
First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea


Abstract
:1. Introduction
2. Materials and Methods
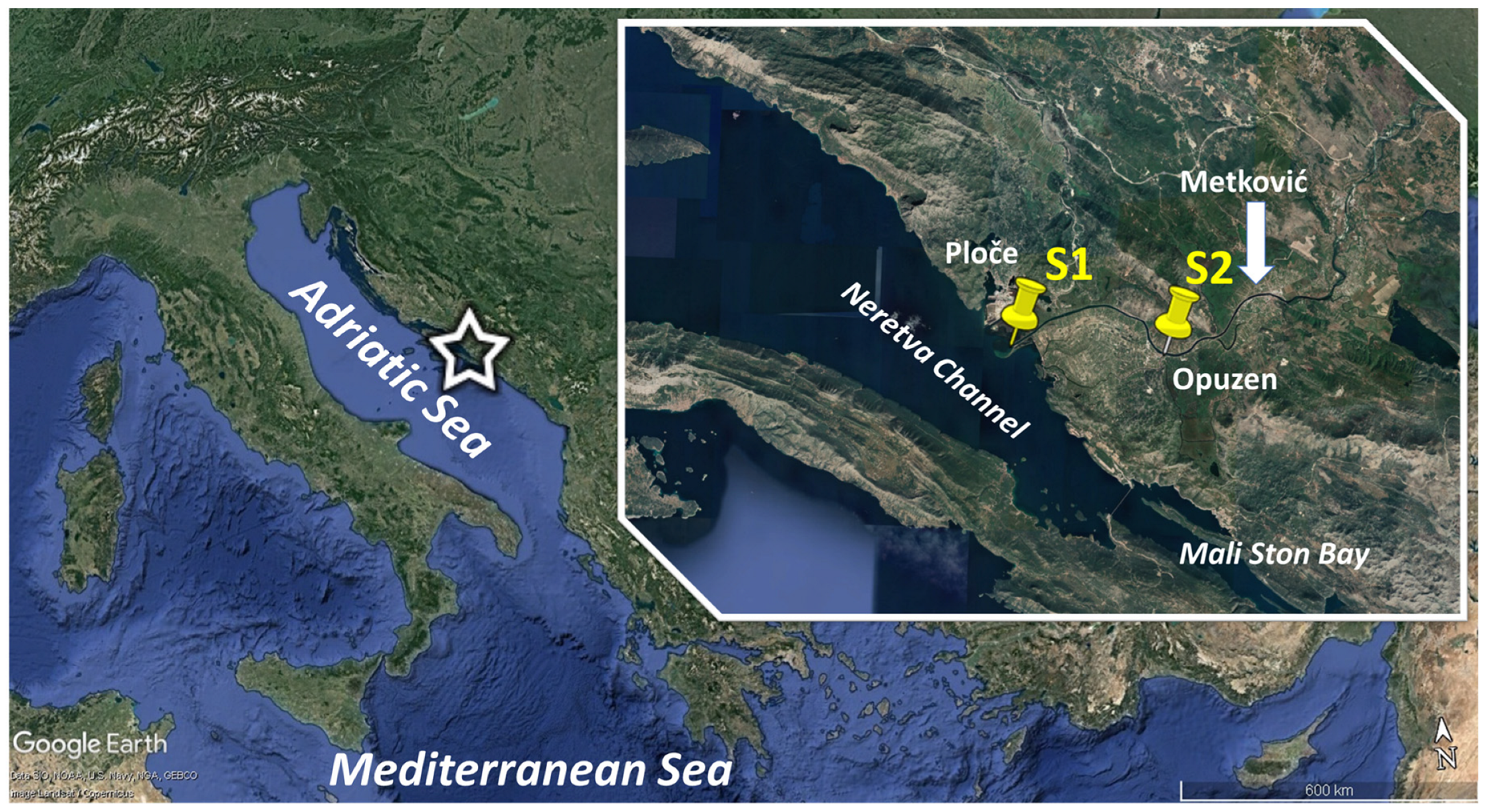
2.1. Study Area
2.2. Field Sampling and Laboratory Processing
2.3. Data Analysis
3. Results
3.1. Morphological Features of Rhizodomus tagatzi
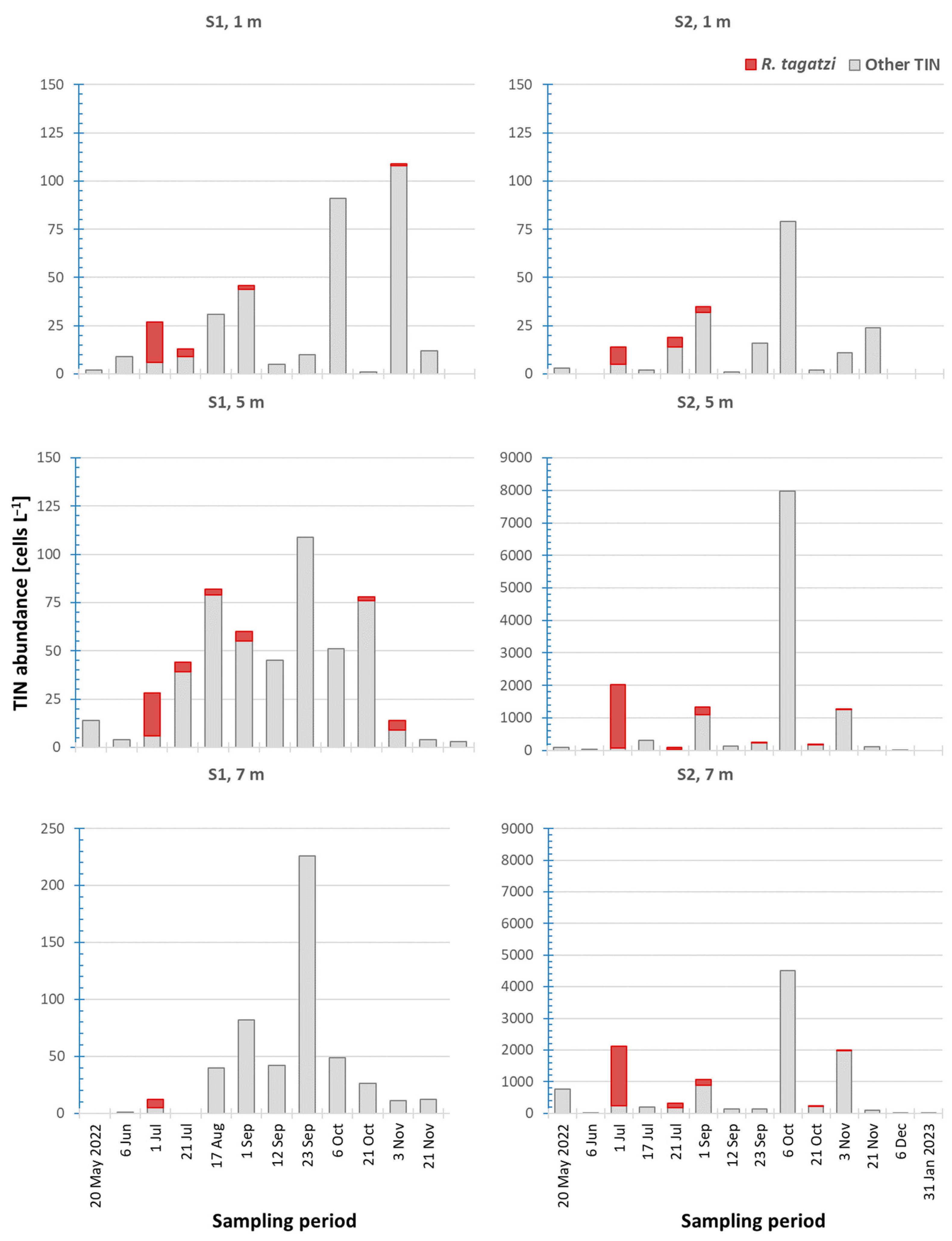
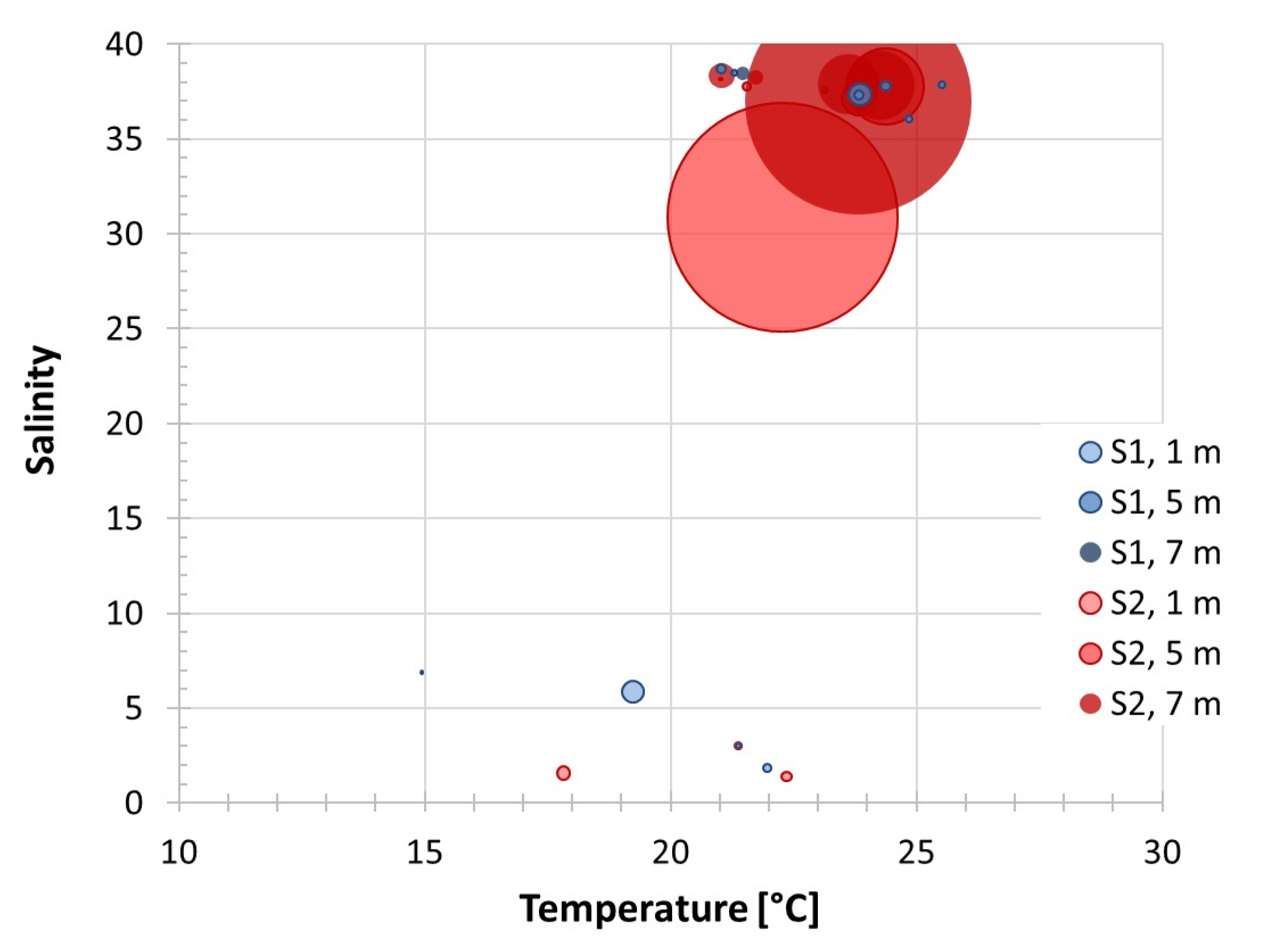
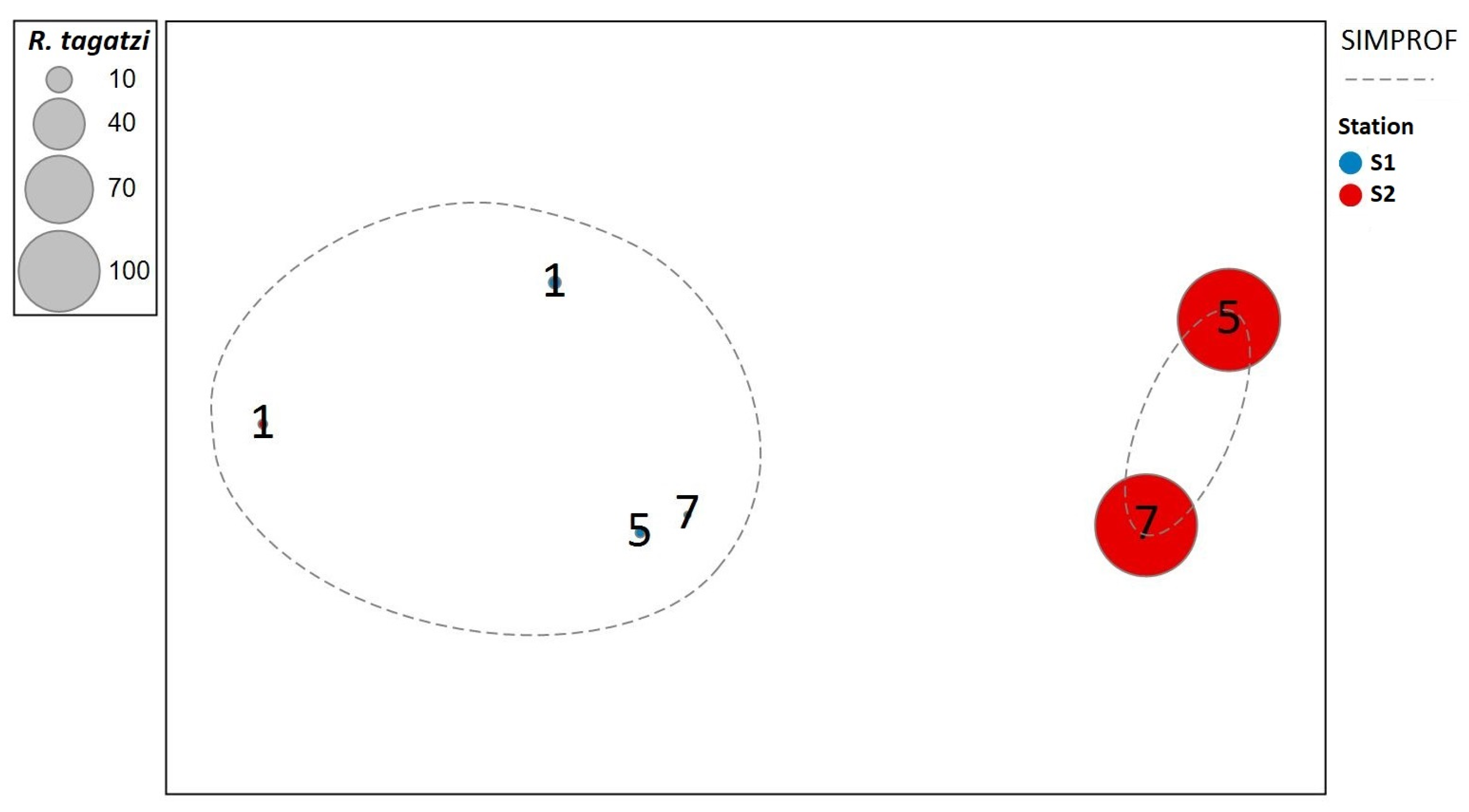
3.2. Distribution Pattern of Rhizodomus tagatzi and Environmental Settings in the Neretva River Estuary
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Tem./Sal. | Abundance/Biomass | Reference |
|---|---|---|---|
| Thau Lagoon, northern Mediterranean Sea | Warmer waters with high salinity 11.8–23.2 °C/31.5–37.9 | June 1994 25,215 µg m−3 | [12] 2 |
| Lake Faro, NE corner of Sicily, Italy (central Mediterranean Sea) | 12.5–29.3 °C/34.1–37.0 19.0 °C and 36.4 | July–September 2022 4.52 × 103 cells L−1 (July 2002) | [4,13] 1,2 |
| Damietta Harbour, Egypt (Eastern Mediterranean Sea) | 27–33 °C | July 2003 Max. total TIN abundance 73.5 × 103 cells m−3 (no data for T. corniger) | [14] 2 |
| Northern Lebanese coastal waters (Eastern Mediterranean Sea) | No data | July–October September 2006 and 2014 (7 cells L−1) August 2010 | [17] 3 |
| Novorossiysk Harbour and Gelendzhik Bay, North-eastern Black Sea | 26–28 °C | 5 × 104 cells m−3 (August-September 2015) | [49] 1 |
| Sevastopol Bay, Black Sea | Summer: 18.2–28.0 °C (23.6 ± 1.7 °C)/15.5–18.0 (17.6 ± 0.2) Autumn: 5.0–20.9 °C (14.1 ± 3.0 °C)/17.2–17.88 (17.63 ± 0.11) | summer–autumn (summer 2010: 45% of total TIN abundance) Mean 1: 12,819 cells m−3 1.01 × 105 cells m−3 (summer) | [18] 1 |
| Marmara Sea, Gulf of Gamlik, Turkish coastal waters | 18–28 °C/16–18 18 °C/16 19 °C and 18 | August–October 2010 850 cells L−1 (October) | [16] 2 |
| Al-Max Bay, Alexandria, Egypt | ~22 °C/~24 | Autumn 2014 16,902 ± 6453 cells m−3 | [50] 3 |
| Neretva River estuary, Adriatic Sea | 21.01–25.51 °C/30.88–38.71 22.28 °C and 30.88 | July–November 2022 1958 cells L−1 (1 July 2022) | This paper 1 |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agatha, S.; Strüder-Kypke, M.C. Phylogeny of the order Choreotrichida (Ciliophora, Spirotricha, Oligotrichea) as inferred from morphology, ultrastructure, ontogenesis, and SSr-RNA gene sequences. Eur. J. Protistol. 2007, 43, 37–63. [Google Scholar] [CrossRef] [PubMed]
- Strüder-Kypke, M.; Lynn, D.H. Morphological versus molecular data-phylogeny of tintinnid ciliates (Ciliophora, Choreotrichia) inferred from small subunit rRNA gene sequences. Denisia 2008, 23, 417–424. [Google Scholar]
- Bachy, C.; Gómez, F.; López-García, I.; Dolan, J.R.; Moreira, D. Molecular phylogeny of tintinnid ciliates (Tintinnida, Ciliophora). Protist 2012, 163, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Saccà, A.; Giuffrè, G. Biogeography and ecology of Rhizodomus tagatzi, a presumptive invasive tintinnid ciliate. J. Plankton Res. 2013, 35, 894–906. [Google Scholar] [CrossRef]
- Dolan, J.R. Planktonic protists: Little bugs pose big problems for biodiversity assessment. J. Plankton Res. 2016, 38, 1044–1051. [Google Scholar] [CrossRef]
- Strelkow, A.A.; Wirketis, M.A. New planktonic infusorian (suborder Tintinnoinea) from Peter the Great Bay. Rep. USSR Acad. Sci. 1950, 74, 389–391. (In Russian) [Google Scholar]
- Hada, Y. New species of the Tintinnida found from Inland Sea. Bull. Suzugamine Women Coll. Nat. Sci. 1964, 11, 1–4. [Google Scholar]
- Balech, E. Tintinnoinea del Mediterraneo; Trabajo Numero 28; Instituto Espanol de Oceanographia: Madrid, Spain, 1968; p. 84. [Google Scholar]
- Aladro-Lubel, M.A. Distribucion de los Tintinidos (Protozoa, Ciliata) de la parte oester suroeste y sur del Golfo de Mexico. Rev. De Las Soc. Mex. De Hist. Nat. 1974, 35, 45–75. [Google Scholar]
- Sharaf, G.M. On some tintinnids (Protozoa: Ciliata) from the Strait of Hormoz and the United Arab Emirates waters. Qatar Univ. Sci. J. 1995, 15, 389–398. [Google Scholar]
- Paulmier, G. Tintinnides (Ciliophora, Oligotrichida, Tintinnina) de l’Atlantique Boréal, de l’Océan Indien et de Quelques Mers Adjacentes: Méditerranée, Mer Caraibe, Mer Rouge. Inventaire et distribution. Observations Basées sur les Loricas; Rapport IFRAMER DRV/RH-ECOHAL: L’Houmeau, France, 1997; p. 127. Available online: https://archimer.ifremer.fr/doc/00424/53609/ (accessed on 22 March 2023).
- Lam-Hoai, T.; Rougoer, C.; Lasserre, G. Tintinnids and rotifers in a northern Mediterranean coastal lagoon. Structural diversity and function through biomass estimation. Mar. Ecol. Prog. Ser. 1997, 152, 13–25. [Google Scholar] [CrossRef]
- Saccà, A.; Guglielmo, L.; Bruni, V. Vertical and temporal microbial community patterns in a meromictic coastal lake influenced by the Straits of Massina upwelling system. Hydrobiologia 2008, 600, 89–104. [Google Scholar] [CrossRef]
- Dorgham, M.M.; Abdel-Aziz, E.N.; El-Ghobashy, E.A.; El-Tohamy, S.W. Preliminary study on protozoan community in Damietta Harbor, Egypt. Glob. Vet. 2009, 3, 495–502. [Google Scholar]
- Yurga, L. Non-native two new Tintinnida (Oligotrichida) taxa in Izmir Bay: Leprotintinnus nordquisti (Brandt 1906) ve Tintinnopsis corniger (Hada, 1964). Ege J. Fish. Aqua. Sci. 2012, 29, 69–72. [Google Scholar] [CrossRef]
- Durmuş, T.; Balci, M.; Balkis, N. Species of genus Tintinnopsis Stein, 1867 in Turkish coastal waters and new record of Tintinnopsis corniger Hada, 1964. Pakisten J. Zool. 2011, 44, 383–388. [Google Scholar]
- Abbou-Abi Saab, M. Tintinnids of the Lebanese Coastal Waters (Eastern Mediterranean); CNRS-Lebanon/UNEP/MAP/RAS/SPA Lebanon: Batroun, Lebanon, 2008; p. 192. [Google Scholar]
- Gavrilova, N.A.; Dovgal, I.V. Tintinnid Ciliates (Ciliophora, Tintinnida) of the Black Sea and the Sea of Azov; A.O. Kovalevsky Institute of Biology of the Southern Seas of Ras: Sevastopol, Crimea, 2019; p. 176. (In Russian) [Google Scholar]
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A.C. Building the European Alien Species Information Network (EASIN): A novel approach for the exploration of distributed alien species data. BioInvasions Rec. 2012, 1, 235–245. [Google Scholar] [CrossRef]
- European Commission-Joint Research Centre-European Alien Species Information Network (EASIN). Available online: https://easin.jrc.ec.europa.eu/ (accessed on 10 February 2023).
- Matić, F. Air-Sea Interaction in Split, Brač and Neretva Channel. Master’s Thesis, Faculty of Science, University of Zagreb, Zagreb, Croatia, 2005; p. 75. (In Croatian). [Google Scholar]
- Samokovlija Dragičević, J. Quality of water and improvement of the delta and drainage area of the Neretva River. Građevinar 2023, 55, 759–765. (In Croatian) [Google Scholar]
- Ljubenkov, I.; Vranješ, M. Numerical model of stratified flow-case study of the Neretva river bed salination (2004). Građevinar 2012, 64, 101–113. [Google Scholar]
- Krvavica, N.; Ružić, I. Assessment of sea-level rise impacts on salt-wedge intrusion in idealized and Neretva River Estuary. Estuar. Coast. Shelf Sci. 2020, 234, 106638. [Google Scholar] [CrossRef]
- Bojanić, N.; Šolić, M.; Krstulović, N.; Šestanović, S.; Marasović, I.; Ninčević, Ž. Temporal variability and abundance and biomass of ciliates and copepods in the eutrophicated part of Kaštela Bay (Middle Adriatic Sea). Helgol. Mar. Res. 2005, 59, 107–120. [Google Scholar] [CrossRef]
- Clark, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 1st ed.; PRIMER-e Ltd.: Plymouth, UK, 2015; pp. 1–300. [Google Scholar]
- Xu, K.; Agatha, S.; Dolan, J. World Ciliophora Database. Rhizodomus tagatzi Strelkow & Wirketis, 1950. Accessed through: World Register of Marine Species 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=689307 (accessed on 10 February 2023).
- Abou Zaid, M.M.; Hellal, A.M. Tiintinnids (Protozoa: Ciliata) from the coast of Hurghada Red Sea, Egypt. Egypt. J. Aquat. Res. 2012, 38, 249–268. [Google Scholar] [CrossRef]
- Abboud-Abi Saab, M.; Ouba, A. Seasonal and interannual changes in abundance and community structure of tintinnids in the Levantine Sea in relation to hydrographic conditions from 2002 to 2014. Reg. Stud. Mar. Sci. 2021, 47, 101990. [Google Scholar] [CrossRef]
- Gavrilova, N.A.; Dovgal, I.V. Tintinnid ciliates (Spirotrichea, Choreotrichia, Tintinnida) of the Black Sea: Recent invasions. Protistology 2016, 10, 91–96. [Google Scholar] [CrossRef]
- Dolan, J.R.; Pierce, R.W. Diversity and distribution of tintinnids. In The biology and ecology of tintinnid ciliates Models for marine plankton, 1st ed.; Dolan, J.R., Montagnes, D.J.S., Agatha, S., Coats, D.W., Stoecker, D.K., Eds.; Wiley-Blackwell A. John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 214–243. [Google Scholar]
- Vidjak, O.; Bojanić, N.; Kušpilić, G.; Gladan Ninčević, Ž.; Tičina, V. Zooplankton community and hidrographycal properties of the Neretva Channel (eastern Adriatic Sea). Helgol. Mar. Res. 2007, 61, 267–282. [Google Scholar] [CrossRef]
- Jasprica, N.; Hafner, D.; Batistić, M.; Kapetanović, T. Phytoplankton in three freshwater lakes in the Neretva River delta /Eastern Adriatic, NE Mediterranean). Nova Hedwig. 2005, 81, 37–54. [Google Scholar] [CrossRef]
- Jasprica, N.; Carić, M.; Kršinić, F.; Kapetanović, T.; Batistić, M.; Njire, J. Planktonic diatoms and their environment in the lower Neretva River estuary (Eastern Adriatic Sea, NE Mediterranean). Nova Hedwig. 2012, 141, 405–430. [Google Scholar]
- Bužančić, M.; Ninčević Glada, Ž.; Marasović, I.; Kušpilić, G.; Grbec, B. Eutrophication influence on phytoplankton community composition in three bays on the eastern Adriatic coast. Oceanologia 2016, 58, 302–316. [Google Scholar] [CrossRef]
- Lončar, G.; Krvavica, N.; Gotovac, H.; Oskoruš, D.; Kulić, T. Numerical analysis of the impact of the dam on the prevention of saltwater intrusion into the Neretva riverbed. Hrvat. Vode 2020, 28, 113–124. (In Croatian) [Google Scholar]
- Skejić, S.; Vilibić, I.; Matijević, S.; Jozić, S.; Ninčević Gladan, Ž.; Morović, M.; Marasović, I.; Prelesnik, H. Long-term regulating mehanisms of phytoplankton biomass in a tradicional shellfish aquaculture area. Fresenius Environ. Bull. 2015, 24, 3001–3013. [Google Scholar]
- Bojanić, N.; Šolić, M.; Krstulović, N.; Šestanović, S.; Ninčević Gladan, Ž.; Marasović, I.; Brautović, I. The role of ciliates within the microbial food web in the eutrophicated part of Kaštela Bay (middle Adriatic Sea). Sci. Mar. 2006, 70, 431–442. [Google Scholar] [CrossRef]
- Bojanić, N.; Vidjak, O.; Šolić, M.; Krstulović, N.; Brautović, I.; Matijević, S.; Kušpilić, G.; Šestanović, S.; Ninčević Gladan, Ž.; Marasović, I. Community structure and seasonal dynamics of tintinnid ciliates in Kaštela Bay (middle Adriatic Sea). J. Plankton Res. 2012, 34, 510–530. [Google Scholar] [CrossRef]
- Gavrilova, N.; Dolan, J.R. A note on species list and ecosystem shift: Black Sea Tintinnids, Ciliates of the microzooplankton. Acta Protozool. 2007, 46, 279–288. [Google Scholar]
- Šolić, M.; Krstulović, N.; Šantić, D.; Šestanović, S.; Ordulj, M.; Bojanić, N.; Kušpilić, G. Structure of microbial communities in phosphorus-limited estuaries along the eastern Adriatic coast. J. Mar. Biol. Assoc. United Kingd. 2015, 95, 1565–1578. [Google Scholar] [CrossRef]
- Kršinić, F. Tintinnids (Tintinnida, Choreotrichia, Ciliata) in the Adriatic Sea, Mediterranean Part II. Ecology; Institute of Oceanography and Fisheries: Split, Croatia, 2010; p. 116. [Google Scholar]
- Kršinić, F.; Precali, R. On the occurrence of oceanic tintinnines with a particular consideration of the species Amphorides laackmanni (Jörgensen, 1924), (Ciliophora, Oligotrichida, Tintinnina) in the northern Adriatic Sea. Mar. Ecol. 1997, 18, 67–81. [Google Scholar] [CrossRef]
- Lunven, M.; Guillaud, J.F.; Youénou, A.; Crassous, M.P.; Berric, R.; Le Gall, E.; Kérouel, R.; Labry, C.; Aminot, A. Nutrient and phytoplankton distribution in the Loire River plume (Bay of Biscay, France) resolved by a new Fine Scale Sampler. Estuar. Coast. Shelf Sci. 2005, 65, 94–108. [Google Scholar] [CrossRef]
- Cermeño, P.; Dutkiewicz, S.; Harris, R.P.; Follows, M.; Schofield, O.; Falkowski, G. The role of nutricline depth in regulation the ocean carbon cycle. Proc. Natl. Acad. Sci. USA 2009, 105, 20344–20349. [Google Scholar] [CrossRef]
- Richardson, K.; Bendtsen, J. Vertical distribution of phytoplankton and primary production in relation to nutricline depth in the open ocean. Mar. Ecol. Prog. Ser. 2019, 620, 34–46. [Google Scholar] [CrossRef]
- Tyler, M.A. Dye tracing of a subsurface chlorophyll maximum of a red-tide dinoflagellate to surface frontal regions. Mar. Biol. 1984, 78, 285–300. [Google Scholar] [CrossRef]
- Akizawa, K.; Iwasaki, N.; Ueda, H. List of microplankton in Uranouchi Inlet, Kochi, Japan during the summer of 1997. Bull. Mar. Sci. Fish. Kochi Univ. 1998, 18, 57–63. [Google Scholar]
- Selifonova, Z.P.; Makarevich, P.R. Invasive alien species of tintinnid ciliates from the northeaster Black Sea, Russian and Abkhazian coast. Protistology 2018, 12, 185–190. [Google Scholar] [CrossRef]
- El-Damhougy, K.A.; El-Sayed, A.A.; Aboul Ezz, S.M.; Abu Husein, M.S. Tintinnida (Ciliophora) as bio-indicator for certain pollutants at al-max area, Alexandria, Egypt. Int. J. Fish. Aquat. Stud. 2017, 5, 297–305. [Google Scholar]
- Feng, M.; Wang, C.; Zhang, W.; Zhang, G.; Xu, H.; Zhao, Y.; Xiao, T.; Wang, C.; Wang, W.; Bi, Y.; et al. Dataset of long-term variation in species occurrence and abundance of tintinnid assemblages in Jiaozhou Bay, China. Data Brief 2018, 19, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Saccà, A.; Strüder-Kypke, M.C.; Lynn, D.H. Redescription of Rhizodomus tagatzi (Ciliophora: Spirotrichea: Tintinnida), based on morphology and small subunit ribosomal RNA gene sequence. J. Eukaryot. Microbiol. 2012, 59, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Pierce, W.R.; Carlton, J.T.; Carlton, D.A.; Geller, J.B. Ballast water as a vector for tintinnid transport. Mar. Ecol. Prog. Ser. 1997, 149, 295–297. [Google Scholar] [CrossRef]
- Uttieri, M.; Aguzzi, L.; Aiese Cigliano, R.; Amato, A.; Bojanić, N.; Brunetta, M.; Camatti, E.; Carotenuto, Y.; Damjanović, T.; Delpy, F.; et al. WGEUROBUS–Working Group “Towards a EURopean OBservarory of the non-indigenous calanoid copepod Pseudodiaptomus marinUS”. Biol. Invasions 2020, 22, 885–906. [Google Scholar] [CrossRef]
- Spagnolo, A.; Auriemma, R.; Bacci, T.; Balković, I.; Bertasi, F.; Bolognini, L.; Cabrini, M.; Cilenti, L.; Cuicchi, C.; Cvitković, I.; et al. Non-indigenous macrozoobentic species on hard substrata of selected harbours in the Adriatic Sea. Mar. Pollut. Bull. 2019, 147, 150–158. [Google Scholar] [CrossRef]
- Lučić, D.; Onofri, I.; Garić, R.; Violić, I.; Vranješ, M.; Gangai Zovko, B.; Jurinović, J.; Njire, J.; Hure, M. Ingeression og the hydromedusa Neotima lucullana (delle chiaje, 1822) into the ecosystem of the Neretva River estuary (south-eastern Adriatic, Croatia). Acta Adriat. 2022, 63, 165–174. [Google Scholar] [CrossRef]
- Hasanspahić, N.; Pećarević, M.; Hrdalo, N.; Čampara, L. Analysos of ballast waer discharged in port–A case study of the port of Ploče (Croatia). J. Mar. Sci. Eng. 2022, 10, 1700. [Google Scholar] [CrossRef]
- Kamiyama, T. Comparative biology of tintinnid cysts. In The Biology and Ecology of Tintinnid Ciliates Models for Marine Plankton, 1st ed.; Dolan, J.R., Montagnes, D.J.S., Agatha, S., Coats, D.W., Stoecker, D.K., Eds.; Wiley-Blackwell A. John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 171–185. [Google Scholar]
- Ganser, M.H.; Bartel, H.; Weiβenbavher, B.; Andosch, A.; Lütz-Meindl, U.; Radacher, P.; Agatha, S. A light and electron microscopical study on resting cyst on the tintinnid Schmidingerella (Alveolata. Ciliophora) including a phylogeny-aware comparison. Eur. J. Protistol. 2022, 86, 125922. [Google Scholar] [CrossRef]
- Basoli, J. Fishing in the Adriatic. Special edition; Znanje: Zagreb, Croatia, 1984; p. 392. (In Croatian) [Google Scholar]
- Vidjak, O.; Bojanić, N.; de Olazabal, A.; Benzi, M.; Brautović, I.; Camatti, E.; Hure, M.; Lipej, L.; Lučić, D.; Pansera, M.; et al. Zooplankton in Adriatic port environments: Indigenous communities and non-indigenous species. Mar. Pollut. Bull. 2019, 147, 133–149. [Google Scholar] [CrossRef]
- Ninčević Gladan, Ž.; Vidjak, O.; Žuljević, A.; Šolić, M.; Cvitković, I.; Matić Skoko, S. Port Baseline Survey; (Final Report) BALMAS project. WP5. Activity 5.1.; Institute of Oceanography and Fisheries: Split, Croatia, 2016; pp. 1–31. [Google Scholar]






| Study Area | Total Lorica Length [µm] | OLD [µm] | Horn Length [µm] | Reference |
|---|---|---|---|---|
| Peter the Great Bay, Russia (subpolar sector of the Japan Sea) | 168.3–244.8 (213.65 ± 1.44) | 30.6–35.7 (30.6) | 40.8–91.8 (70.25 ± 0.93) | [6] 1 |
| Strait of Hormoz and the United Arab Emirates waters | 166–200 | 29–31 | >1/3 total length | [10] 2 |
| Urbino Lagoon (Corsica, Western Mediterranean Sea | 170–190 | 30–35 | No data | [11] 3 |
| Lebanese coastal waters (Eastern Mediterranean Sea) | 130–190 | 27–28 | 44–46 | [17] 3 |
| Marmara Sea, Gulf of Gamlik, Turkish coastal waters | 165–170 (167 ± 1.9) | 30–33 (31 ± 1.1) | 40–45 (43 ± 2) | [16] 2 |
| Lake Faro, NE corner of Sicily (central Mediterranean Sea) | 133–250 (178) | 29–35 (32) external | 35–60 (50) | [52] 1 |
| Izmir Bay, Turkish coastal water | 185 | 33 | Up to 40 | [15] 2 |
| Sevastopol Bay, Black Sea | 133–205 | 30–40 | 44–46 | [18] 1 |
| Adriatic Sea, Neretva River estuary | 149.1–217.6 (175.3 ± 19.7) | 28.7–36.1 (32.1 ± 1.6) | 34.3–56.5 (44.7 ± 6.0) | This paper 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njire, J.; Bojanić, N.; Lučić, D.; Violić, I. First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water 2023, 15, 1821. https://doi.org/10.3390/w15101821
Njire J, Bojanić N, Lučić D, Violić I. First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water. 2023; 15(10):1821. https://doi.org/10.3390/w15101821
Chicago/Turabian StyleNjire, Jakica, Natalia Bojanić, Davor Lučić, and Ivana Violić. 2023. "First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea" Water 15, no. 10: 1821. https://doi.org/10.3390/w15101821
APA StyleNjire, J., Bojanić, N., Lučić, D., & Violić, I. (2023). First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water, 15(10), 1821. https://doi.org/10.3390/w15101821