
Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Climate Change and Its Impacts on Aquatic Ecosystems
1.2. Phytoplankton as Indicators of Climate Change
2. Materials and Methods
Data Sources, Search Criteria, Analyses
3. Results and Discussion
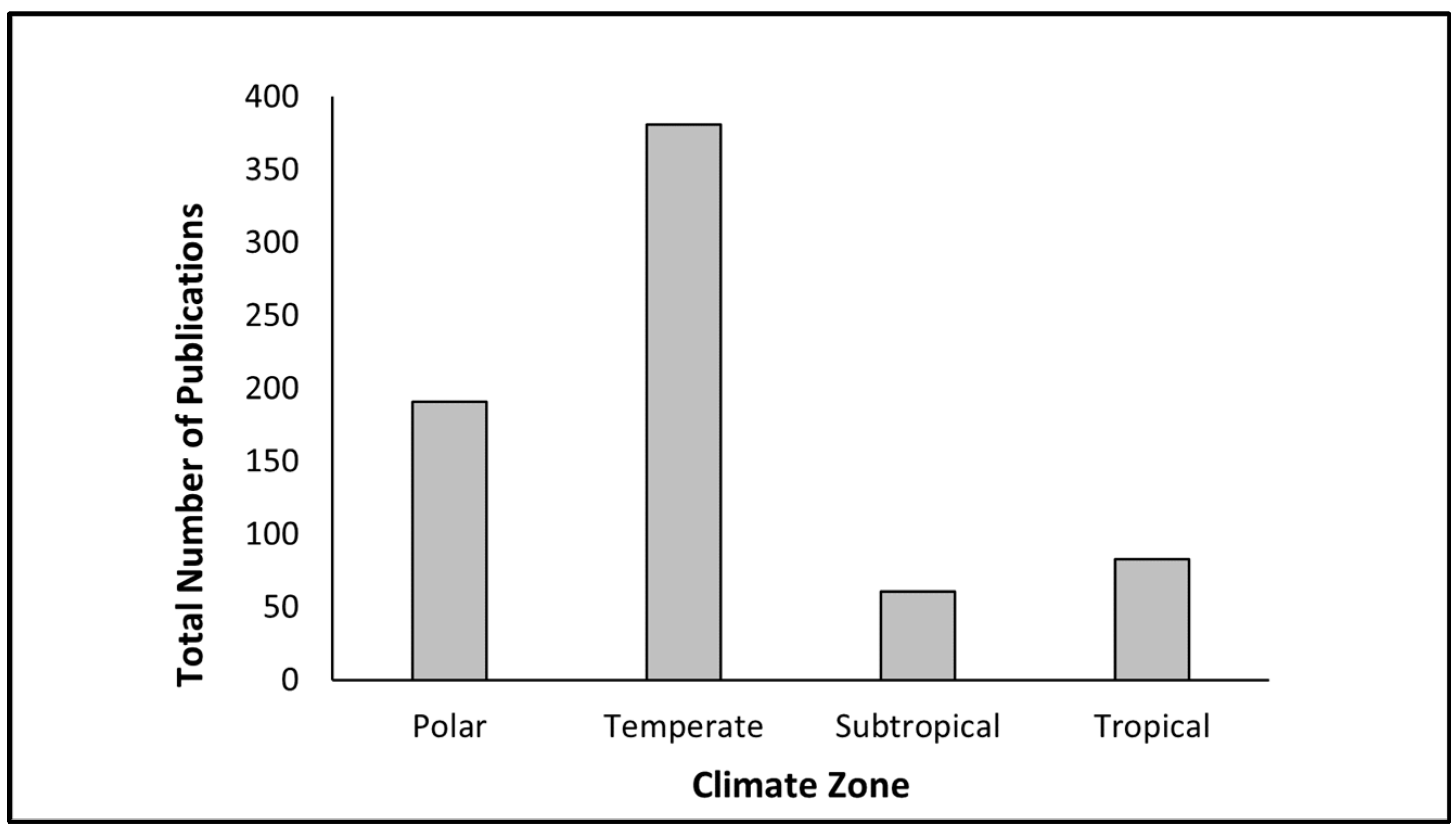
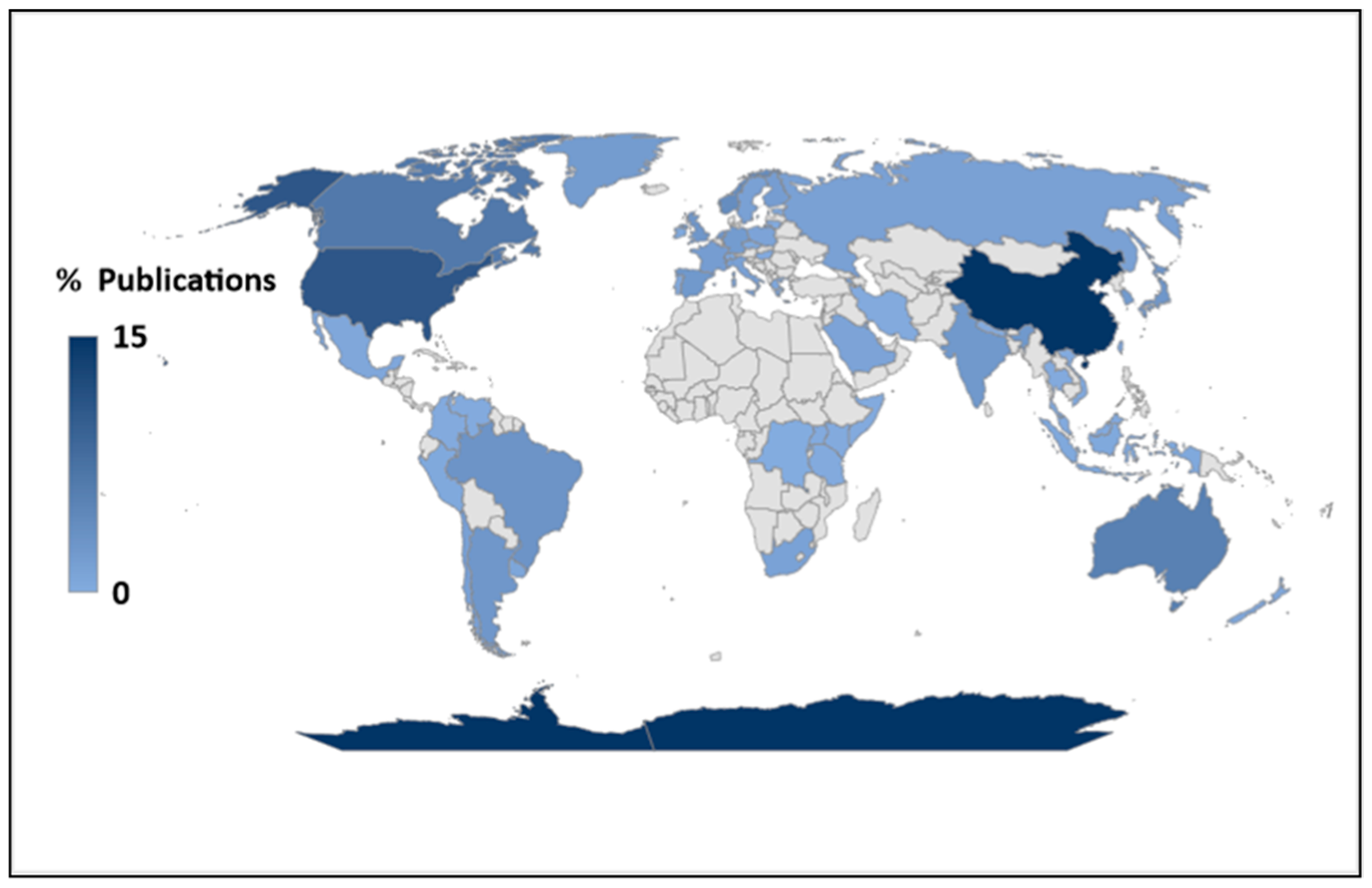
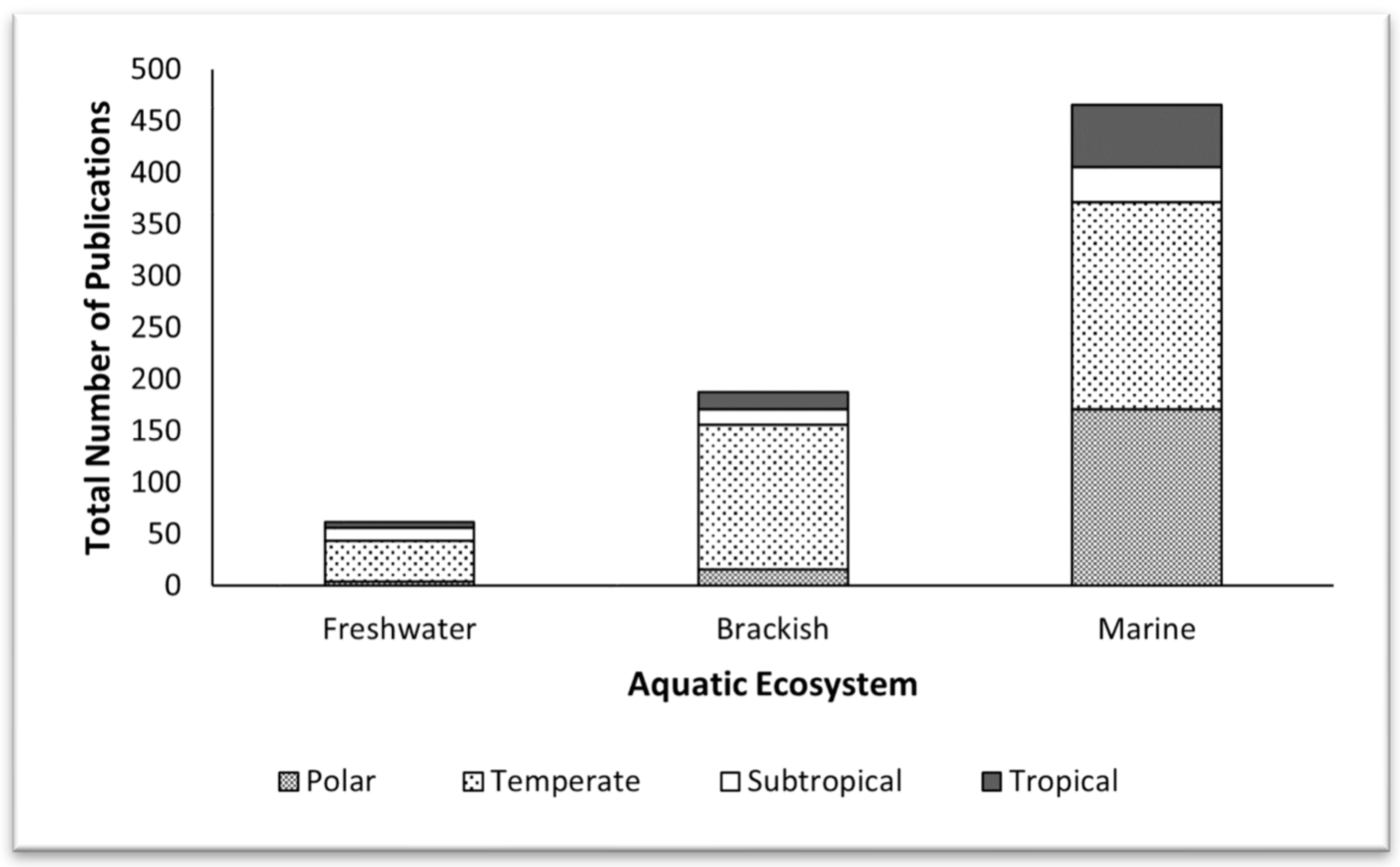
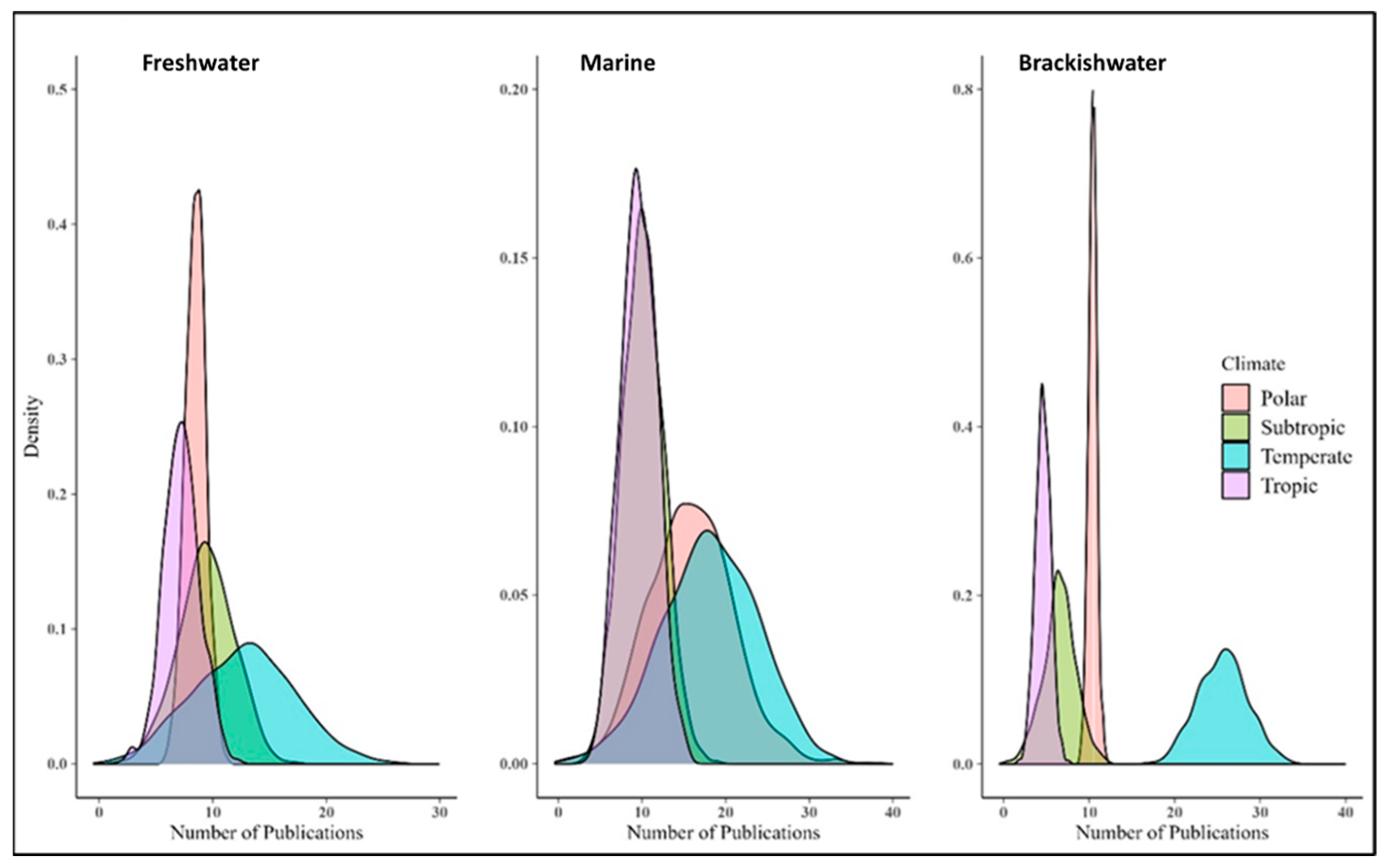
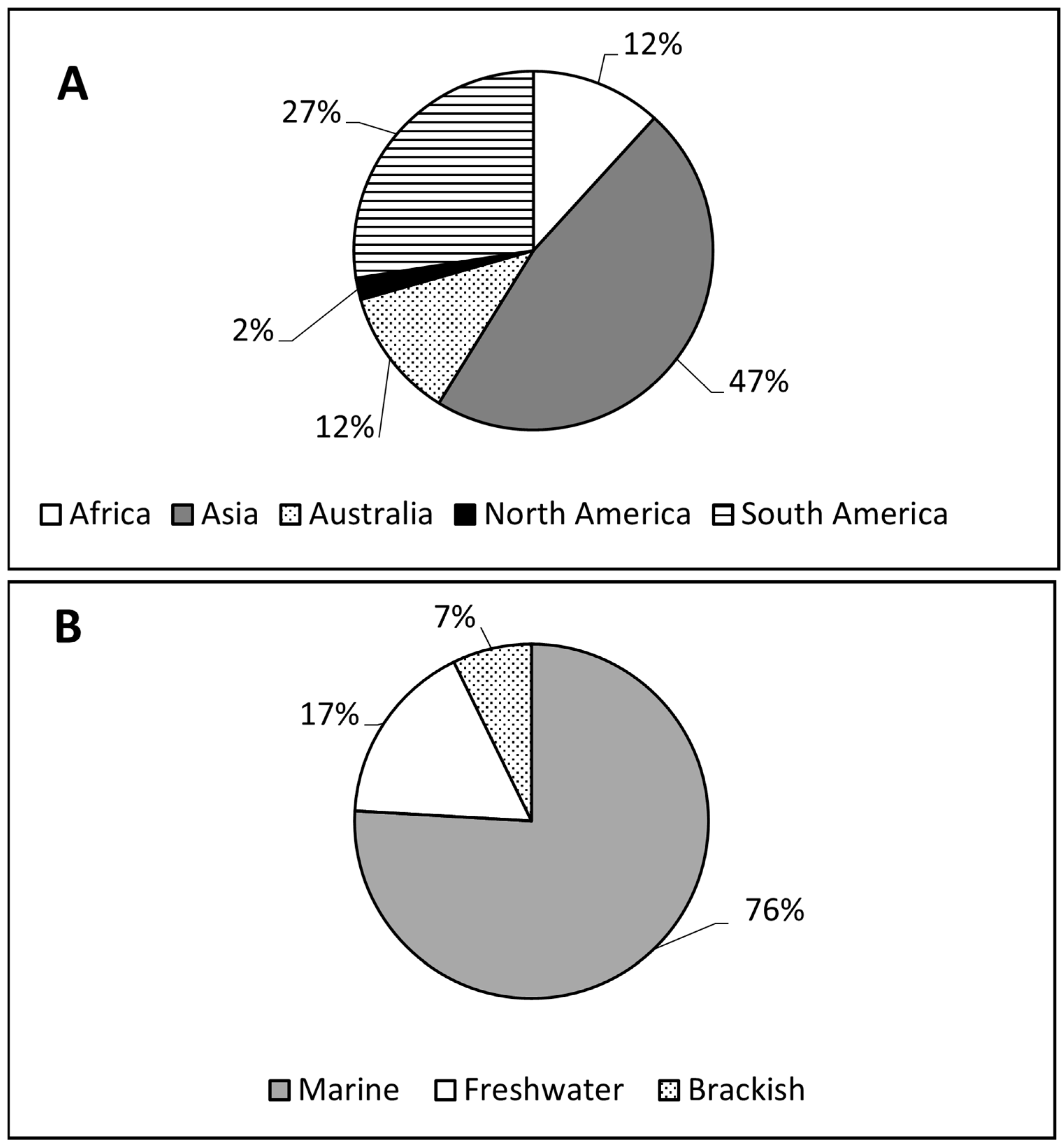
3.1. Under-Representation of Phytoplankton Research in the Tropics
3.2. Addressing the Under-Representation of Phytoplankton Research in the Tropics
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and Threats of Contamination on Aquatic Ecosystems. In Bioremediation and Biotechnology; Hakeem, K.R., Bhat, R.A., Qadri, H., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–26. ISBN 978-3-030-35690-3. [Google Scholar]
- Grizzetti, B.; Lanzanova, D.; Liquete, C.; Reynaud, A.; Cardoso, A.C. Assessing water ecosystem services for water resource management. Environ. Sci. Policy 2016, 61, 194–203. [Google Scholar] [CrossRef]
- Chapman, P.M. Assessing and managing stressors in a changing marine environment. Mar. Pollut. Bull. 2017, 124, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Jones, K.R.; Klein, C.J.; Halpern, B.S.; Venter, O.; Grantham, H.; Kuempel, C.D.; Shumway, N.; Friedlander, A.M.; Possingham, H.P.; Watson, J.E.M. The Location and Protection Status of Earth’s Diminishing Marine Wilderness. Curr. Biol. 2018, 28, 2506–2512.e3. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B. Marine ecosystem services. Curr. Biol. 2017, 27, R507–R510. [Google Scholar] [CrossRef]
- Freeman, L.A.; Corbett, D.R.; Fitzgerald, A.M.; Lemley, D.A.; Quigg, A.; Steppe, C.N. Impacts of Urbanization and Development on Estuarine Ecosystems and Water Quality. Estuaries Coasts 2019, 42, 1821–1838. [Google Scholar] [CrossRef]
- WIOMSA/IOC UNESCO. United Nations Ocean Decade for Africa: The Science We Need for the Ocean We Want in Africa; WIOMSA: Zanzibar, Tanzania, 2022. [Google Scholar]
- IPCC. The Ocean and Cryosphere in a Changing Climate: Special Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: Cambridge, UK, 2022; ISBN 978-1-00-915796-4. [Google Scholar]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2023; p. 2391. [Google Scholar] [CrossRef]
- Emori, S.; Taylor, K.; Hewitson, B.; Zermoglio, F.; Juckes, M.; Lautenschlager, M.; Stockhause, M. CMIP5 Data Provided at the IPCC Data Distribution Centre. Fact Sheet of the Task Group on Data and Scenario Support for Impact and Climate Analysis (TGICA) of the Intergovernmental Panel on Climate Change (IPCC), 8p. 2016. Available online: https://www.ipcc.ch/site/assets/uploads/2020/11/TGICA_Fact_Sheet_CMIP5_data_provided_at_the_IPCC_DDC_Ver_1_2016.pdf (accessed on 30 October 2023).
- Gattuso, J.-P.; Magnan, A.; Billé, R.; Cheung, W.W.L.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M.; et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, aac4722. [Google Scholar] [CrossRef]
- Hernández Ruiz, L.; Ekumah, B.; Asiedu, D.A.; Albani, G.; Acheampong, E.; Jónasdóttir, S.H.; Koski, M.; Nielsen, T.G. Climate change and oil pollution: A dangerous cocktail for tropical zooplankton. Aquat. Toxicol. 2021, 231, 105718. [Google Scholar] [CrossRef]
- Prakash, S. Impact of Climate Change on Aquatic Ecosystem and Its Biodiversity: An Overview. Int. J. Biol. Innov. 2021, 3, 312–317. [Google Scholar] [CrossRef]
- Wernberg, T.; Smale, D.A.; Thomsen, M.S. A decade of climate change experiments on marine organisms: Procedures, patterns and problems. Glob. Chang. Biol. 2012, 18, 1491–1498. [Google Scholar] [CrossRef]
- Kordas, R.L.; Harley, C.D.G.; O’Connor, M.I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. J. Exp. Mar. Biol. Ecol. 2011, 400, 218–226. [Google Scholar] [CrossRef]
- Nguyen, K.D.T.; Morley, S.A.; Lai, C.-H.; Clark, M.S.; Tan, K.S.; Bates, A.E.; Peck, L.S. Upper Temperature Limits of Tropical Marine Ectotherms: Global Warming Implications. PLoS ONE 2011, 6, e29340. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.D.G.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The impacts of climate change in coastal marine systems: Climate change in coastal marine systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Schaum, C.-E.; Buckling, A.; Smirnoff, N.; Studholme, D.J.; Yvon-Durocher, G. Environmental fluctuations accelerate molecular evolution of thermal tolerance in a marine diatom. Nat. Commun. 2018, 9, 1719. [Google Scholar] [CrossRef]
- Dunne, J.P. Fall and rise of the phytoplankton. Nat. Clim. Chang. 2022, 12, 708–709. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Laws, E.A.; Barber, R.T.; Murray, J.W. Phytoplankton and Their Role in Primary, New, and Export Production. In Ocean Biogeochemistry; Fasham, M.J.R., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 99–121. ISBN 978-3-642-62691-3. [Google Scholar]
- Hedges, J.I. Why Dissolved Organics Matter. In Biogeochemistry of Marine Dissolved Organic Matter; Elsevier: Amsterdam, The Netherlands, 2002; pp. 1–33. ISBN 978-0-12-323841-2. [Google Scholar]
- Nebbioso, A.; Piccolo, A. Molecular characterization of dissolved organic matter (DOM): A critical review. Anal. Bioanal. Chem. 2013, 405, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P. Ocean Science: The power of plankton. Nature 2012, 483, S17–S20. [Google Scholar] [CrossRef]
- Thomas, M.K.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. A Global Pattern of Thermal Adaptation in Marine Phytoplankton. Science 2012, 338, 1085–1088. [Google Scholar] [CrossRef]
- Kwok, K.W.; Leung, K.M.; Lui, G.S.; Chu, V.K.; Lam, P.K.; Morritt, D.; Maltby, L.; Brock, T.C.; Van den Brink, P.J.; Warne, M.S.J.; et al. Comparison of tropical and temperate freshwater animal species’ acute sensitivities to chemicals: Implications for deriving safe extrapolation factors: Tropical versus Temperate Species Sensitivity. Integr. Environ. Assess. Manag. 2007, 3, 49–67. [Google Scholar] [CrossRef]
- Falkowski, P.G. Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 1997, 387, 272–275. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Morán, X.A.G.; Ducklow, H. Microbial growth in the polar oceans—Role of temperature and potential impact of climate change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef]
- Beaugrand, G. Monitoring pelagic ecosystems using plankton indicators. ICES J. Mar. Sci. 2005, 62, 333–338. [Google Scholar] [CrossRef]
- Guinder, V.A.; Popovich, C.A.; Molinero, J.C.; Marcovecchio, J. Phytoplankton summer bloom dynamics in the Bahía Blanca Estuary in relation to changing environmental conditions. Cont. Shelf Res. 2013, 52, 150–158. [Google Scholar] [CrossRef]
- Irwin, A.J.; Finkel, Z.V.; Müller-Karger, F.E.; Troccoli Ghinaglia, L. Phytoplankton adapt to changing ocean environments. Proc. Natl. Acad. Sci. USA 2015, 112, 5762–5766. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.; Peters, J.; van Beusekom, J.E.E.; St. John, M.A. Interactive effects of temperature and light during deep convection: A case study on growth and condition of the diatom Thalassiosira weissflogii. ICES J. Mar. Sci. 2015, 72, 2061–2071. [Google Scholar] [CrossRef]
- Collins, S.; Rost, B.; Rynearson, T.A. Evolutionary potential of marine phytoplankton under ocean acidification. Evol. Appl. 2014, 7, 140–155. [Google Scholar] [CrossRef]
- Feng, Y.; Warner, M.E.; Zhang, Y.; Sun, J.; Fu, F.-X.; Rose, J.M.; Hutchins, D.A. Interactive effects of increased pCO2, temperature and irradiance on the marine coccolithophore Emiliania huxleyi (Prymnesiophyceae). Eur. J. Phycol. 2008, 43, 87–98. [Google Scholar] [CrossRef]
- Murphy, G.E.P.; Romanuk, T.N.; Worm, B. Cascading effects of climate change on plankton community structure. Ecol. Evol. 2020, 10, 2170–2181. [Google Scholar] [CrossRef]
- Adams, S.M.; Greeley, M.S. Ecotoxicological Indicators of Water Quality: Using Multi-response Indicators to Assess the Health of Aquatic Ecosystems. Water Air Soil Pollut. 2000, 123, 103–115. [Google Scholar] [CrossRef]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The natural indicator of environmental pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef]
- Van De Waal, D.B.; Litchman, E. Multiple global change stressor effects on phytoplankton nutrient acquisition in a future ocean. Phil. Trans. R. Soc. B 2020, 375, 20190706. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, F.M.; Nabout, J.C.; Bini, L.M. Trends in the scientific literature on phytoplankton. Limnology 2008, 9, 153–158. [Google Scholar] [CrossRef]
- Bonachela, J.A.; Klausmeier, C.A.; Edwards, K.F.; Litchman, E.; Levin, S.A. The role of phytoplankton diversity in the emergent oceanic stoichiometry. J. Plankton Res. 2016, 38, 1021–1035. [Google Scholar] [CrossRef]
- Hülse, D.; Arndt, S.; Wilson, J.D.; Munhoven, G.; Ridgwell, A. Understanding the causes and consequences of past marine carbon cycling variability through models. Earth-Sci. Rev. 2017, 171, 349–382. [Google Scholar] [CrossRef]
- Haeder, D.-P.; Villafane, V.E.; Helbling, E.W. Productivity of aquatic primary producers under global climate change. Photochem. Photobiol. Sci. 2014, 13, 1370–1392. [Google Scholar] [CrossRef] [PubMed]
- Assmy, P.; Smetacek, V. Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Abdul, W.O.; Adekoya, E.O.; Ademolu, K.O.; Omoniyi, I.T.; Odulate, D.O.; Akindokun, T.E.; Olajide, A.E. The effects of environmental parameters on zooplankton assemblages in tropical coastal estuary, South-west, Nigeria. Egypt. J. Aquat. Res. 2016, 42, 281–287. [Google Scholar] [CrossRef]
- Bopp, S.K.; Lettieri, T. Gene regulation in the marine diatom Thalassiosira pseudonana upon exposure to polycyclic aromatic hydrocarbons (PAHs). Gene 2007, 396, 293–302. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiragushi, N.; Kitaguchi, H.; Mitsutani, A.; Nagai, S.; Yoshimura, M. A highly CO2-tolerant diatom, Thalassiosira weissflogii H1, enriched from coastal sea, and its fatty acid composition. Fish. Sci. 2000, 66, 655–659. [Google Scholar] [CrossRef]
- Vincent, F.; Bowler, C. Diatoms Are Selective Segregators in Global Ocean Planktonic Communities. mSystems 2020, 5, e00444-19. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, K.; Riebesell, U. CO2-induced seawater acidification affects physiological performance of the marine diatom Phaeodactylum tricornutum. Biogeosciences 2010, 7, 2915–2923. [Google Scholar] [CrossRef]
- Boyce, D.G.; Dowd, M.; Lewis, M.R.; Worm, B. Estimating global chlorophyll changes over the past century. Prog. Oceanogr. 2014, 122, 163–173. [Google Scholar] [CrossRef]
- Lancelot, C.; Muylaert, K. 7.02-trends in estuarine phytoplankton ecology. In Treatise on Estuarine and Coastal Science; Academic Press: Waltham, MA, USA, 2011; pp. 5–15. [Google Scholar]
- Birkle, C.; Pendlebury, D.A.; Schnell, J.; Adams, J. Web of Science as a data source for research on scientific and scholarly activity. Quant. Sci. Stud. 2020, 1, 363–376. [Google Scholar] [CrossRef]
- Conrad, E.; Christie, M.; Fazey, I. Is research keeping up with changes in landscape policy? A review of the literature. J. Environ. Manag. 2011, 92, 2097–2108. [Google Scholar] [CrossRef]
- Culumber, Z.W.; Anaya-Rojas, J.M.; Booker, W.W.; Hooks, A.P.; Lange, E.C.; Pluer, B.; Ramírez-Bullón, N.; Travis, J. Widespread Biases in Ecological and Evolutionary Studies. BioScience 2019, 69, 631–640. [Google Scholar] [CrossRef]
- Felton, A.; Fischer, J.; Lindenmayer, D.B.; Montague-Drake, R.; Lowe, A.R.; Saunders, D.; Felton, A.M.; Steffen, W.; Munro, N.T.; Youngentob, K.; et al. Climate change, conservation and management: An assessment of the peer-reviewed scientific journal literature. Biodivers. Conserv. 2009, 18, 2243–2253. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y.; Li, X.; Lai, Z.; Tackx, M.; Lek, S. A bibliometric analysis of scientific trends in phytoplankton research. Ann. Limnol.-Int. J. Limnol. 2015, 51, 249–259. [Google Scholar] [CrossRef]
- Clarke, D.A.; York, P.H.; Rasheed, M.A.; Northfield, T.D. Does Biodiversity–Ecosystem Function Literature Neglect Tropical Ecosystems? Trends Ecol. Evol. 2017, 32, 320–323. [Google Scholar] [CrossRef]
- Stroud, J.T.; Feeley, K.J. Neglect of the Tropics Is Widespread in Ecology and Evolution: A Comment on Clarke et al. Trends Ecol. Evol. 2017, 32, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Karagulle, D. World Climate Regions. Available online: https://storymaps.arcgis.com/stories/61a5d4e9494f46c2b520a984b2398f3b (accessed on 10 January 2023).
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2006; ISBN 1-139-46093-5. [Google Scholar]
- Edelman, A.; Gelding, A.; Konovalov, E.; McComiskie, R.; Penny, A.; Roberts, N.; Templeman, S.; Trewin, D.; Ziembicki, M.; Trewin, B. State of the Tropics 2014 Report; James Cook University: Townsville, Australia, 2014. [Google Scholar]
- Bloch, C.; Sorensen, M.P. The size of research funding: Trends and implications. Sci. Public Policy 2015, 42, 30–43. [Google Scholar] [CrossRef]
- Titley, M.A.; Snaddon, J.L.; Turner, E.C. Scientific research on animal biodiversity is systematically biased towards vertebrates and temperate regions. PLoS ONE 2017, 12, e0189577. [Google Scholar] [CrossRef]
- Valdés, L. Global Ocean Science Report: The Current Status of Ocean Science around the World; UNESCO: Paris, France, 2017. [Google Scholar]
- State of the Tropics. State of the Tropics 2020 Report; James Cook University: Townsville, Australia, 2020. [Google Scholar]
- Santos, J.W.; Correia, R.A.; Malhado, A.C.M.; Campos-Silva, J.V.; Teles, D.; Jepson, P.; Ladle, R.J. Drivers of taxonomic bias in conservation research: A global analysis of terrestrial mammals. Anim. Conserv. 2020, 23, 679–688. [Google Scholar] [CrossRef]
- Burian, R.M. How the Choice of Experimental Organism Matters: Epistemological Reflections on an Aspect of Biological Practice. J. Hist. Biol. 1993, 26, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, M.R.; Burnett, N.J.; Braun, D.C.; Suski, C.D.; Hinch, S.G.; Cooke, S.J.; Kerr, J.T. Taxonomic bias and international biodiversity conservation research. FACETS 2017, 1, 105–113. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Reavie, E.D. Taxonomic bias in freshwater phytoplankton communities and its effect on environmental assessment metrics. Phycologia 2022, 61, 436–443. [Google Scholar] [CrossRef]
- Tyree, M.A.; Bishop, I.W.; Hawkins, C.P.; Mitchell, R.; Spaulding, S.A. Reduction of taxonomic bias in diatom species data. Limnol. Oceanogr. Methods 2020, 18, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Kwiatkowski, L.; Bopp, L.; Aumont, O.; Ciais, P.; Cox, P.M.; Laufkötter, C.; Li, Y.; Séférian, R. Emergent constraints on projections of declining primary production in the tropical oceans. Nat. Clim. Chang. 2017, 7, 355–358. [Google Scholar] [CrossRef]
- Flombaum, P.; Wang, W.-L.; Primeau, F.W.; Martiny, A.C. Global picophytoplankton niche partitioning predicts overall positive response to ocean warming. Nat. Geosci. 2020, 13, 116–120. [Google Scholar] [CrossRef]
- Estrada, M.; Delgado, M.; Blasco, D.; Latasa, M.; Cabello, A.M.; Benítez-Barrios, V.; Fraile-Nuez, E.; Mozetič, P.; Vidal, M. Phytoplankton across Tropical and Subtropical Regions of the Atlantic, Indian and Pacific Oceans. PLoS ONE 2016, 11, e0151699. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A. do N. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef]
- Timmermann, A.; Jin, F.-F. Phytoplankton influences on tropical climate: Phytoplankton Influences on Tropical Climate. Geophys. Res. Lett. 2002, 29, 19-1–19-4. [Google Scholar] [CrossRef]
- Gorham, T.; Dowling Root, E.; Jia, Y.; Shum, C.K.; Lee, J. Relationship between cyanobacterial bloom impacted drinking water sources and hepatocellular carcinoma incidence rates. Harmful Algae 2020, 95, 101801. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.; Zhang, Y.; Li, J.; Shi, K. Research Trends in the Remote Sensing of Phytoplankton Blooms: Results from Bibliometrics. Remote Sens. 2021, 13, 4414. [Google Scholar] [CrossRef]
- Schlüter, M.; Mcallister, R.R.J.; Arlinghaus, R.; Bunnefeld, N.; Eisenack, K.; Hölker, F.; Milner-Gulland, E.J.; Müller, B.; Nicholson, E.; Quaas, M.; et al. New horizons for managing the environment: A review of coupled social-ecological systems modelling: Social-ecological systems modelling. Nat. Resour. Model. 2012, 25, 219–272. [Google Scholar] [CrossRef]
- Vári, Á.; Podschun, S.A.; Erős, T.; Hein, T.; Pataki, B.; Iojă, I.-C.; Adamescu, C.M.; Gerhardt, A.; Gruber, T.; Dedić, A.; et al. Freshwater systems and ecosystem services: Challenges and chances for cross-fertilization of disciplines. Ambio 2022, 51, 135–151. [Google Scholar] [CrossRef]
- Jacobs, Z.L.; Yool, A.; Jebri, F.; Srokosz, M.; van Gennip, S.; Kelly, S.J.; Roberts, M.; Sauer, W.; Queirós, A.M.; Osuka, K.E.; et al. Key climate change stressors of marine ecosystems along the path of the East African coastal current. Ocean Coast. Manag. 2021, 208, 105627. [Google Scholar] [CrossRef]
- Saatchi, S.; Longo, M.; Xu, L.; Yang, Y.; Abe, H.; André, M.; Aukema, J.E.; Carvalhais, N.; Cadillo-Quiroz, H.; Cerbu, G.A.; et al. Detecting vulnerability of humid tropical forests to multiple stressors. One Earth 2021, 4, 988–1003. [Google Scholar] [CrossRef]
- Frölicher, T.L.; Rodgers, K.B.; Stock, C.A.; Cheung, W.W.L. Sources of uncertainties in 21st century projections of potential ocean ecosystem stressors. Glob. Biogeochem. Cycles 2016, 30, 1224–1243. [Google Scholar] [CrossRef]
- Rodgers, K.B.; Lin, J.; Frölicher, T.L. Emergence of multiple ocean ecosystem drivers in a large ensemble suite with an Earth system model. Biogeosciences 2015, 12, 3301–3320. [Google Scholar] [CrossRef]
- Arnosti, C.; Steen, A.D.; Ziervogel, K.; Ghobrial, S.; Jeffrey, W.H. Latitudinal Gradients in Degradation of Marine Dissolved Organic Carbon. PLoS ONE 2011, 6, e28900. [Google Scholar] [CrossRef]
- Brunskill, G.J. Tropical Margins. In Carbon and Nutrient Fluxes in Continental Margins; Liu, K.-K., Atkinson, L., Quiñones, R., Talaue-McManus, L., Eds.; Global Change–The IGBP Series; Springer: Berlin/Heidelberg, Germany, 2010; pp. 423–493. ISBN 978-3-540-92734-1. [Google Scholar]
- Lønborg, C.; Müller, M.; Butler, E.C.V.; Jiang, S.; Ooi, S.K.; Trinh, D.H.; Wong, P.Y.; Ali, S.M.; Cui, C.; Siong, W.B.; et al. Nutrient cycling in tropical and temperate coastal waters: Is latitude making a difference? Estuar. Coast. Shelf Sci. 2021, 262, 107571. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecological Research Tool: Challenges and Benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef]
- Nordling, L. World Bank pours hundreds of millions into African science. Nature 2018, 561, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Berger, S. Brain Drain, Brain Gain and Its Net Effect; Global Knowledge Partnerships on Migration and Development (KNOMAD): Washington, DC, USA, 2022. [Google Scholar]
- Batten, S.D.; Abu-Alhaija, R.; Chiba, S.; Edwards, M.; Graham, G.; Jyothibabu, R.; Kitchener, J.A.; Koubbi, P.; McQuatters-Gollop, A.; Muxagata, E.; et al. A Global Plankton Diversity Monitoring Program. Front. Mar. Sci. 2019, 6, 321. [Google Scholar] [CrossRef]
- Asiedu, D.A. Copepod Abundance and Impact of Selected Human Induced Stressors on Calanoid Copepod in the Coastal Sea of Ghana. Ph.D. Thesis, University of Cape Coast, Cape Coast, Ghana, 2020. [Google Scholar]
- Castilla, E.P.; Cunha, D.G.F.; Lee, F.W.F.; Loiselle, S.; Ho, K.C.; Hall, C. Quantification of phytoplankton bloom dynamics by citizen scientists in urban and peri-urban environments. Environ. Monit. Assess. 2015, 187, 690. [Google Scholar] [CrossRef]
- Esenkulova, S.; Suchy, K.D.; Pawlowicz, R.; Costa, M.; Pearsall, I.A. Harmful Algae and Oceanographic Conditions in the Strait of Georgia, Canada Based on Citizen Science Monitoring. Front. Mar. Sci. 2021, 8, 725092. [Google Scholar] [CrossRef]
- Vose, P.B.; Cervellini, A. Problems of scientific research in developing countries. IAEA Bull. 1983, 25, 37–40. [Google Scholar]
- Musar, A. Equipment for Science Education Constraints and Opportunities; World Bank: Washington, DC, USA, 1993. [Google Scholar]
- Song, K.; Mohseni, M.; Taghipour, F. Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: A review. Water Res. 2016, 94, 341–349. [Google Scholar] [CrossRef]
- Steinberg, M.K.; First, M.R.; Lemieux, E.J.; Drake, L.A.; Nelson, B.N.; Kulis, D.M.; Anderson, D.M.; Welschmeyer, N.A.; Herring, P.R. Comparison of techniques used to count single-celled viable phytoplankton. J. Appl. Phycol. 2012, 24, 751–758. [Google Scholar] [CrossRef]
- Jones, M.L.; Blunt, P. ‘Twinning’as a method of sustainable institutional capacity building. Public Adm. Dev. 1999, 19, 381–402. [Google Scholar] [CrossRef]
- Clare, B. Knowledge exchange: Plankton education and outreach activities. In Proceedings of the Book of Abstracts; VLIZ Special Publication 60–Flanders Marine Institute: Bruges, Belgium, 2012. [Google Scholar]
- Auranen, O.; Nieminen, M. University research funding and publication performance—An international comparison. Res. Policy 2010, 39, 822–834. [Google Scholar] [CrossRef]
- Deikumah, J.P.; Mcalpine, C.A.; Maron, M. Biogeographical and Taxonomic Biases in Tropical Forest Fragmentation Research: Biases in Forest Fragmentation Research. Conserv. Biol. 2014, 28, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; van Aarde, R.J. Geographical and taxonomic biases in research on biodiversity in human-modified landscapes. Ecosphere 2012, 3, art119. [Google Scholar] [CrossRef]
- Whitworth, J.A.; Kokwaro, G.; Kinyanjui, S.; Snewin, V.A.; Tanner, M.; Walport, M.; Sewankambo, N. Strengthening capacity for health research in Africa. Lancet 2008, 372, 1590–1593. [Google Scholar] [CrossRef]
- Winter, M.; Fiedler, W.; Hochachka, W.M.; Koehncke, A.; Meiri, S.; De la Riva, I. Patterns and biases in climate change research on amphibians and reptiles: A systematic review. R. Soc. Open Sci. 2016, 3, 160158. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhiambo, R.; Mensah, P.K.; Acheampong, E. Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review. Water 2023, 15, 4288. https://doi.org/10.3390/w15244288
Adhiambo R, Mensah PK, Acheampong E. Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review. Water. 2023; 15(24):4288. https://doi.org/10.3390/w15244288
Chicago/Turabian StyleAdhiambo, Rael, Paul Kojo Mensah, and Emmanuel Acheampong. 2023. "Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review" Water 15, no. 24: 4288. https://doi.org/10.3390/w15244288