
Effect of Subirrigation and Silicon Antitranspirant Application on Biomass Yield and Carbon Dioxide Balance of a Three-Cut Meadow

Abstract
:1. Introduction
2. Materials and Methods
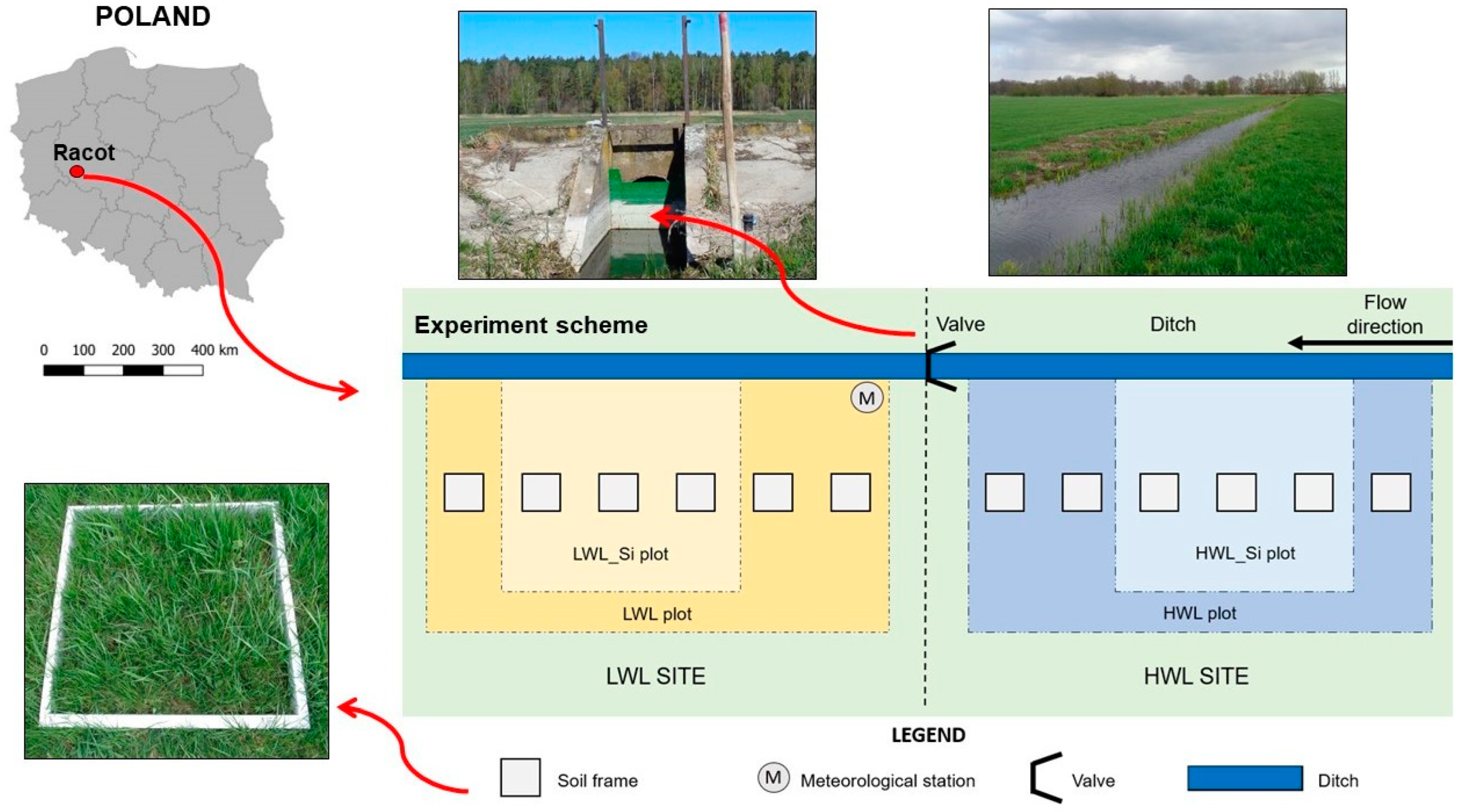
2.1. Site Description
2.2. Experimental Design
2.3. Auxiliary Data
2.4. Chamber Measurements of CO2 Fluxes
2.5. CO2 Flux Calculation and Gap Filling
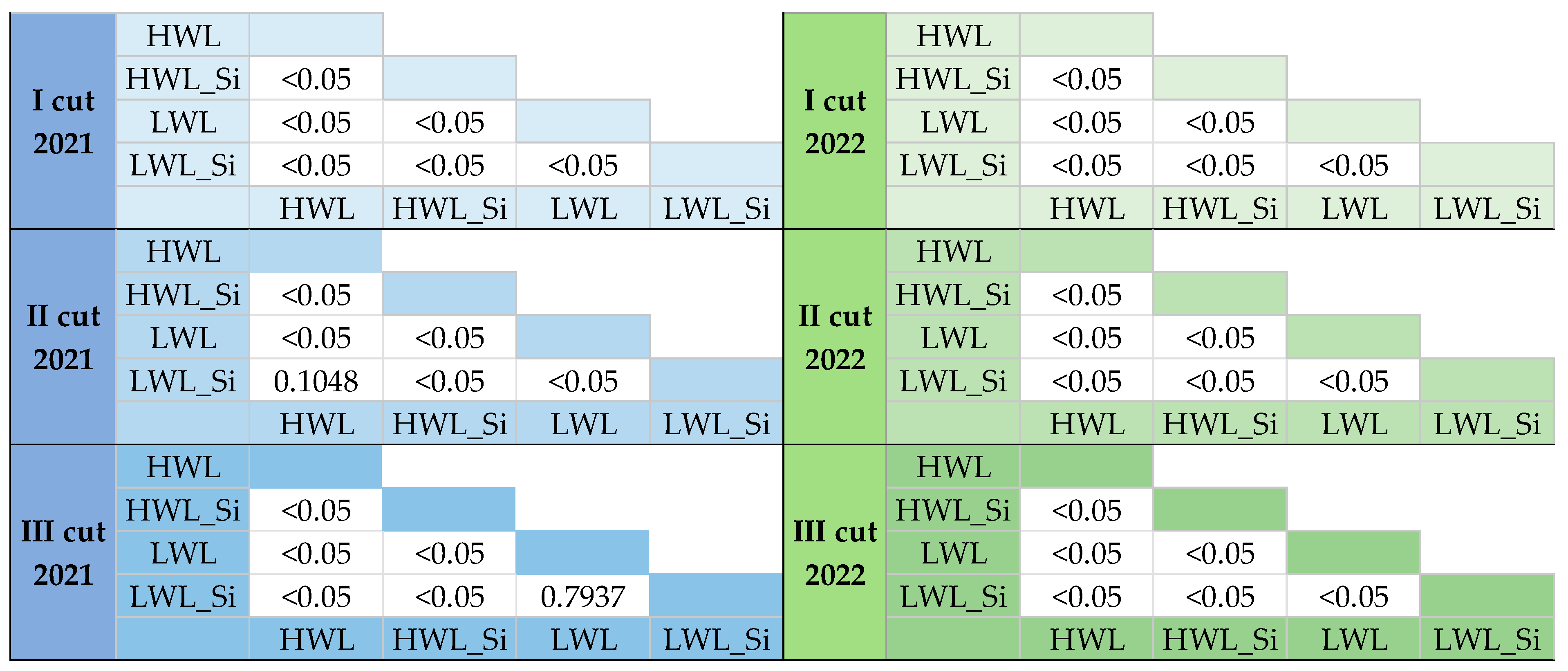
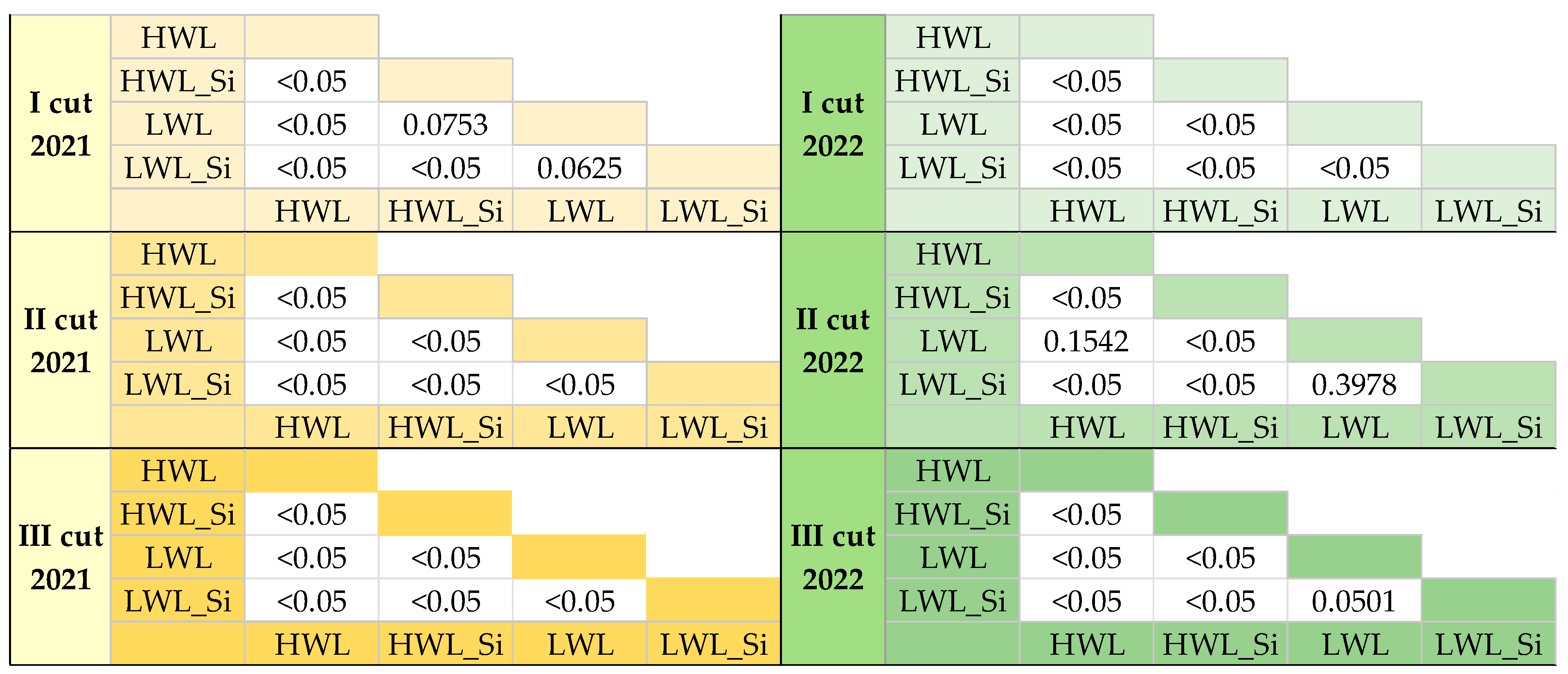
2.6. Statistical Analyses
3. Results
3.1. Environmental Conditions
3.2. GPP, Reco, and NEE Fluxes
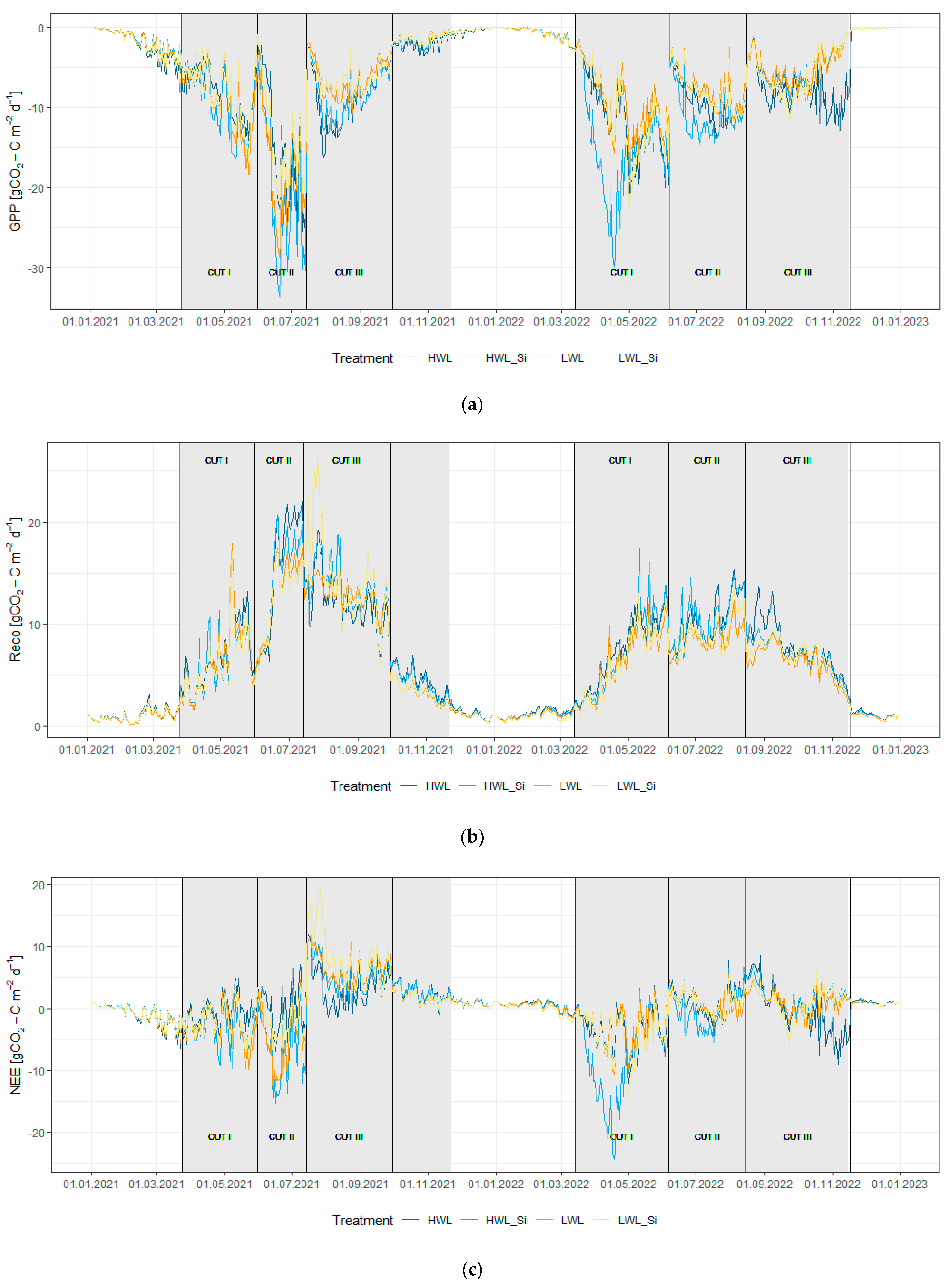
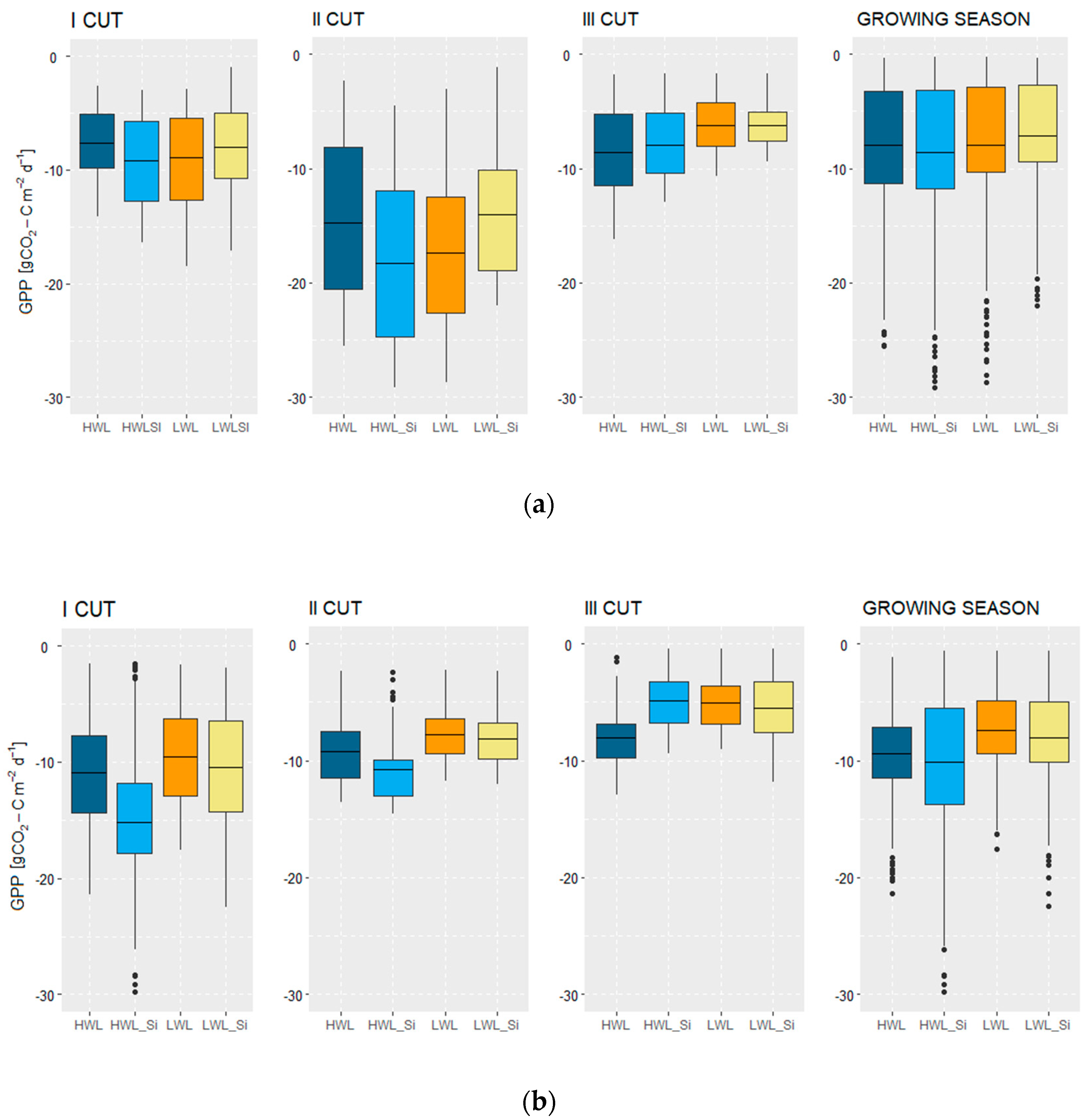
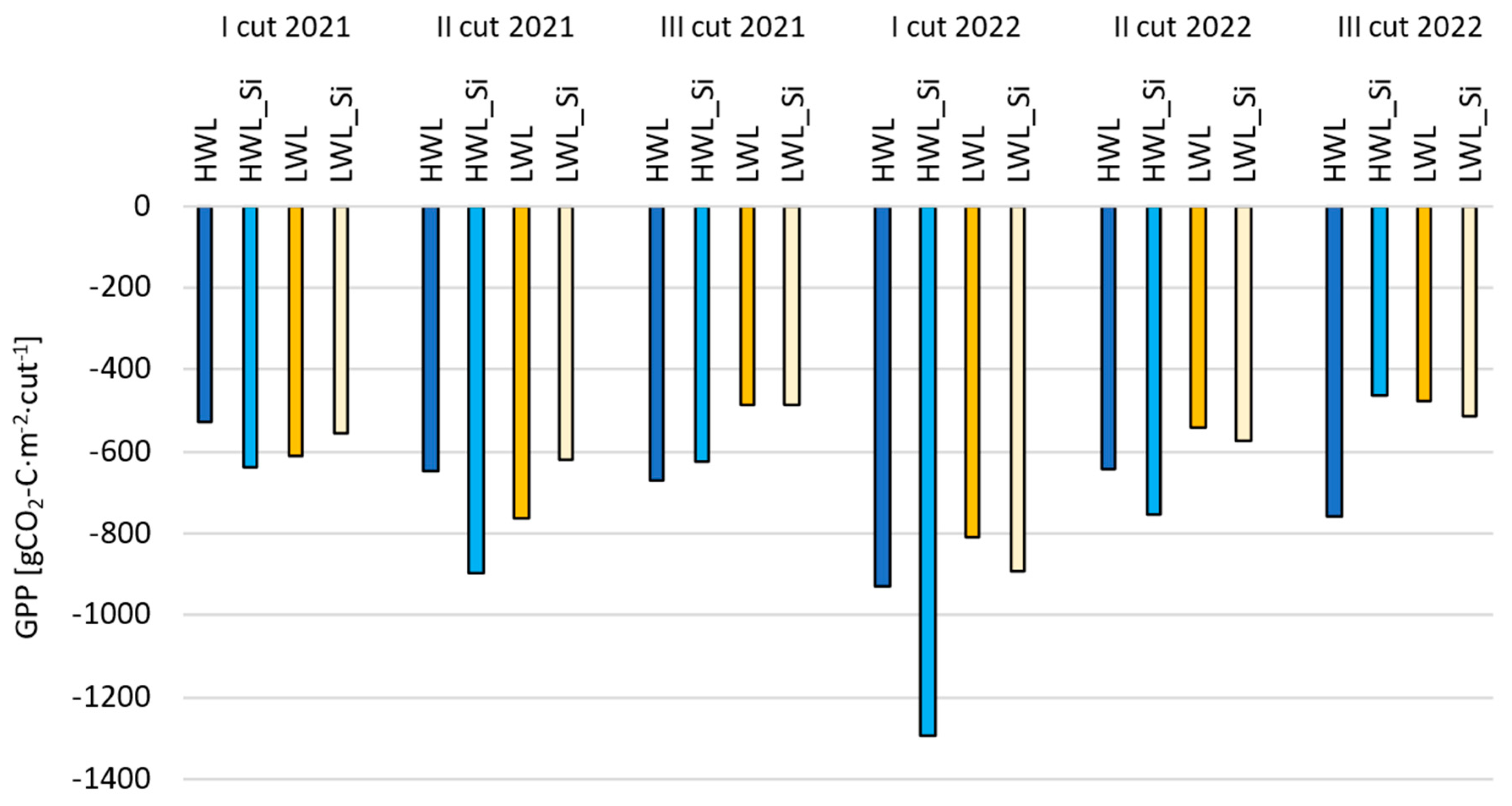
3.2.1. Gross Primary Production (GPP)
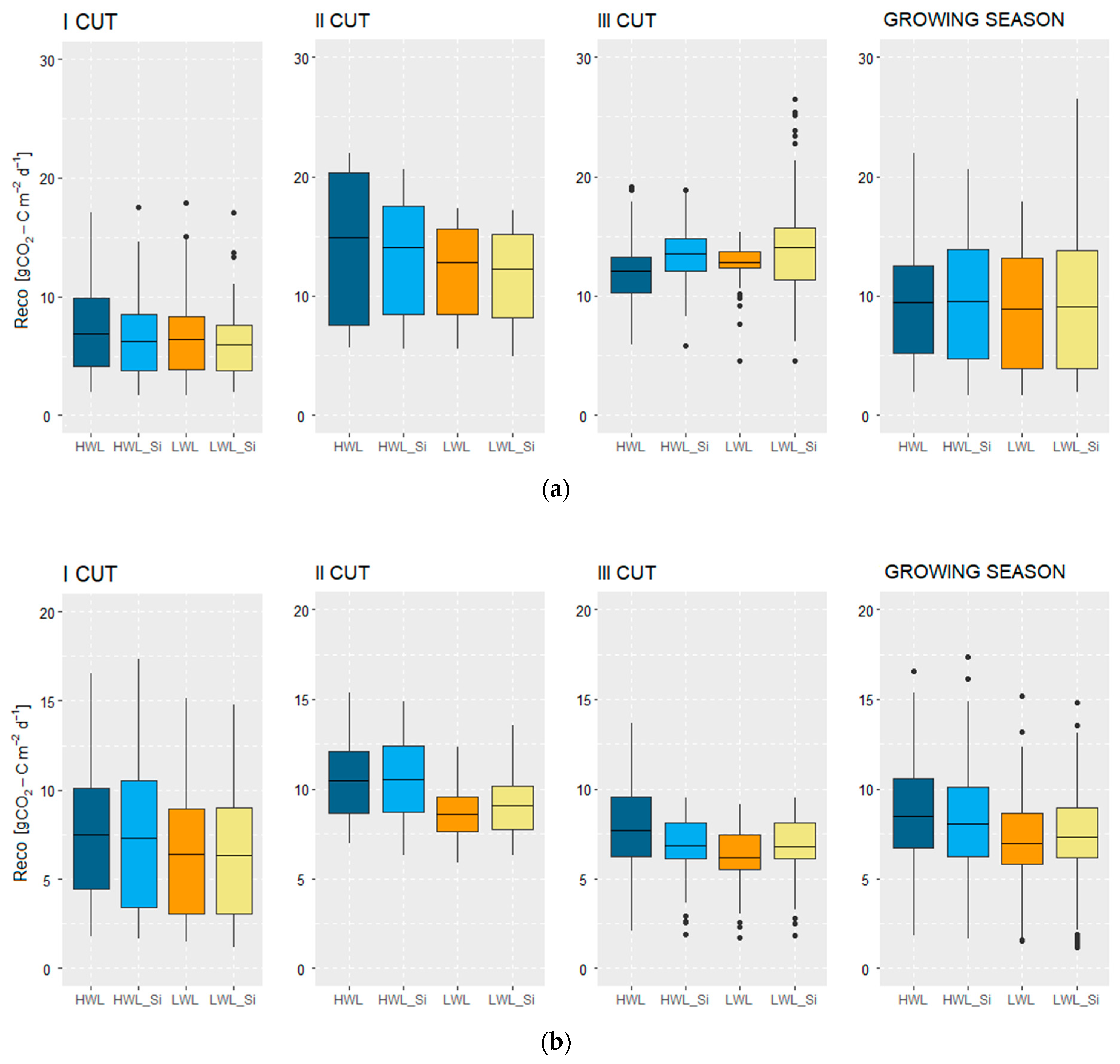
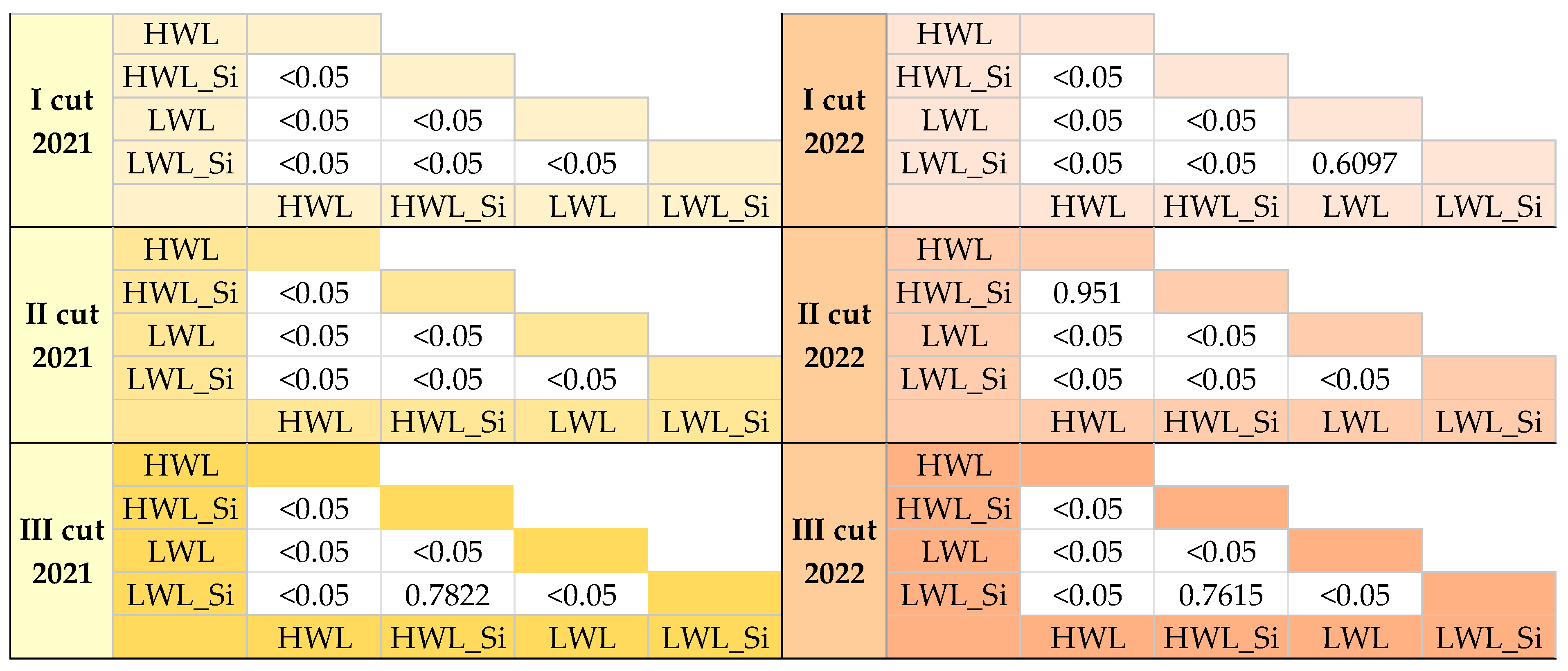
3.2.2. Ecosystem Respiration (Reco)
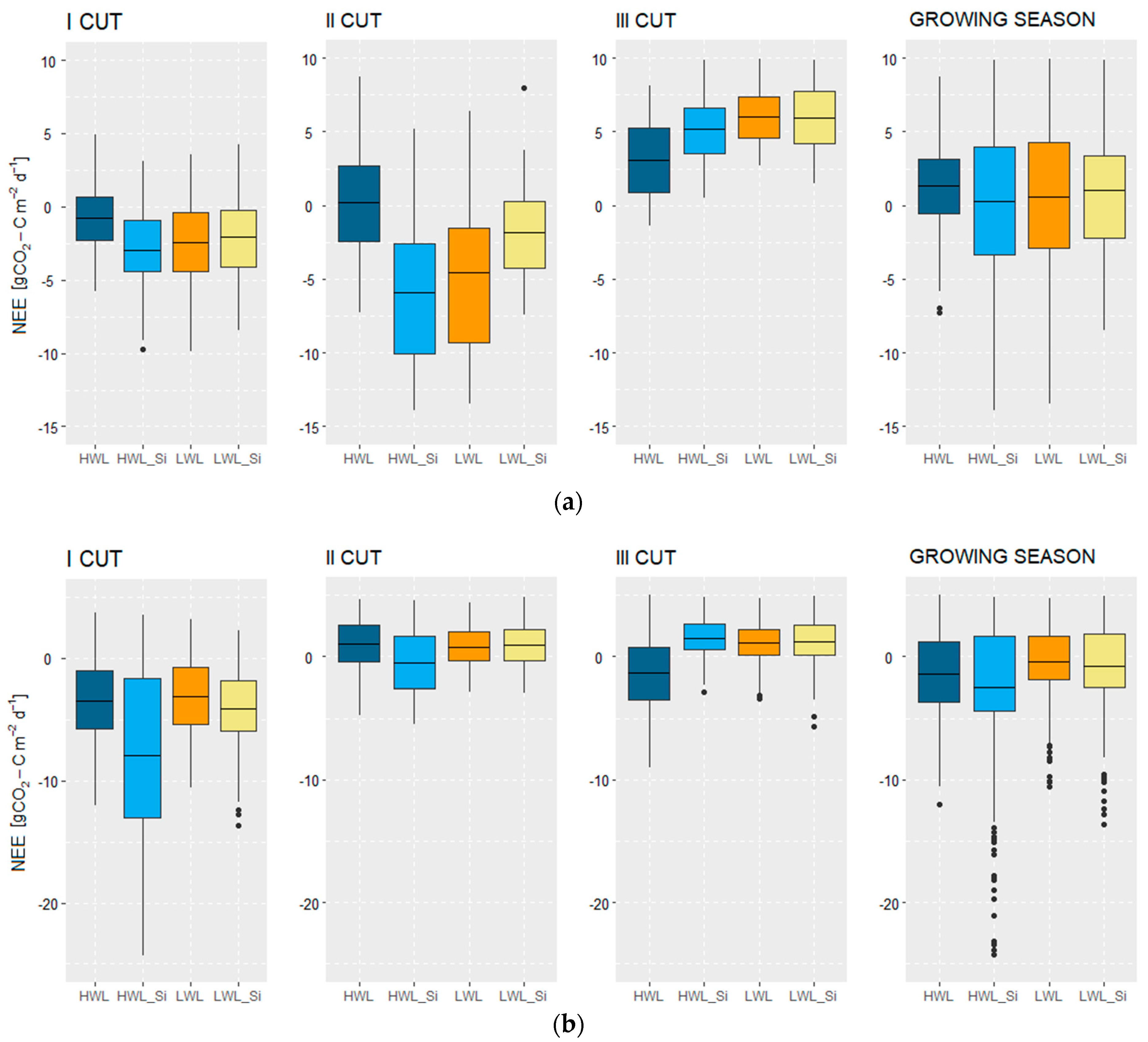
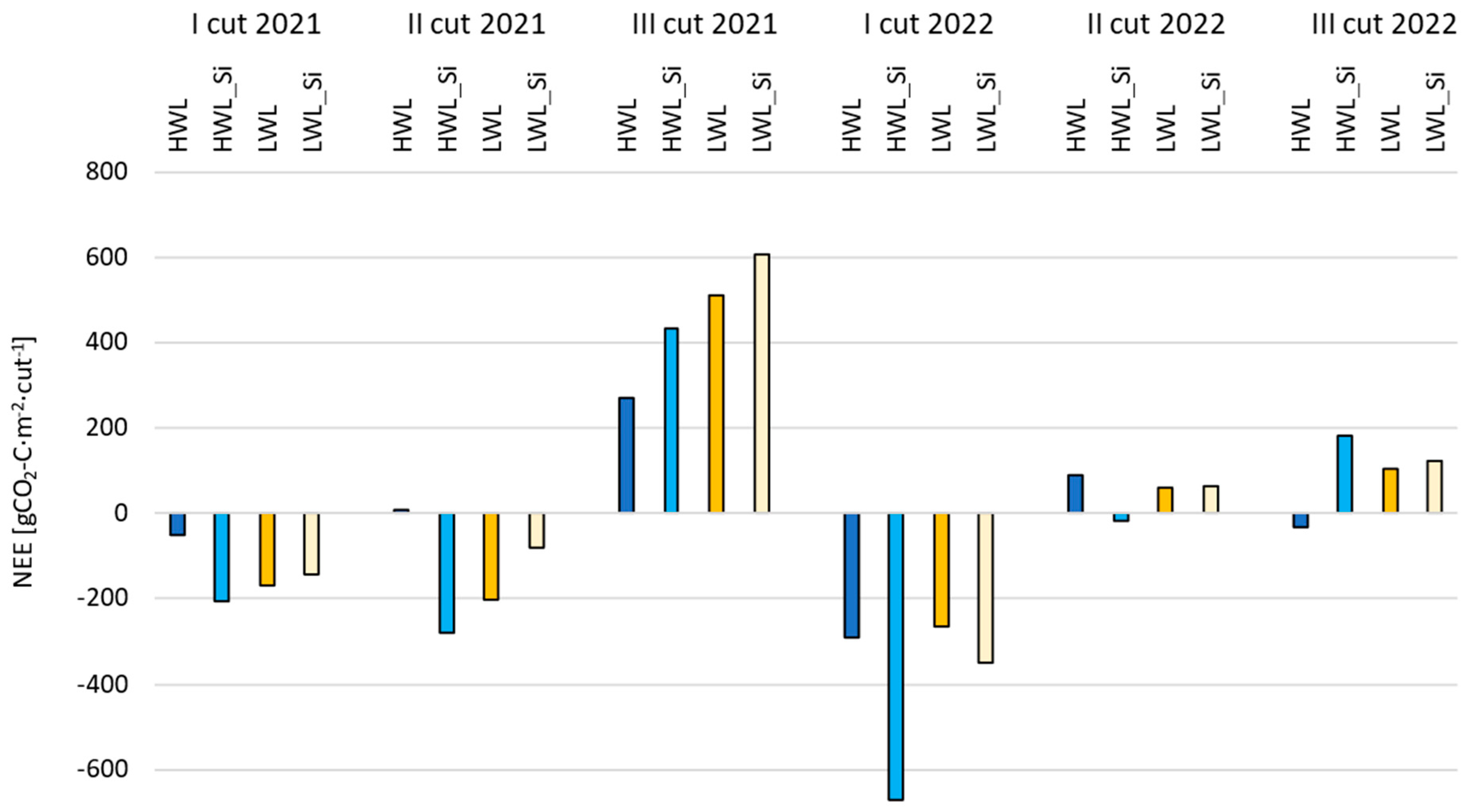
3.2.3. Net Ecosystem Exchange (NEE)
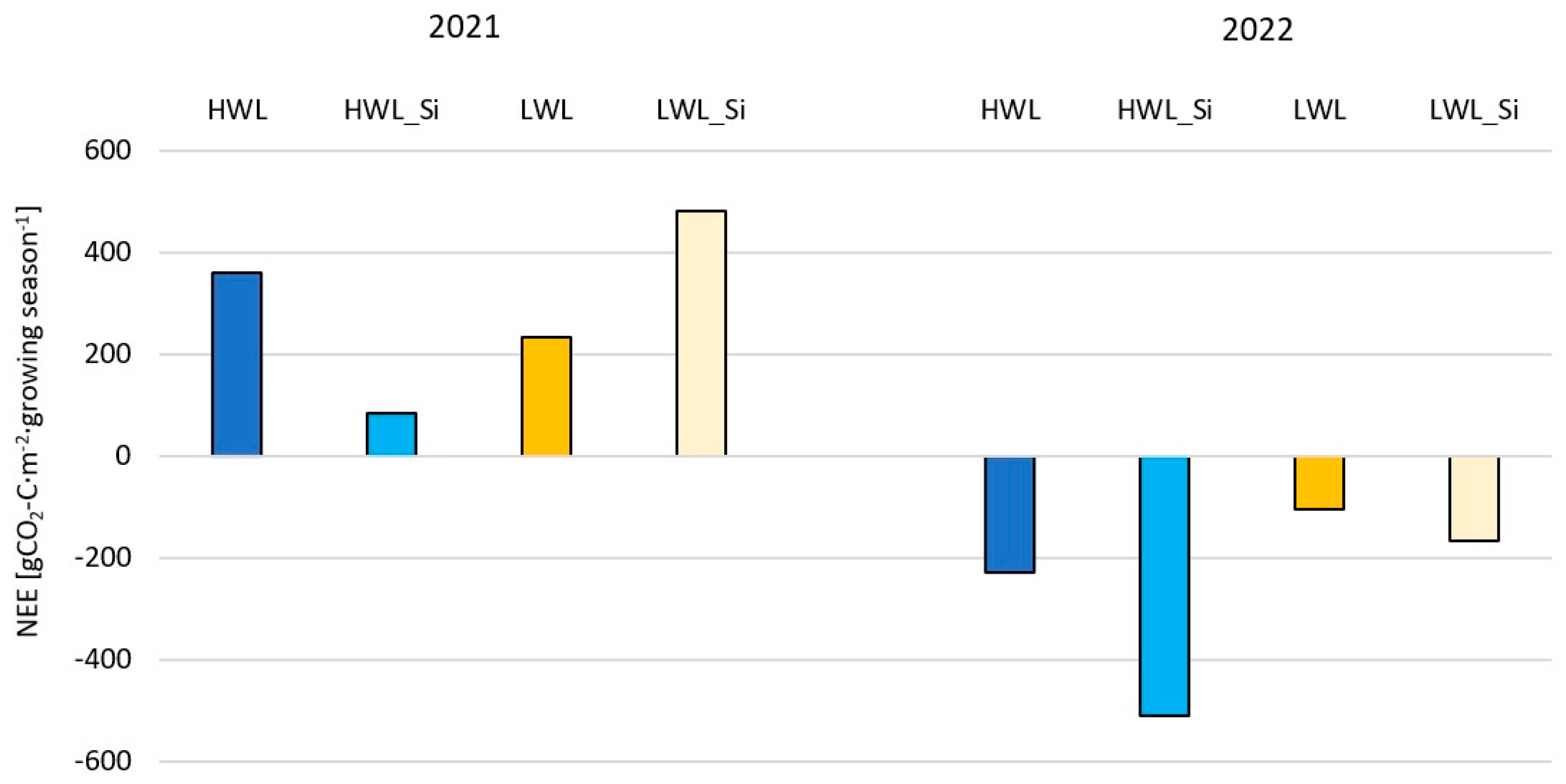
3.3. Growing Season and Annual Carbon Dioxide Balances
3.4. Meadow Yield
4. Discussion
4.1. Subirrigation and WTD Impacts on Yield and GPP
4.2. Subirrigation and WTD Impacts on CO2 Emissions
4.3. Silicon Antitranspirant Impact on Yield and GPP
4.4. Silicon Antitranspirant Impact on CO2 Emissions
4.5. Silicon Antitranspirant and Subirrigation’s Impact on Net Carbon Balances
4.6. Importance of Results and Future Research Directions
5. Conclusions
- In the drier and colder year (2021), net CO2 emissions predominated, whereas net CO2 assimilation predominated in the warmer and wetter year (2022) for all the plots, which highlights the impact of meteorological conditions on the annual NEE of grasslands.
- Higher WTD and higher soil moisture promote CO2 emissions from the meadow (Reco is higher), most probably due to an increase in the autotrophic respiration of plants due to higher aboveground biomass.
- Higher WTD and higher soil moisture promote higher yields of aboveground biomass. The yields were higher by 5.4% (in 2021) up to 11.7% (in 2022) at plot with a higher WTD, which highlights the role of the WTD in maintaining high production in meadows.
- Silicon antitranspirant application has a positive impact on meadow productivity (GPP), but only on plots with higher WTD.
- Silicon antitranspirant application has a negative impact on the yield of aboveground biomass (reduction of annual yield from 11.1% to 17.8%). The reduction of yield is higher at the plot with a higher WTD.
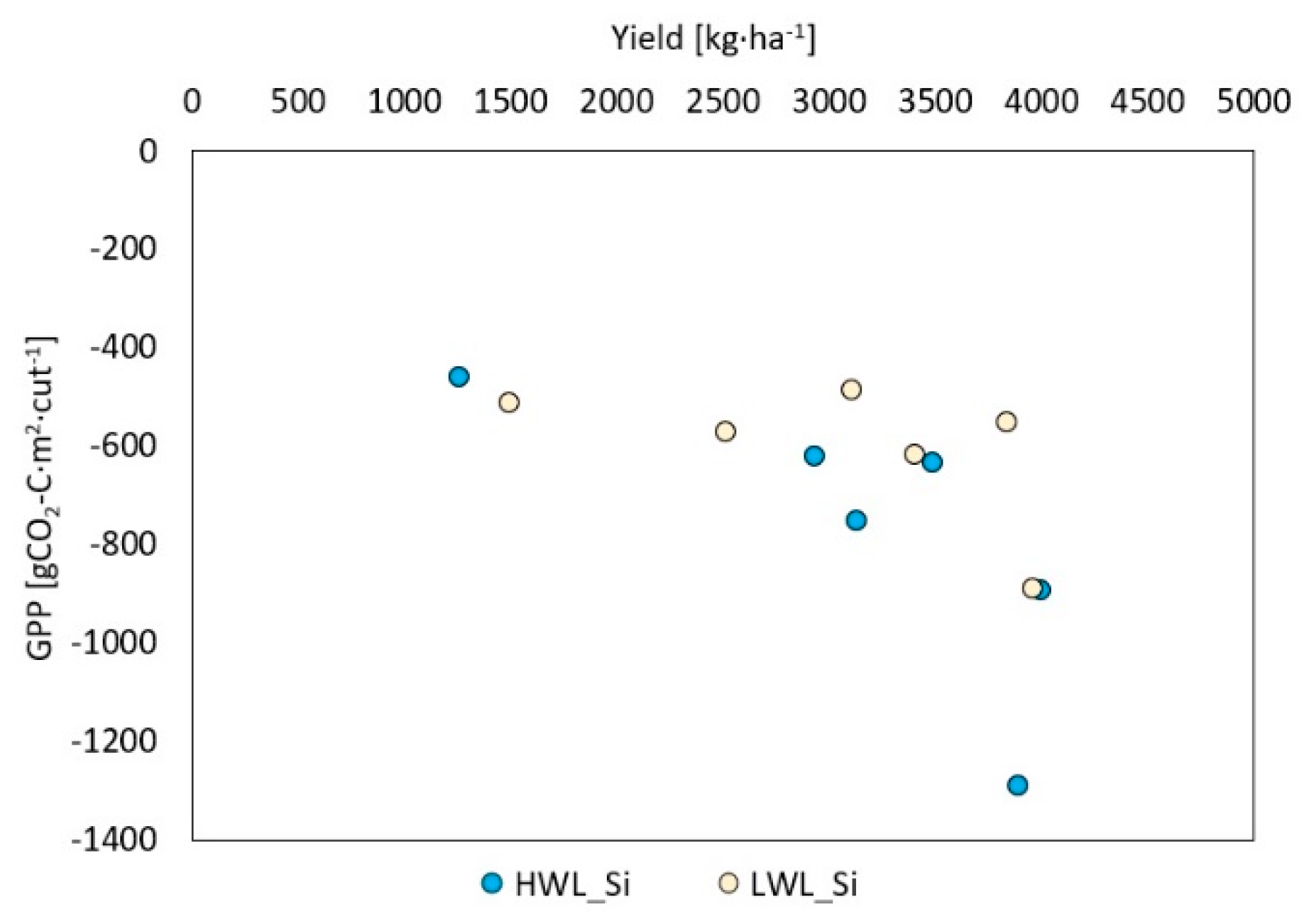
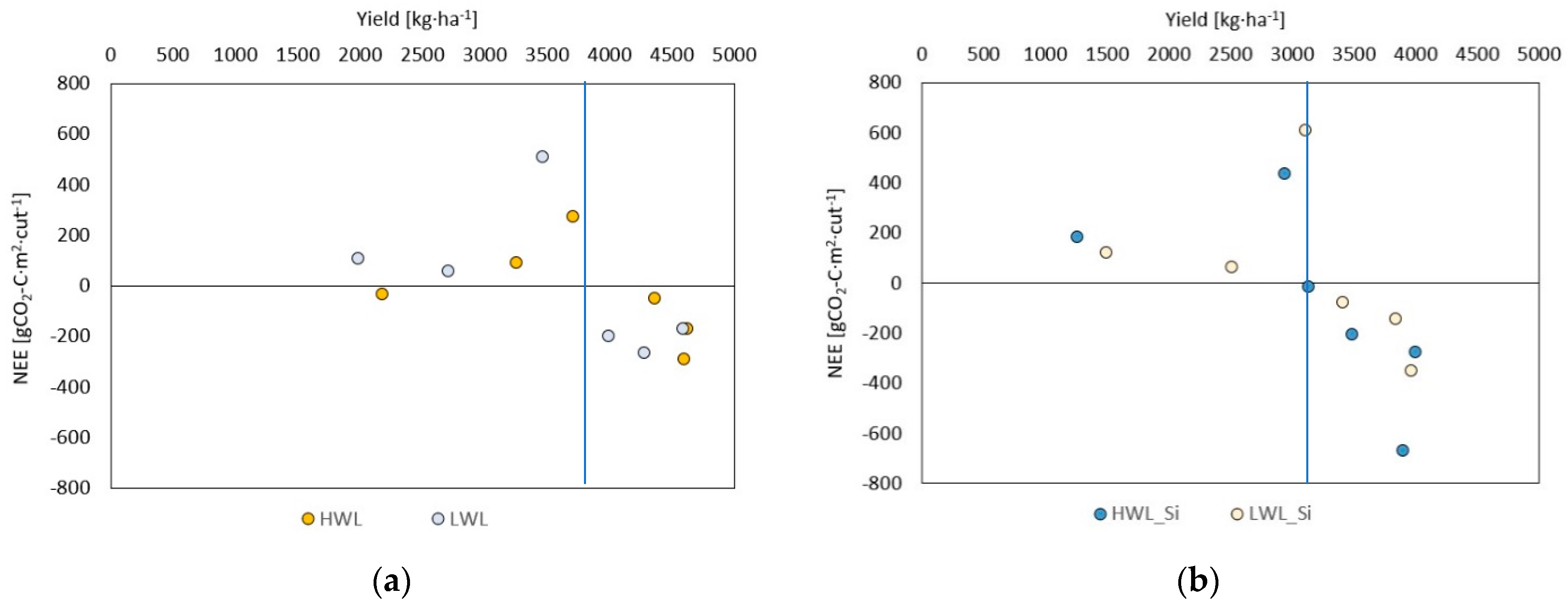
- The yield at which the meadow turns from being a net source to a net sink of CO2 in single cuts shifts from around 3800 kg·ha−1 at plots without silicon antitranspirant application to 3200 kg·ha−1 at treated plots, while cumulated NEE is more negative at the same biomass yield for plots with silicon application. It indicates that silicon antitranspirant application may have a positive effect on improving the carbon balance of meadows (either by reducing net emissions or increasing net assimilations).
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gilbert, N. One-third of our greenhouse gas emissions come from agriculture. Nature 2012, 31, 10–12. [Google Scholar] [CrossRef]
- Smith, P.; Clark, H.; Dong, H.; Elsiddig, E.A.; Haberl, H.; Harper, R.; House, J.; Jafari, M.; Masera, O.; Mbow, C.; et al. Agriculture, Forestry and Other Land Use (AFOLU). In Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O.R., Pichs-Madruga, Y., Sokona, E., Farahani, S., Kadner, K., Seyboth, A., Adler, I., Baum, S., Brunner, P., Eickemeier, B., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; Available online: https://www.ipcc.ch/pdf/assessment-report/ar5/wg3/ipcc_wg3_ar5_chapter11.pdf (accessed on 26 July 2023).
- Zhou, W.; Li, J.; Yue, T. Remote Sensing Monitoring and Evaluation of Degraded Grassland in China. Accounting of Grassland Carbon Source and Carbon Sink; Springer Geography; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Budzyńska, M.; Dąbrowska-Zielińska, K.; Tomaszewska, M.; Bartold, M.; Gatkowska, M. Badania nad obiegiem węgla na obszarach łąkowych [Studies on carbon exchange in grassland areas]. Łąkarstwo w Polsce 2015, 18, 47–58. [Google Scholar]
- Tiemeyer, B.; Freibauer, A.; Borraz, E.A.; Augustin, J.; Bechtold, M.; Beetz, S.; Beyer, C.; Ebli, M.; Eickenscheidt, T.; Fiedler, S.; et al. A new methodology for organic soils in national greenhouse gas inventories: Data synthesis, derivation and application. Ecol. Indic. 2020, 109, 105838. [Google Scholar] [CrossRef]
- Tiemeyer, B.; Frings, J.; Kahle, P.; Köhne, S.; Lennartz, B. A comprehensive study of nutrient losses, soil properties and groundwater concentrations in a degraded peatland used as an intensive meadow–implications for re-wetting. J. Hydrol. 2007, 345, 80–101. [Google Scholar] [CrossRef]
- Oleszczuk, R.; Regina, K.; Szajdak, L.; Maryganova, V. Impacts of Agricultural Utilization of Peat—Soil on the Greenhouse Gas Balance. In Peatlands and Climate Change; Strack, M., Ed.; International Peat Society, Saarijärven Offset Oy: Saarijärvi, Finland, 2008; pp. 70–97. [Google Scholar]
- Erkens, G.; van der Meulen, M.J.; Middelkoop, H. Double trouble: Subsidence and CO2 respiration due to 1,000 years of Dutch coastal peatlands cultivation. Hydrogeol. J. 2016, 24, 551–568. [Google Scholar] [CrossRef]
- Tanner, C.C.; Nguyen, M.L.; Sukias, J.P.S. Nutrient removal by a constructed wetland treating subsurface drainage from grazed dairy pasture. Agric. Ecosyst. Environ. 2005, 105, 145–162. [Google Scholar] [CrossRef]
- Łabędzki, L.; Kaca, E.; Brandyk, A. Irrigation and Drainage in Polish Agriculture: State, Problems and Needs. In Quality of Water Resources in Poland; Springer Water; Springer: Cham, Switzerland, 2021; pp. 91–118. [Google Scholar] [CrossRef]
- Sojka, M.; Kozłowski, M.; Stasik, R.; Napierała, M.; Kęsicka, B.; Jaskuła, J.; Liberacki, D.; Bykowski, J.; Wróżyński, R. Sustainable Water Management in Agriculture-The Impact of Drainage Water Management on Groundwater Table Dynamics and Subsurface Outflow. Sustainability 2019, 11, 4201. [Google Scholar] [CrossRef]
- Li, J.; Fei, L.; Li, S.; Shi, Z.; Liu, L. The influence of optimized allocation of agricultural water and soil resources on irrigation and drainage in the Jingdian Irrigation District, China. Irrig. Sci. 2020, 38, 1–11. [Google Scholar] [CrossRef]
- Meier, M.; Gerlach, R.; Schirmel, J.; Buhk, C. Plant diversity in a water-meadow landscape: The role of irrigation ditches. Plant Ecology 2017, 218, 971–981. [Google Scholar] [CrossRef]
- Beetz, S.; Liebersbach, H.; Glatzel, S.; Jurasinski, G.; Buczko, U.; Höper, H. Effects of land use intensity on the full greenhouse gas balance in an Atlantic peat bog. Biogeosciences 2013, 10, 1067–1082. [Google Scholar] [CrossRef]
- Leiber-Sauheitl, K.; Fuß, R.; Voigt, C.; Freibauer, A. High greenhouse gas fluxesfrom grassland on histic gleysol along soil C and drainage gradients. Biogeosci. Discuss. 2013, 10, 11283–11317. [Google Scholar] [CrossRef]
- Conant, R.T.; Paustian, K.; Elliott, E.T. Grassland management and conversion into grassland: Effects on soil carbon. Ecol. Appl. 2001, 11, 343–355. [Google Scholar] [CrossRef]
- Olsson, A.; Campana, P.E.; Lind, M.; Yan, J. Potential for carbon sequestration and mitigation of climate change by irrigation of grasslands. Appl. Energy 2014, 136, 1145–1154. [Google Scholar] [CrossRef]
- Schrier-Uijl, A.P.; Kroon, P.S.; Hendriks, D.M.D.; Hensen, A.; Van Huissteden, J.; Berendse, F.; Veenendaal, E.M. Agricultural peatlands: Towards a greenhouse gas sink—A synthesis of a Dutch landscape study. Biogeosciences 2014, 11, 4559–4576. [Google Scholar] [CrossRef]
- Jurczuk, S. Emisja dwutlenku węgla ze zmeliorowanych gleb organicznych w Polsce [Carbon dioxide emission from reclaimed organic soils in Poland]. Woda-Sr.-Obsz. Wiej. 2012, 12, 63–76. [Google Scholar]
- Buhk, C.; Schirmel, J.; Rebekka, G.; Frör, O. Traditional water meadows—A sustainable management type for the future? In Irrigation in Agroecosystems; Ondrasek, G., Ed.; InTechOpen: London, UK, 2018; ISBN 978-953-51-6428-9. [Google Scholar] [CrossRef]
- Chimner, R.A.; Cooper, D.J.; Bidwell, M.D.; Culpepper, A.; Zillich, K.; Nydick, K. A new method for restoring ditches in peatlands: Ditch filling with fiber bales. Restor. Ecol. 2019, 27, 63–69. [Google Scholar] [CrossRef]
- Schimelpfenig, D.W.; Cooper, D.J.; Chimner, R.A. Effectiveness of ditch blockage for restoring hydrologic and soil processes in mountain peatlands. Restor. Ecol. 2014, 22, 257–265. [Google Scholar] [CrossRef]
- Vasander, H.; Tuittila, E.-S.; Lode, E.; Lundin, L.; Ilomets, M.; Sallantaus, T.; Heikkilä, R.; Pitkänen, A.; Laine, J. Status and restoration of peatlands in northern Europe. Wetl. Ecol. Manag. 2003, 11, 51–63. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability; Working Group II contribution to the Sixth Assessment Report of the Intergovernamental Panel on Climate Change; IPCC: Cambridge, UK; New York, NY, USA, 2022; ISBN 9781009325844. [CrossRef]
- Kocięcka, J.; Liberacki, D. The Potential of Using Chitosan on Cereal Crops in the Face of Climate Change. Plants 2021, 10, 1160. [Google Scholar] [CrossRef]
- Kocięcka, J.; Liberacki, D.; Stróżecki, M. The Role of Antitranspirants in Mitigating Drought Stress in Plants of the Grass Family (Poaceae)—A Review. Sustainability 2023, 15, 9165. [Google Scholar] [CrossRef]
- Mphande, W.; Kettlewell, P.S.; Grove, I.G.; Farrell, A.D. The potential of antitranspirants in drought management of arable crops: A review. Agric. Water Manag. 2020, 236, 106143. [Google Scholar] [CrossRef]
- Kettlewell, P.S.; Heath, W.L.; Haigh, I.M. Yield enhancement of droughted wheat by film antitranspirant application: Rationale and evidence. Agric. Sci. 2010, 1, 143–147. [Google Scholar] [CrossRef]
- Gerhards, M.; Schlerf, M.; Rascher, U.; Udelhoven, T.; Juszczak, R.; Alberti, G.; Miglietta, F.; Inoue, Y. Analysis of airborne optical and thermal hyperspectral imagery for detection of water stress symptoms. Remote Sens. 2018, 10, 1139. [Google Scholar] [CrossRef]
- Kocięcka, J.; Liberacki, D.; Kupiec, J.M.; Stróżecki, M.; Dłużewski, P. Effects of Silicon Application and Groundwater Level in a Subirrigation System on Yield of a Three-Cut Meadow. Water 2023, 15, 2103. [Google Scholar] [CrossRef]
- Chemirol. Available online: https://chemirol.com.pl/pl_PL/product/krzemian (accessed on 28 July 2023).
- Huculak, W.; Makowiec, M. Wyznaczenie meteorologicznego okresu wegetacyjnego na podstawie jednorocznych materiałów obserwacyjnych. Zesz. Nauk. SGGW 1977, 25, 65–72. [Google Scholar]
- Kozuchowski, K.; Degirmendzic, J. Contemporary changes of climate in Poland: Trends and variation in thermal and solarconditions related to plant vegetation. Pol. J. Ecol. 2005, 53, 283–297. [Google Scholar]
- Juszczak, R.; Acosta, M.; Olejnik, J. Comparison of daytime and nighttime ecosystem respiration measured by the closed chamber technique on a temperate mire in Poland. Polish J. Environ. Stud. 2012, 21, 643–658. [Google Scholar]
- Juszczak, R.; Humphreys, E.; Acosta, M.; Michalak-Galczewska, M.; Kayzer, D.; Olejnik, J. Ecosystem respiration in a heterogeneous temperate peatland and its sensitivity to peat temperature and water table depth. Plant Soil 2013, 366, 505–520. [Google Scholar] [CrossRef]
- Juszczak, R.; Uździcka, B.; Stróżecki, M.; Sakowska, K. Improving remote estimation of winter crops gross ecosystem production by inclusion of leaf area index in a spectral model. PeerJ 2018, 6, e5613. [Google Scholar] [CrossRef]
- Acosta, M.; Juszczak, R.; Chojnicki, B.; Pavelka, M.; Havránková, K.; Leśny, J.; Foltýnová, L.; Urbaniak, M.; Machacova, K.; Olejnik, J. CO2 Fluxes from Different Vegetation Communities on a Peatland Ecosystem. Wetlands 2017, 37, 423–435. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurements fordensity effects due to heat and water vapor transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Kutzbach, L.; Schneider, J.; Sachs, T.; Giebels, M.; Nykänen, H.; Shurpali, N.J.; Martikainen, P.J.; Alm, J.; Wilmking, M. CO2 flux determination by closed-chamber methods can be seriously biased by inappropriate application of linearregression. Biogeosciences 2007, 4, 1005–1025. [Google Scholar] [CrossRef]
- Drösler, M. Trace Gas Exchange and Climatic Relevance of Bog Ecosystem, Southern Germany. Ph.D. Dissertation, Lehrstuhl für Vegetationsokologie, Department für Ökologie, Technischen Universität München, München, Germany, 2005. [Google Scholar]
- Hoffmann, M.; Jurisch, N.; Albiac Borraz, E.; Hagemann, U.; Drösler, M.; Sommer, M.; Augustin, J. Automated modeling of ecosystem CO2 fluxes based on periodic closed chamber measurements: A standardized conceptual and practical approach. Agric. For. Meteorol. 2015, 200, 30–45. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the Temperature Dependence of Soil Respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Ma, L.; Zhu, G.; Chen, B.; Zhang, K.; Niu, S.; Wang, J.; Ciais, P.; Zuo, H. A globally robust relationship between water table decline, subsidence rate, and carbon release from peatlands. Commun. Earth Environ. 2022, 3, 254. [Google Scholar] [CrossRef]
- Abdalla, M.; Feigenwinter, I.; Richards, M.; Vetter, S.H.; Wohlfahrt, G.; Skiba, U.; Pintér, K.; Nagy, Z.; Hejduk, S.; Buchmann, N.; et al. Evaluation of the ECOSSE Model for Estimating Soil Respiration from Eight European Permanent Grassland Sites. Agronomy 2023, 13, 1734. [Google Scholar] [CrossRef]
- Wang, C.; Fu, B.; Zhang, L.; Xu, Z. Soil moisture–plant interactions: An ecohydrological review. J. Soils Sediments 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Rodríguez-Iturbe, I.; Porporato, A. Ecohydrology of Water-Controlled Ecosystems: Soil Moisture and Plant Dynamics; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Kou, X.; Han, W.; Kang, J. Responses of Root System Architecture to Water Stress at Multiple Levels: A Meta-Analysis of Trials under Controlled Conditions. Front. Plant Sci. 2022, 13, 5157. [Google Scholar] [CrossRef]
- Wang, N.; Xia, L.; Goodale, C.L.; Butterbach-Bahl, K.; Kiese, R. Climate change can accelerate depletion of montane grassland C stocks. Glob. Biogeochem. Cycles 2021, 35, e2020GB006792. [Google Scholar] [CrossRef]
- de Vries, F.T.; Bloem, J.; Quirk, H.; Stevens, C.J.; Bol, R.; Bardgett, R.D. Extensive management promotes plant and microbial nitrogen retention in temperate grassland. PLoS ONE 2012, 7, e51201. [Google Scholar] [CrossRef]
- Poyda, A.; Reinsch, T.; Struck, I.J.; Skinner, R.H.; Kluß, C.; Taube, F. Low assimilate partitioning to root biomass is associated with carbon losses at an intensively managed temperate grassland. Plant Soil 2021, 460, 31–50. [Google Scholar] [CrossRef]
- Poyda, A.; Reinsch, T.; Kluß, C.; Loges, R.; Taube, F. Greenhouse gas emissions from fen soils used for forage production in northern Germany. Biogeosciences 2016, 13, 5221–5244. [Google Scholar] [CrossRef]
- Eickenscheidt, T.; Heinichen, J.; Drösler, M. The greenhouse gas balance of a drained fen peatland is mainly controlled by land-use rather than soil organic carbon content. Biogeosciences 2015, 12, 5161–5184. [Google Scholar] [CrossRef]
- Renou-Wilson, F.; Müller, C.; Moser, G.; Wilson, D. To graze or not to graze? Four years greenhouse gas balances and vegetation composition from a drained and a rewetted organic soil under grassland. Agr. Ecosyst. Environ. 2016, 222, 156–170. [Google Scholar] [CrossRef]
- Schmitt, M.; Bahn, M.; Wohlfahrt, G.; Tappeiner, U.; Cernusca, A. Land use affects the net ecosystem CO2 exchange and its components in mountain grasslands. Biogeosciences 2010, 7, 2297–2309. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Hammerle, A.; Haslwanter, A.; Bahn, M.; Tappeiner, U.; Cernusca, A. Seasonal and inter-annual variability of the net ecosystem CO2 exchange of a temperate mountain grassland: Effects of weather and management. J. Geophys. Res. 2008, 113, D08110. [Google Scholar] [CrossRef]
- Turbiak, J.; Ćwiklińska, P.; Duda, F. Carbon dioxide emission from raised bog surface after peat extraction. J. Water Land Dev. 2017, 35, 237. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Hastings, M.; Bell, J.; Smith, J.U.; Richards, M.; Nilsson, M.B.; Peichl, M.; Löfvenius, M.O.; Lund, M.; Helfter, C.; et al. Simulation of CO2 and attribution analysis at six European peatland sites using the ECOSSE model. Water Air Soil Pollut. 2014, 225, 2182. [Google Scholar] [CrossRef]
- Hardie, S.M.L.; Garnett, M.H.; Fallick, A.E.; Ostle, N.J.; Rowland, A.P. Bomb 14C analysis of ecosystem respiration reveals that peatland vegetation facilitates release of old carbon. Geoderma 2009, 153, 393–401. [Google Scholar] [CrossRef]
- Weideveld, S.T.J.; Liu, W.; van den Berg, M.; Lamers, L.P.M.; Fritz, C. Conventional subsoil irrigation techniques do not lower carbon emissions from drained peat meadows. Biogeosciences 2021, 18, 3881–3902. [Google Scholar] [CrossRef]
- Boonman, J.; Hefting, M.M.; van Huissteden, C.J.A.; van den Berg, M.; van Huissteden, J.; Erkens, G.; Melman, R.; van der Velde, Y. Cutting peatland CO2 emissions with water management practices. Biogeosciences 2022, 19, 5707–5727. [Google Scholar] [CrossRef]
- Radkowski, A.; Radkowska, I. Effects of Silicate Fertilizer on Seed Yield in Timothy-Grass (Phleum pratense L.). Ecol. Chem. Eng. 2018, 25, 169–180. [Google Scholar] [CrossRef]
- Aires, L.M.I.; Pio, C.A.; Pereira, J.S. Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years. Glob. Chang. Biol. 2008, 14, 539–555. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.; Xu, M.; Zhu, J.; Chen, N.; Jiang, Y.; Huang, K.; Zu, J.; Liu, Y.; Yu, G. Water availability is more important than temperature in driving the carbon fluxes of an alpine meadow on the Tibetan Plateau. Agric. For. Meteorol. 2018, 256, 22–31. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Dates | Duration (Days) | Average Daily Temperature (°C) | Precipitation (mm) | Average Soil Moisture HWL (%) | Average Soil Moisture LWL (%) | Average WTD at HWL (mbgl) | Average WTD at LWL (mbgl) |
|---|---|---|---|---|---|---|---|---|
| I cut 2021 | 24.03.21–31.05.21 | 69 | 9.0 | 98.2 | 56 | 50 | 0.34 | 0.61 |
| II cut 2021 | 1.06.21–14.07.21 | 44 | 19.4 | 95.7 | 47 | 42 | 0.66 | 0.81 |
| III cut 2021 | 15.07.21–30.09.21 | 78 | 16.8 | 138.5 | 41 | 33 | 0.79 | 0.98 |
| Growing season 2021 | 24.03.21–22.11.21 | 244 | 13.3 | 404.4 | 47 | 41 | 0.64 | 0.85 |
| I cut 2022 | 14.03.22–6.06.22 | 85 | 10.2 | 91.3 | 37 | 32 | 0.61 | 0.82 |
| II cut 2022 | 7.06.22–15.08.22 | 70 | 19.6 | 129.7 | 18 | 16 | 0.88 | 1.05 |
| III cut 2022 | 16.08.22–17.11.22 | 94 | 12.9 | 237.4 | 47 | 42 | 0.52 | 0.75 |
| Growing season 2022 | 14.03.22–15.11.22 | 247 | 13.9 | 458.4 | 35 | 31 | 0.65 | 0.86 |
| Growing Season (gCO2-C·m−2·Growing Season−1) | Annual (gCO2-C·m−2·Year−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Plot | HWL | HWL_Si | LWL | LWL_Si | HWL | HWL_Si | LWL | LWL_Si |
| 2021 | Reco | 2310.57 | 2327.85 | 2175.86 | 2225.61 | 2445.37 | 2446.85 | 2291.03 | 2353.52 |
| GPP | −1950.27 | −2245.61 | −1944.28 | −1743.83 | −2131.12 | −2397.30 | −2106.13 | −1912.36 | |
| NEE | 360.31 | 82.24 | 231.58 | 481.77 | 314.25 | 49.55 | 184.89 | 441.17 | |
| 2022 | Reco | 2092.23 | 1996.66 | 1723.40 | 1810.22 | 2234.95 | 2129.56 | 1847.76 | 1924.61 |
| GPP | −2321.79 | −2508.84 | −1829.08 | −1976.68 | −2389.14 | −2565.75 | −1888.21 | −2043.31 | |
| NEE | −229.55 | −512.18 | −105.69 | −166.46 | −154.19 | −436.19 | −40.45 | −118.70 | |
| 2021 | ||||
|---|---|---|---|---|
| 1st Cut | 2nd Cut | 3rd Cut | Year | |
| HWL | 4365.39 | 4621.57 | 3706.08 | 12,693.04 |
| HWL_Si | 3489.42 (−20.1%) | 3999.88 (−13.5%) | 2939.83 (−20.7%) | 10,429.13 (−17.8%) |
| LWL | 4587.14 | 3993.01 | 3467.47 | 12,047.61 |
| LWL_Si | 3842.25 (−16.2%) | 3410.8 (−14.6%) | 3111.19 (−10.3%) | 10,364.24 (−14.0%) |
| 2022 | ||||
| HWL | 4598.28 | 3253.43 | 2179.26 | 10,030.97 |
| HWL_Si | 3897.24 (−15.2%) | 3135.94 (−3.6%) | 1260.74 (−42.1%) | 8293.92 (−17.3%) |
| LWL | 4283.50 | 2710.34 | 1984.71 | 8978.55 |
| LWL_Si | 3966.99 (−7.4%) | 2513.96 (−7.2%) | 1497.3 (−24.6%) | 7978.25 (−11.1%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocięcka, J.; Stróżecki, M.; Juszczak, R.; Liberacki, D. Effect of Subirrigation and Silicon Antitranspirant Application on Biomass Yield and Carbon Dioxide Balance of a Three-Cut Meadow. Water 2023, 15, 3057. https://doi.org/10.3390/w15173057
Kocięcka J, Stróżecki M, Juszczak R, Liberacki D. Effect of Subirrigation and Silicon Antitranspirant Application on Biomass Yield and Carbon Dioxide Balance of a Three-Cut Meadow. Water. 2023; 15(17):3057. https://doi.org/10.3390/w15173057
Chicago/Turabian StyleKocięcka, Joanna, Marcin Stróżecki, Radosław Juszczak, and Daniel Liberacki. 2023. "Effect of Subirrigation and Silicon Antitranspirant Application on Biomass Yield and Carbon Dioxide Balance of a Three-Cut Meadow" Water 15, no. 17: 3057. https://doi.org/10.3390/w15173057