.; Rath, H.; Steil, L.; Becher, D.; Volker, U.; Mader, U.; et al. Double trouble: Bacillus depends on a functional Tat machinery to avoid severe oxidative stress and starvation upon entry into a NaCl-depleted environment. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118914. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, Y.; Hou, J.; Zhou, X.; Chen, J.; Yao, C.; Zhang, Y.; Li, Y. Biodegradation of bisphenol A by alginate immobilized Phanerochaete chrysosporium beads: Continuous cyclic treatment and degradation pathway analysis. Biochem. Eng. J. 2022, 177, 108212. [Google Scholar] [CrossRef]
- Watkins, D.J.; Ferguson, K.K.; Del Toro, L.V.A.; Alshawabkeh, A.N.; Cordero, J.F.; Meeker, J.D. Associations between urinary phenol and paraben concentrations and markers of oxidative stress and inflammation among pregnant women in Puerto Rico. Int. J. Hyg. Environ. Health 2015, 218, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Lu, S.; Dai, Y.; Rui, C.; Wang, Y.; Zhou, Y.; Li, Y.; Pang, Q.; Fan, R. Higher dermal exposure of cashiers to BPA and its association with DNA oxidative damage. Environ. Int. 2017, 98, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P. Molecular mechanisms of DNA mismatch repair. DNA Repair. 2001, 486, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yu, H.; Qin, H.; Long, Y.; Ye, J.; Qu, Y. Bisphenol A degradation pathway and associated metabolic networks in Escherichia coli harboring the gene encoding CYP450. J. Hazard. Mater. 2020, 388, 121737. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Yang, P. BPA biodegradation driven by isolated strain SQ-2 and its metabolism mechanism elucidation. Biochem. Eng. J. 2022, 185, 108540. [Google Scholar] [CrossRef]
- Fukuda, T.; Uchida, H.; Takashima, Y.; Uwajima, T.; Kawabata, T.; Suzuki, M. Degradation of bisphenol a by purified laccase from Trametes villosa. Biochem. Biophys. Res. Commun. 2001, 284, 704–706. [Google Scholar] [CrossRef]
- Galliker, P.; Hommes, G.; Schlosser, D.; Corvini, P.F.X.; Shahgaldian, P. Laccase-modified silica nanoparticles efficiently catalyze the transformation of phenolic compounds. J. Colloid Interface Sci. 2010, 349, 98–105. [Google Scholar] [CrossRef]
- Huang, Q.G.; Weber, W.J. Transformation and removal of bisphenol A from aqueous phase via peroxidase-mediated oxidative coupling reactions: Efficacy, products, and pathways. Environ. Sci. Technol. 2005, 39, 6029–6036. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Temperature (°C) | Initial pH | Inoculum Size (%, v/v) | Initial Concentration of BPA (mg/L) |
|---|---|---|---|
| 20, 25, 30, 35, 40 | 5, 6, 7, 8 | 1, 3, 5, 7, 9 | 15, 30, 50, 70, 100 |
| Gene ID | Gene Name | Enzyme | Expression Level | Homologous Protein (Sequence Identity) |
|---|---|---|---|---|
| GE000741 | P-hydroxybenzoate hydroxylase | - | Hydroxylase a (39.39%) | |
| GE003776 | CotA | Copper oxidase | - | CotAb (24.32%) |
| GE000932 | Copper oxidase | 2.19 | CotAb (22.9%) | |
| GE001625 | Lac | Multi-copper polyphenol oxidoreductase laccase | 3.12 | Lacc (26.89) |
| GE001899 | bisdA | Ferredoxin | 3.16 | bisdAd (30%) |
| GE002971 | Ferredoxin | 3.34 | bisdAd (31.37%) | |
| GE002964 | P450 | 3.83 | bisdBe (27.86%) | |
| GE002994 | Cytochrome P450 | 3.76 | bisdBe (24.86%) | |
| GE002989 | ProJ | Hydrolase | 1.58 | proJf (27.05%) |
| GE003785 | Aldo/keto reductase | - | reductase g (32.01%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Tian, K.; Qiu, Q.; Yu, Y.; Li, H.; Chang, M.; Sun, X.; Gu, J.; Zhang, F.; Wang, Y.; et al. Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1. Water 2023, 15, 830. https://doi.org/10.3390/w15040830
Li S, Tian K, Qiu Q, Yu Y, Li H, Chang M, Sun X, Gu J, Zhang F, Wang Y, et al. Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1. Water. 2023; 15(4):830. https://doi.org/10.3390/w15040830
Chicago/Turabian StyleLi, Shuaiguo, Kejian Tian, Qing Qiu, Yue Yu, Han Li, Menghan Chang, Xuejian Sun, Jinming Gu, Fenglin Zhang, Yibing Wang, and et al. 2023. "Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1" Water 15, no. 4: 830. https://doi.org/10.3390/w15040830