
The Impact of Environmental Factors on the Spatiotemporal Heterogeneity of Phytoplankton Community Structure and Biodiversity in the Qiongzhou Strait

Abstract
:1. Introduction
2. Materials and Methods
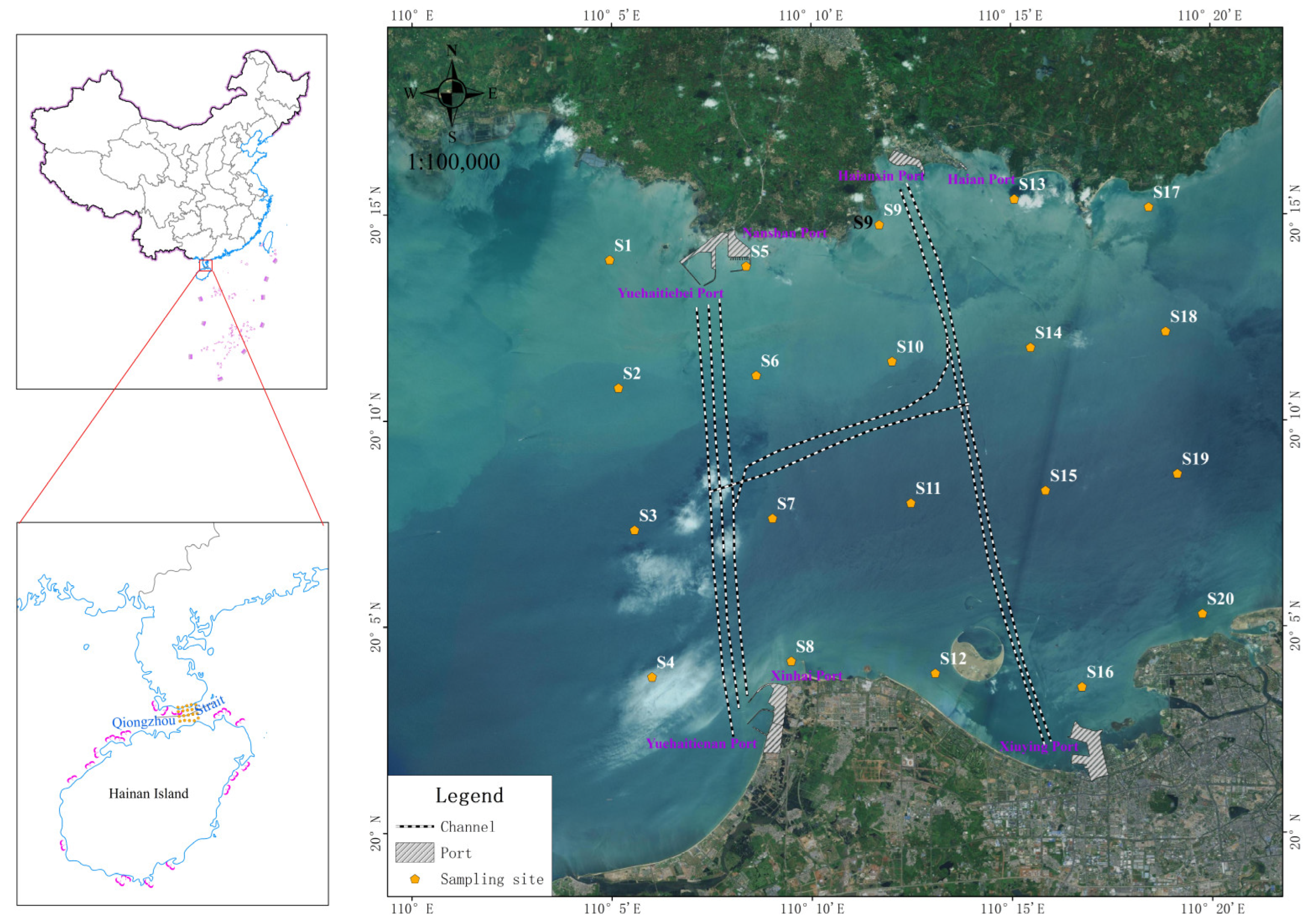
2.1. Study Area
2.2. Sample Collection
2.3. Data Analysis
3. Results
3.1. Differences in Water Quality between the Two Seasons
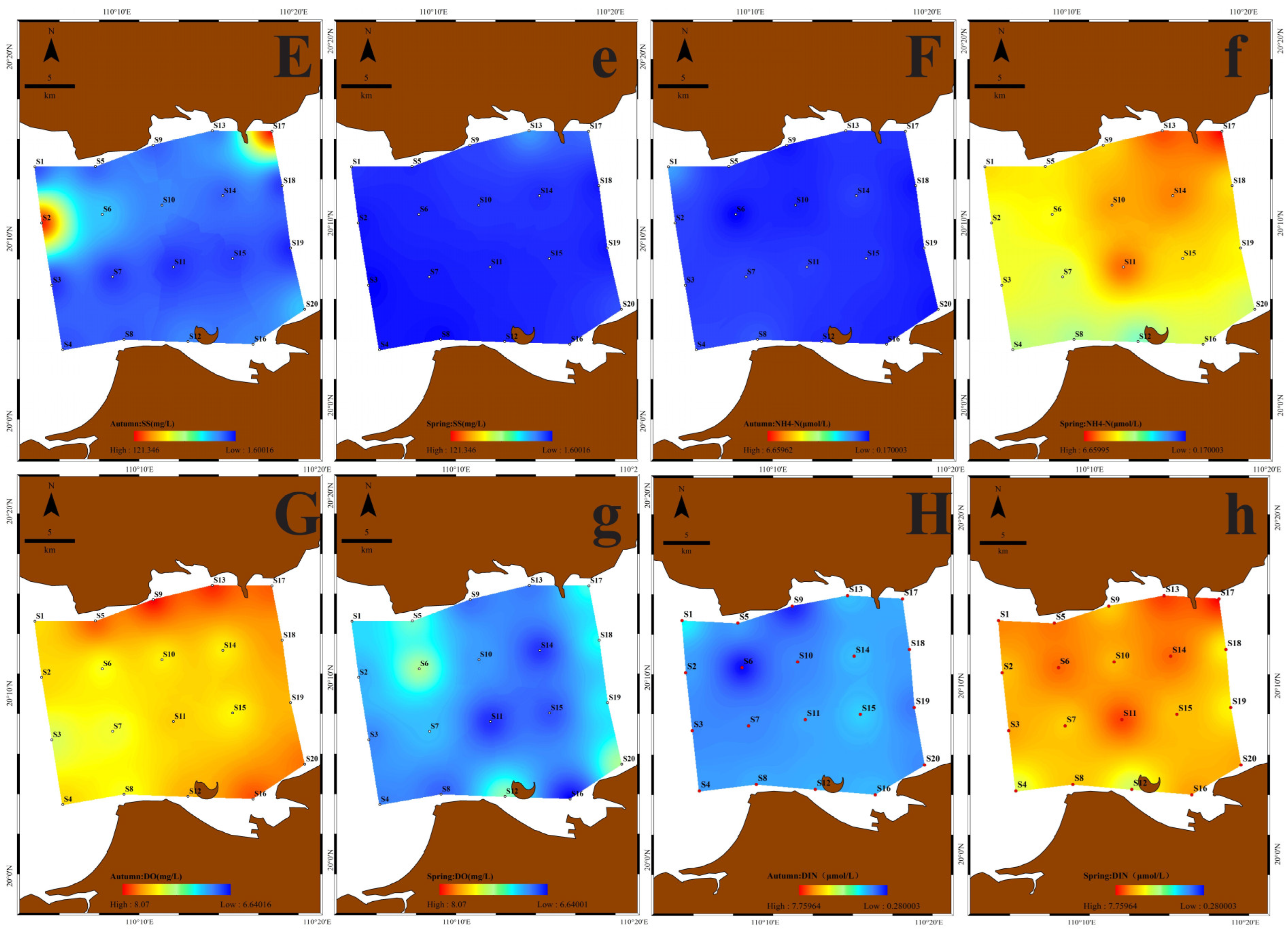
3.2. The Spatial and Temporal Distribution of Water Quality
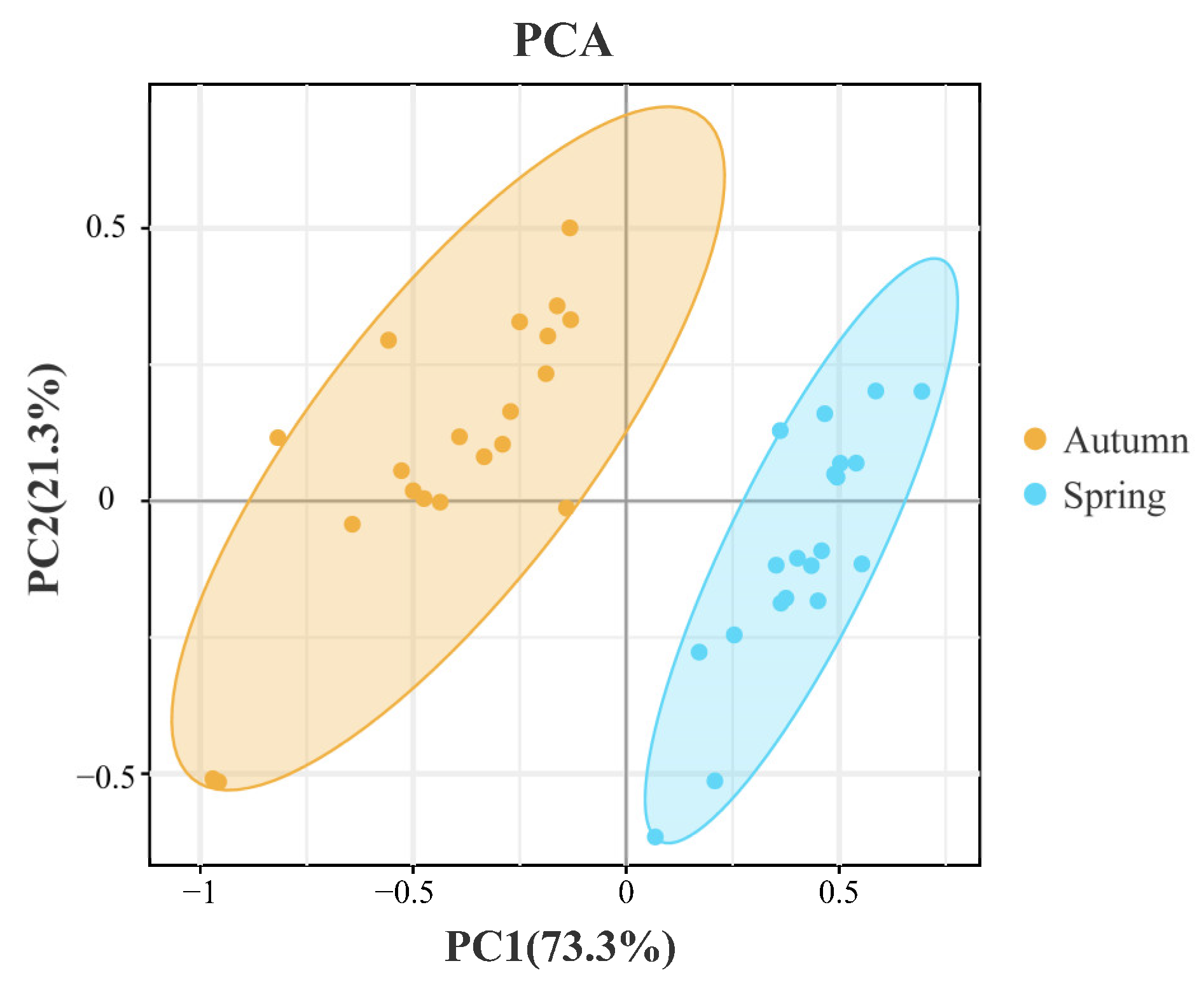
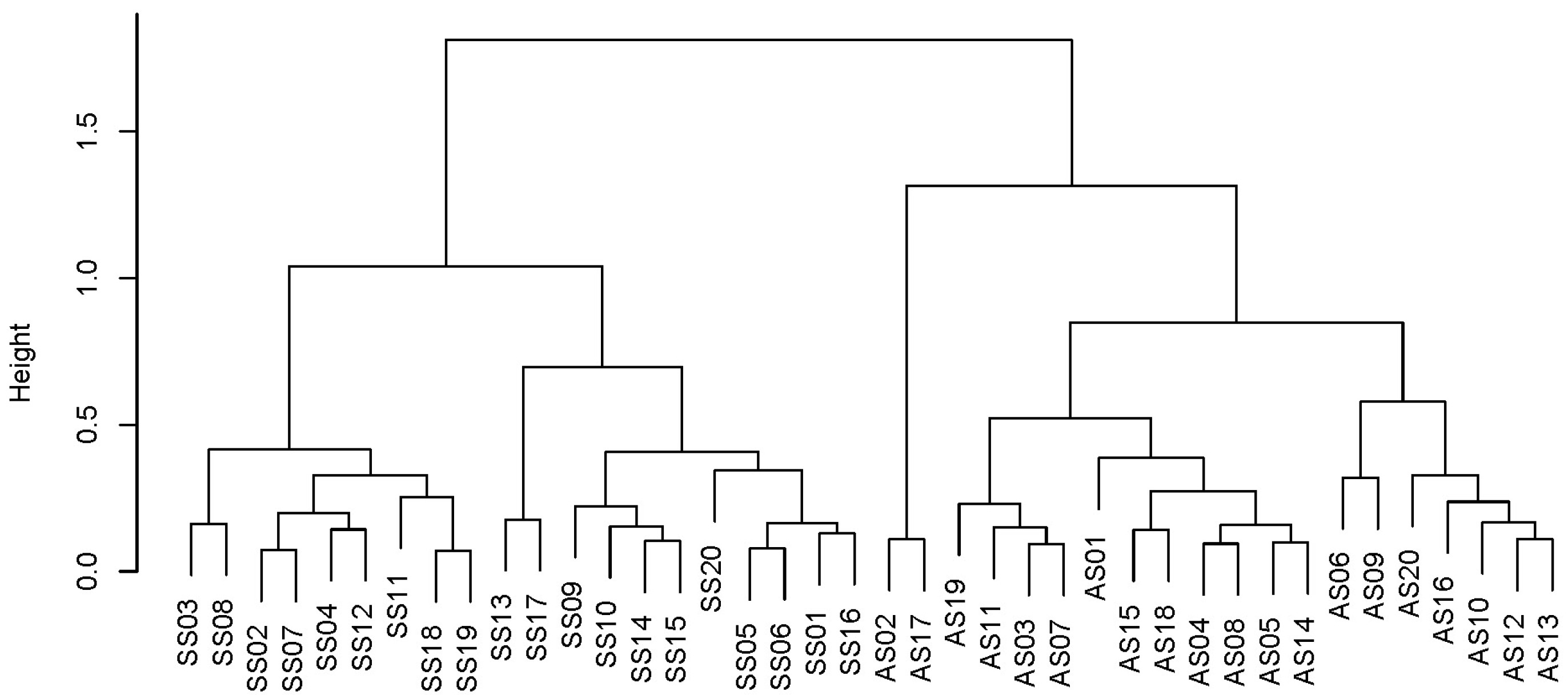
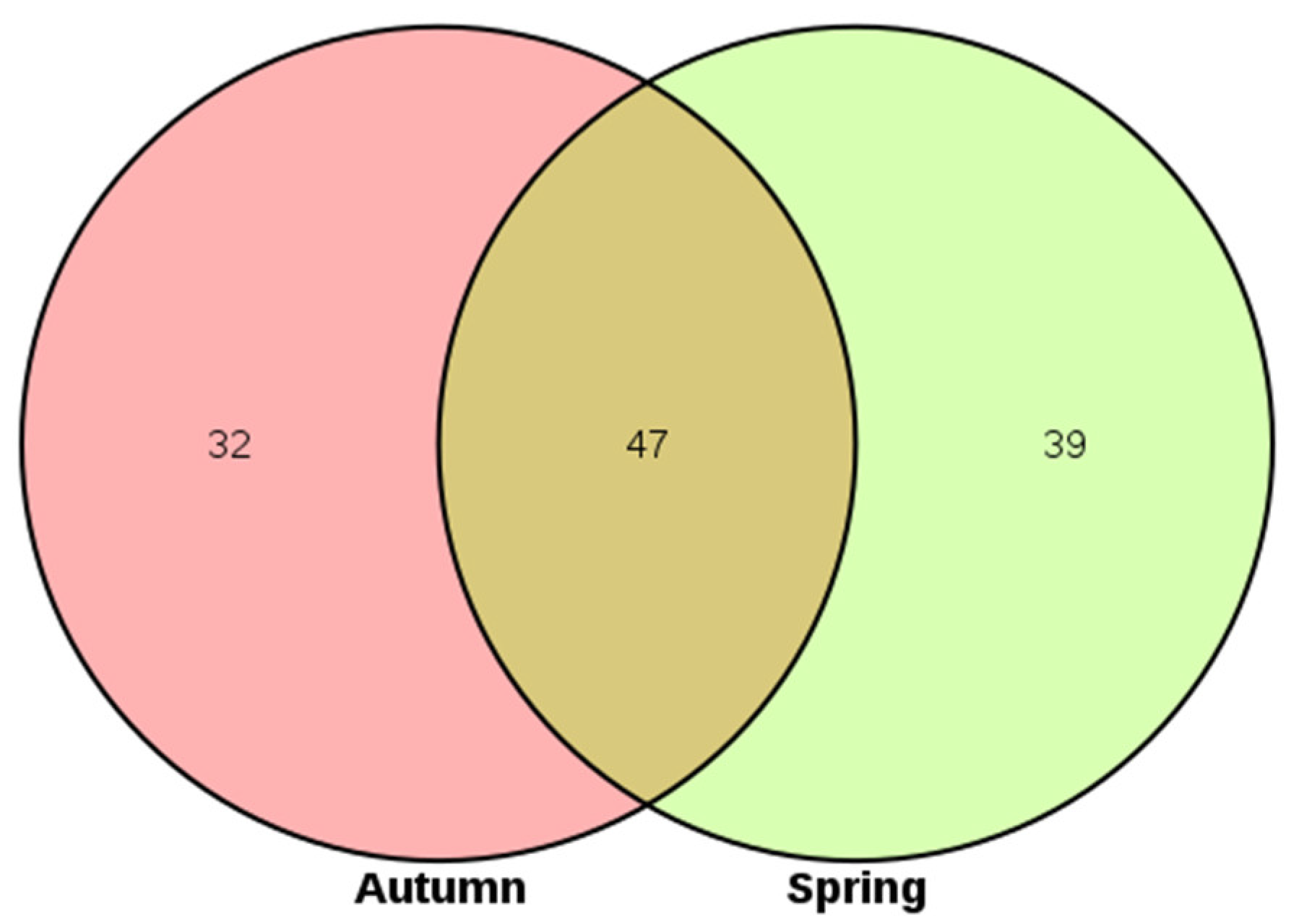
3.3. The Phytoplankton Community between the Two Seasons
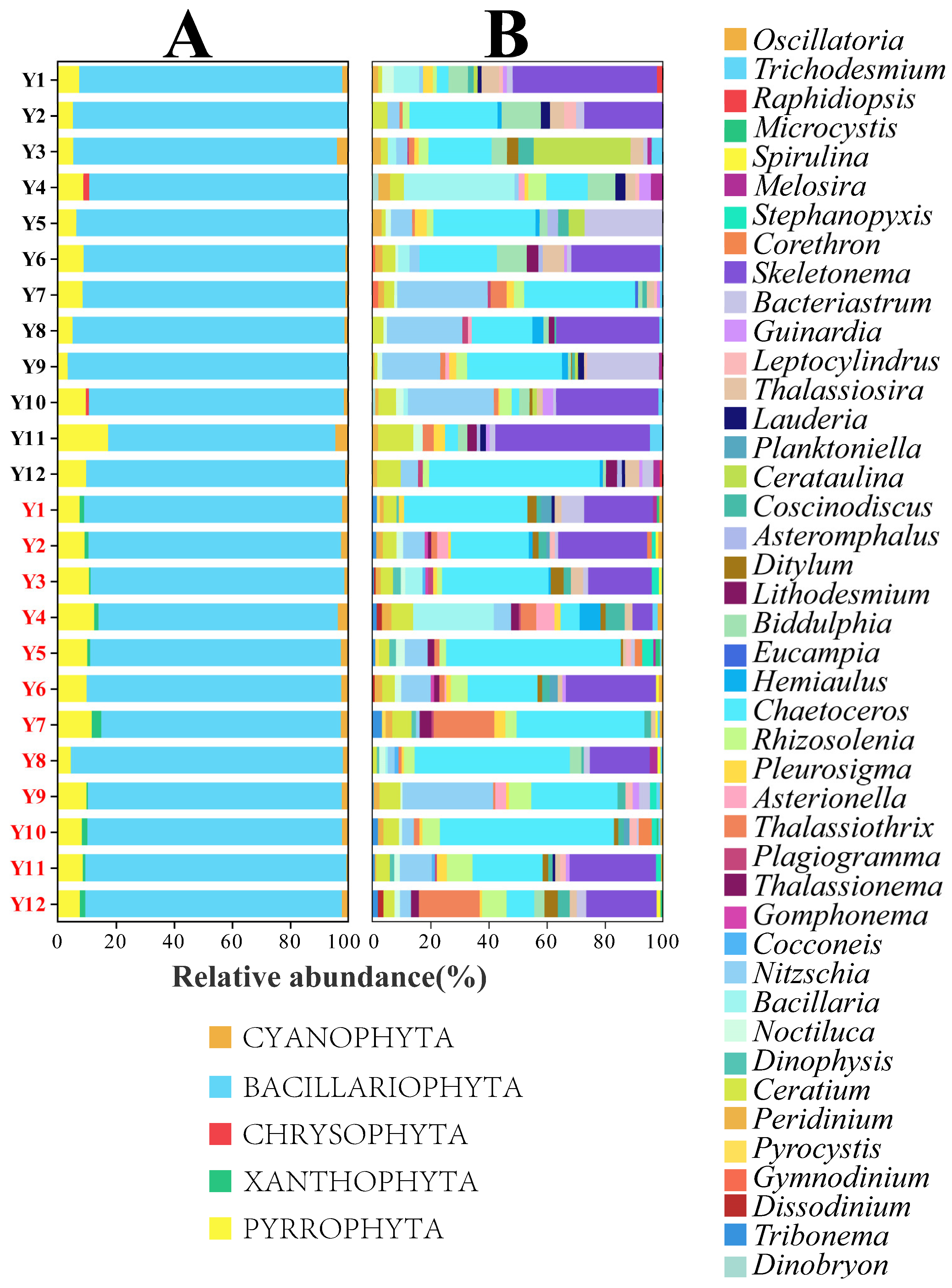
3.4. The Community Composition and Dominant Species in the Two Seasons
3.5. The Distribution of Diversity and Abundance
3.6. The Relationship between Environmental Factors and Biodiversity of Phytoplankton
4. Discussion
4.1. Hydrodynamic Factors and Artificial Activities Impacting Water Quality
4.2. Vital Environmental Factors Induced Differences in the Phytoplankton Community
4.3. Relationship between Intensively Varied Environmental Factors and Important Dominant Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, B.; Xu, Z.; Ya, H.; Chen, X.; Xu, M. Impact of the water input from the eastern Qiongzhou Strait to the Beibu Gulf on Guangxi coastal circulation. Acta Oceanol. Sin. 2019, 38, 1–11. [Google Scholar] [CrossRef]
- Shi, M.; Chen, C.; Xu, Q.; Lin, H.; Liu, G.; Wang, H.; Wang, F.; Yan, J. The Role of Qiongzhou Strait in the Seasonal Variation of the South China Sea Circulation. J. Phys. Oceanogr. 2002, 32, 103–121. [Google Scholar] [CrossRef]
- Ding, C.; Li, J.; Zhong, W.; Li, F. Observation and simulation of a heavy fog event in Qiongzhou Strait. J. Atmos. Sol.-Terr. Phys. 2022, 240, 105958. [Google Scholar] [CrossRef]
- Wang, L.; Pan, W.; Zhuang, W.; Yan, X.-H.; Klemas, V.V. Analysis of seasonal characteristics of water exchange in Beibu Gulf based on a particle tracking model. Reg. Stud. Mar. Sci. 2018, 18, 35–43. [Google Scholar] [CrossRef]
- Shao, W.; Sheng, Y.; Li, H.; Shi, J.; Ji, Q.; Tan, W.; Zuo, J. Analysis of Wave Distribution Simulated by WAVEWATCH-III Model in Typhoons Passing Beibu Gulf, China. Atmosphere 2018, 9, 265. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.; Su, F. Automatic Extraction of Offshore Platforms in Single SAR Images Based on a Dual-Step-Modified Model. Sensors 2019, 19, 231. [Google Scholar] [CrossRef]
- Jiang, P.; Fan, Z.; Pan, M.; Hu, W. Research of traffic flow saturation on waters of the coastal ship routing system. Ocean. Eng. 2022, 263, 112417. [Google Scholar] [CrossRef]
- Bauer, A.; Radziejewska, T.; Liang, K.; Kowalski, N.; Dellwig, O.; Bosselmann, K.; Stark, A.; Xia, Z.; Harff, J.; Böttcher, M.E.; et al. Regional Differences of Hydrographical and Sedimentological Properties in the Beibu Gulf, South China Sea. J. Coast. Res. 2013, 66, 49–71. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Y.; Wang, Y.; Sun, F.; Sun, C.; Jiang, Z.; Cheng, H. Influence of environmental changes on phytoplankton pattern in Daya Bay, South China Sea. Rev. De Biol. Mar. Oceanogr. 2014, 49, 323–337. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Wang, Y.; Lou, Z.; Sun, C.; Sun, S. Ecological environment changes in Daya Bay, China, from 1982 to 2004. Mar. Pollut. Bull. 2008, 56, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, Y.; Dong, J.; Sun, C.; Wang, Y.; Sun, F.; Cheng, H. Investigation of Spatial and Temporal Trends in Water Quality in Daya Bay, South China Sea. Int. J. Environ. Res. Public Health 2011, 8, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- GB 12763; Specifications for Oceanographic Survey. The Second Institute of Oceanography of the State Oceanic Administration; Standards Press of China: Beijing, China, 2008.
- Stachelek, J.; Madden, C.J. Application of inverse path distance weighting for high-density spatial mapping of coastal water quality patterns. Int. J. Geogr. Inf. Sci. 2015, 29, 1240–1250. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.M.E.; Overstreet, R.M.; et al. Emerging Marine Diseases—Climate Links and Anthropogenic Factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Liu, D.; Shi, D.; Yang, S.; Pan, W. Analysis of the spatial and temporal distributions of ecological variables and the nutrient budget in the Beibu Gulf. Acta Oceanol. Sin. 2021, 40, 14–31. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, C.; Sun, X.; Zheng, J.; Liu, Y.; Yao, X. Distribution characteristics of planktonic copipods and its influencing factors in the coastal waters off Leizhou peninsula. Oceanol. Limnol. Sin. 2022, 53, 375–383. [Google Scholar]
- Ding, Y.; Bao, X.; Yao, Z.; Zhang, C.; Wan, K.; Bao, M.; Li, R.; Shi, M. A modeling study of the characteristics and mechanism of the westward coastal current during summer in the northwestern South China Sea. Ocean Sci. J. 2017, 52, 11–30. [Google Scholar] [CrossRef]
- Gao, J.; Wu, G.; Ya, H. Review of the circulation in the Beibu Gulf, South China Sea. Cont. Shelf Res. 2017, 138, 106–119. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, G.; Guo, X.; Hu, J.; Dai, M. Reconstruction of High-Resolution Sea Surface Salinity over 2003–2020 in the South China Sea Using the Machine Learning Algorithm LightGBM Model. Remote Sens. 2022, 14, 6147. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, R.; Jiang, F.; Pan, H.; Li, J.; Yu, K.; Lai, J. Spatiotemporal Variation in Phytoplankton and Physiochemical Factors during Phaeocystis globosa Red-Tide Blooms in the Northern Beibu Gulf of China. Water 2022, 14, 1099. [Google Scholar] [CrossRef]
- Wei, S.; Fu, D.; Wang, D.; Yu, G.; Luo, Y.; Xu, H. Tracer study of CDOM for nitrogen and phosphorus pollution in the offshore Leizhou Peninsula, China. Ecol. Indic. 2023, 148, 110019. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, Y.; Peng, C.; Dai, P.; Lai, J.; Zhao, L.; Zhang, J. Spatiotemporal variation, composition of DIN and its contribution to eutrophication in coastal waters adjacent to Hainan Island, China. Reg. Stud. Mar. Sci. 2020, 37, 101332. [Google Scholar] [CrossRef]
- Ren, C.; Peng, D.; Shi, X.; Yang, S.; Zhang, P.; Zhang, J. Seasonal Total Nitrogen and Phosphorus Variation, Speciation, and Composition in the Maowei Sea Affected by Riverine Flux Input, South China Sea. Water 2022, 14, 2584. [Google Scholar] [CrossRef]
- Fernandez-Jover, D.; Sanchez-Jerez, P.; Bayle-Sempere, J.; Carratala, A.; Leon, V.M. Addition of dissolved nitrogen and dissolved organic carbon from wild fish faeces and food around Mediterranean fish farms: Implications for waste-dispersal models. J. Exp. Mar. Biol. Ecol. 2007, 340, 160–168. [Google Scholar] [CrossRef]
- Guo, Z.; Xiao, Y.; Liu, Y.; Wu, P.; Li, C. Long-Term Variations of Biogenic Elements and Nutritional Status in Daya Bay, Northern South China Sea. J. Mar. Sci. Eng. 2023, 11, 904. [Google Scholar] [CrossRef]
- Lao, Q.; Zhang, S.; Li, Z.; Chen, F.; Zhou, X.; Jin, G.; Huang, P.; Deng, Z.; Chen, C.; Zhu, Q.; et al. Quantification of the Seasonal Intrusion of Water Masses and Their Impact on Nutrients in the Beibu Gulf Using Dual Water Isotopes. J. Geophys. Res. Oceans 2022, 127, e2021JC018065. [Google Scholar] [CrossRef]
- Pan, H.; Yu, X.; Liu, D.; Shi, D.; Yang, S.; Pan, W. Marine Environmental Regionalization for the Beibu Gulf Based on a Physical-Biological Model. J. Mar. Sci. Eng. 2021, 9, 187. [Google Scholar] [CrossRef]
- Ma, R.; Xie, S.; Jia, H.; Pu, L.; Peng, L.; Ge, C. Nutritional Profile and Bioremediation of Tropical Marine Aquaculture with an Integrated Microalgae and Bacteria Symbiosis. Water Air Soil Pollut. 2022, 233, 172. [Google Scholar] [CrossRef]
- Wang, J.; Kong, F.; Geng, H.; Zhang, Q.; Yuan, Y.; Yu, R. CHEMTAX analysis of phytoplankton assemblages revealed potential indicators for blooms of haptophyte Phaeocystis globosa. Ecol. Indic. 2021, 131, 108177. [Google Scholar] [CrossRef]
- Hinga, K.R. Effects of pH on coastal marine phytoplankton. Mar. Ecol. Prog. Ser. 2002, 238, 281–300. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Y.; Cheng, H.; Sun, F.; Fei, J.; Sun, C.; Yin, J.; Zhao, H.; Wang, Y. Phytoplankton community, structure and succession delineated by partial least square regression in Daya Bay, South China Sea. Ecotoxicology 2020, 29, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Kholssi, R.; Lougraimzi, H.; Moreno-Garrido, I. Effects of global environmental change on microalgal photosynthesis, growth and their distribution. Mar. Environ. Res. 2023, 184, 105877. [Google Scholar] [CrossRef]
- Dupuis, A.P.; Hann, B.J. Warm spring and summer water temperatures in small eutrophic lakes of the Canadian prairies: Potential implications for phytoplankton and zooplankton. J. Plankton Res. 2009, 31, 489–502. [Google Scholar] [CrossRef]
- Silva, C.A.d.; Train, S.; Rodrigues, L.C. Phytoplankton assemblages in a Brazilian subtropical cascading reservoir system. Hydrobiologia 2005, 537, 99–109. [Google Scholar] [CrossRef]
- Zhang, W.; Han, S.; Zhang, D.; Shan, B.; Wei, D. Variations in dissolved oxygen and aquatic biological responses in China’s coastal seas. Environ. Res. 2023, 223, 115418. [Google Scholar] [CrossRef] [PubMed]
- Rogato, A.; Amato, A.; Iudicone, D.; Chiurazzi, M.; Ferrante, M.I.; d’Alcalà, M.R. The diatom molecular toolkit to handle nitrogen uptake. Mar. Genom. 2015, 24, 95–108. [Google Scholar] [CrossRef]
- Verity, P.G. Expansion of potentially harmful algal taxa in a Georgia Estuary (USA). Harmful Algae 2010, 9, 144–152. [Google Scholar] [CrossRef]
- Zhang, L.; Xiong, L.; Li, J.; Huang, X. Long-term changes of nutrients and biocenoses indicating the anthropogenic influences on ecosystem in Jiaozhou Bay and Daya Bay, China. Mar. Pollut. Bull. 2021, 168, 112406. [Google Scholar] [CrossRef]
- Trevisant, R.; Poggi, C.; Squartinis, A. Factors affecting diatom dynamics in the alpine lakes of Colbricon (Northern Italy): A 10-year survey. J. Limnol. 2010, 69, 199–208. [Google Scholar] [CrossRef]
- Liu, S.; Wang, T.; Lu, J.; Li, Z. Seawater quality criteria derivation and ecological risk assessment for the neonicotinoid insecticide imidacloprid in China. Mar. Pollut. Bull. 2023, 190, 114871. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Ou, L.; Dai, X.; Cui, L.; Dong, Y.; Wang, P.; Li, D.; Lu, D. An overview of Prorocentrum donghaiense blooms in China: Species identification, occurrences, ecological consequences, and factors regulating prevalence. Harmful Algae 2022, 114, 102207. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Luo, H.; He, N.; Chen, W.; Yang, F.; Huang, W.; Li, N.; Sun, L.; Peng, S. Phytoplankton Communities and Their Relationship with Environmental Factors in the Waters around Macau. Int. J. Environ. Res. Public Health 2022, 19, 7788. [Google Scholar] [CrossRef]
- Yang, S.; Liu, X. Characteristics of phytoplankton assemblages in the southern Yellow Sea, China. Mar. Pollut. Bull. 2018, 135, 562–568. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Season (Mean ± Std) | p | |
|---|---|---|---|
| Autumn (n = 20) | Spring (n = 20) | ||
| Temperature (°C) | 25.75 ± 0.42 | 23.23 ± 1.19 | 0.000 ** |
| salinity | 30.31 ± 0.67 | 31.17 ± 0.21 | 0.000 ** |
| pH | 8.09 ± 0.02 | 8.24 ± 0.02 | 0.000 ** |
| COD (mg/L) | 0.56 ± 0.21 | 0.38 ± 0.09 | 0.001 ** |
| DO (mg/L) | 7.75 ± 0.18 | 6.98 ± 0.21 | 0.000 ** |
| NO2-N (μmol/L) | 0.24 ± 0.11 | 0.26 ± 0.06 | 0.367 |
| NH4-N (μmol/L) | 0.51 ± 0.30 | 4.66 ± 0.97 | 0.000 ** |
| NO3-N (μmol/L) | 1.00 ± 0.38 | 1.30 ± 0.59 | 0.059 |
| DIN (μmol/L) | 1.75 ± 0.56 | 6.22 ± 0.86 | 0.000 ** |
| PO4-P (μmol/L) | 0.01 ± 0.01 | 0.02 ± 0.02 | 0.075 |
| SS (mg/L) | 26.40 ± 33.17 | 6.91 ± 5.83 | 0.014 * |
| Items | Season (Mean ± Std) | p | |
|---|---|---|---|
| Autumn (n = 12) | Spring (n = 12) | ||
| Phylum | 3.00 ± 0.43 | 3.83 ± 0.39 | 0.000 ** |
| Family | 10.08 ± 1.44 | 13.42 ± 1.44 | 0.000 ** |
| Genus | 15.83 ± 2.69 | 19.08 ± 1.83 | 0.002 ** |
| Species numbers | 22.92 ± 3.06 | 27.75 ± 2.34 | 0.000 ** |
| Abundance (×104 cells/m3) | 79.90 ± 19.38 | 168.57 ± 23.08 | 0.000 ** |
| Diversity | 3.28 ± 0.24 | 3.49 ± 0.17 | 0.021 * |
| Evenness | 0.73 ± 0.05 | 0.72 ± 0.03 | 0.831 |
| Dominant Species | Autumn | RA (%) | Spring | RA (%) |
|---|---|---|---|---|
| Skeletonema costatum | 0.09 * | 0.19 ± 0.22 | 0.11 * | 0.16 ± 0.13 |
| Chaetoceros curvisetus | 0.06 * | 0.11 ± 0.13 | 0.08 * | 0.12 ± 0.13 |
| Chaetoceros lorenzianus | 0.04 * | 0.07 ± 0.13 | 0.01 | 0.02 ± 0.06 |
| Chaetoceros affinis (p < 0.05) | 0.01 | 0.01 ± 0.03 | 0.07 * | 0.12 ± 0.13 |
| Nitzschia delicatissima | 0.03 * | 0.05 ± 0.08 | 0.00 | 0.01 ± 0.01 |
| Nitzschia pungens | 0.01 | 0.01 ± 0.02 | 0.03 * | 0.05 ± 0.08 |
| Bacteriastrum hyalinum | 0.03 * | 0.05 ± 0.10 | 0.00 | 0.01 ± 0.01 |
| Cerataulina compacta | 0.02 * | 0.04 ± 0.09 | / | / |
| Thalassiothrix frauenfeldii | 0.00 | 0.01 ± 0.02 | 0.03 * | 0.05 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Ma, Y.; Zhang, H.; Li, X.; Chen, Q. The Impact of Environmental Factors on the Spatiotemporal Heterogeneity of Phytoplankton Community Structure and Biodiversity in the Qiongzhou Strait. Water 2023, 15, 3792. https://doi.org/10.3390/w15213792
Xu C, Ma Y, Zhang H, Li X, Chen Q. The Impact of Environmental Factors on the Spatiotemporal Heterogeneity of Phytoplankton Community Structure and Biodiversity in the Qiongzhou Strait. Water. 2023; 15(21):3792. https://doi.org/10.3390/w15213792
Chicago/Turabian StyleXu, Chunling, Yu Ma, Hao Zhang, Xiaoming Li, and Qi Chen. 2023. "The Impact of Environmental Factors on the Spatiotemporal Heterogeneity of Phytoplankton Community Structure and Biodiversity in the Qiongzhou Strait" Water 15, no. 21: 3792. https://doi.org/10.3390/w15213792