
Allometric Relationships between Dry Matter Weights of Maize Organs and Their Responses to Drought
by
 ,
,
Huiqing Ming
1,
Fu Cai
2,3,*,
Na Mi
2,3,
Yushu Zhang
2,3,
Hui Zhang
4,
Shujie Zhang
2,3,
Xianli Zhao
2 and
Yanrui Feng
2,5 1
Liaoning Province Meteorological Service Center, Shenyang 110166, China
2
Institute of Atmospheric Environment, China Meteorological Administration, Shenyang 110166, China
3
Key Laboratory of Agrometeorological Disasters, Shenyang 110166, China
4
Jinzhou Ecology and Agriculture Meteorological Center, Jinzhou 121001, China
5
Department of Meteorology, Shenyang Agricultural University, Shenyang 110161, China
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 99; https://doi.org/10.3390/w15010099
Submission received: 9 November 2022
/
Revised: 15 December 2022
/
Accepted: 24 December 2022
/
Published: 28 December 2022
(This article belongs to the Section Water, Agriculture and Aquaculture)
Abstract
:To understand the drought response mechanisms of dry matter partitioning of maize, pot experiments under drought conditions were conducted during the vegetative and reproductive growth periods of maize. The aim was to calculate allometric relationships between the dry matter weight of different organs and their responses to drought. Results showed that allometric relationships between the roots, above-ground plant, and total biomass gradually increased with maize growth approaching maturity under a normal water supply. Drought during the vegetative period reinforced allometric relationships during the growth process and after rewatering and increased the size-dependence of the root shoot ratio. However, drought during the reproductive period weakened them. The ear–shoot allometric relationship was more significant during growth than at later growth under normal conditions but strengthened during later growth in those plants suffering droughts during the vegetative and reproductive stages. The ear–shoot allometric relationship and the size-dependence of harvest index at later growth were significantly enhanced by drought during the reproductive period compared to the vegetative period.
1. Introduction
The photosynthate of plants is distributed into different organs and the allocation pattern is reflected by the adaptive strategy of plants to environmental change [1]. The dry matter partitioning (DMP) ratio of plant organs has traditionally been considered as reflecting the distribution status of biomass [2] and the DMP has been conceptualized as the process driven by the ratio [3,4]. Root shoot ratio (RSR), harvest index (HI), and reproductive effort are regarded as the vital parameters which reflect the DMP pattern of a plant and play a key role in assessing crop yield and building models [5]. In the above-mentioned ratio-based perspective, allocation patterns are size-independent, but other observations suggest otherwise [6]. Some scholars believe that DMP is allometric, regarded as a process in which the dry matter weight of different organs depends on the evolution of plant size and there is a functional relationship between growth and allocation, rather than a simple allocation ratio or proportion issue [7], and various proportion indices are also correlated with plant size [8]. This means that the change in the DMP pattern is the response of the allometric trajectory to the environment and the allometric relationship (AR) can be used to replace the allocation ratio to explain the effect of environmental or production mode change on DMP more clearly and effectively [1]. The identification of ARs between different organs such as leaf and stalk, leaf and root, vegetative and reproductive organs, and aboveground and underground biomass are important indicators for understanding the environmental response strategies of plants [9,10].
In terms of crops, biomass production and allocation are essential for yield formation and they are influenced by abiotic factors such as nutrition, temperature, light, and soil moisture [11,12]. Drought is one of the most important meteorological disasters affecting grain production [13]. As a consequence of drought, the reduction of photosynthetic products can be caused by the constrained photosynthetic process [14] and the growth of roots and leaves can be affected by the altered DMP pattern which, in turn, affects physiological processes such as nutrient absorption and photosynthesis [15]. In addition, the ripening effect of drought inhibits the redistribution of dry matter in the reproductive growth stage [16].
The current mainstream crop models do not explain the effects of drought conditions on DMP and they lack the ability to simulate the drought process [16]. Although relevant studies have been conducted [17], root biomass, or especially total root biomass is rarely considered, which severely limits the understanding of the drought response on DMP. The existing studies on DMP based on allometric theory have mostly focused on the effects of planting density [8], sowing date [10], tillage methods [18], and soil fertility [19], while those considering the effect of drought are still scarce. The application of allometric theory to the investigation of drought response of DMP can contribute to an in-depth understanding of the mechanisms involved, play a key role in refining plant disaster theory, and improve crop model parameter schemes [20]. As one of the world’s three major grains, maize has ranked first in planting area and yield in recent years and plays an increasingly key role in guaranteeing world food security and economic development [14]. Maize is also sensitive to water during its whole growth period, so drought has become the most important disaster affecting maize production [16], and it is vital to investigate the effect of drought on the DMP of maize based on allometric theory.
There have been few investigations of the ARs between different organs of maize and their variation with growth stage, and it is still difficult to measure abiotic stress for the above-mentioned ARs. This study set out to solve these problems by conducting a series of field experiments and by collecting a substantial body of observational data associated with the biomass of different organs, especially, for roots. The objective of this research was to explore ARs between maize organs at different growth stages by using the allometric method and to investigate the response characteristics of ARs to drought during different growth stages of maize and following rehydration based on maize pot experiments for two consecutive years. The main structure of the article is introduced as follows. In materials and methods, experimental design, calculation of irrigation amount and soil water content, parameter expression, and allometric method are expounded. Results consist of “ARs between roots, aboveground and total biomass”, “ARs between the ear and various parts of maize”, and “size-dependence of HI and its response to drought and rewatering”. Discussion includes two parts, i.e., “ARs between maize root and shoot and total plant and their response to drought” and “ARs between the ear and different biomass and their drought responses”. The last section presents the conclusion.
2. Materials and Methods
2.1. Experimental Design
The pot experiment for maize suffering water stress was conducted at the Jinzhou Agricultural Meteorological Experimental Station in Liaoning Province using the Xianyu 335 maize variety, which is a hybrid of middle and late-ripening maize. In the study area, typical brown soil had particle size exceeding 0.05 mm, 0.002 to 0.05 mm, and less than 0.002 mm accounting for 40.73%, 41.56%, and 17.71%, respectively; a pH of 6.3 and contained soil organic matter content of 15.24 g·kg−1, nitrogen of 1.04 g·kg−1, phosphorus of 0.50 g·kg−1 and potassium of 22.62 g·kg−1 [16]. The diameter and height of the experiment PVC textured pots were 40 cm and 100 cm. In the autumn before the experiment year, surface soil up to 20 cm was evenly mixed, weighed, and loaded into the pots. The top 20 cm of soil had an average field capacity of 21.3%, a wilting point of 6.0%, and a bulk density of 1.60 g·cm−3. Three seeds were manually planted into the pots at a 5 cm soil depth, and the seedlings were thinned so that one strong plant remained when the plant had grown its fifth leaf. The experiment consisted of drought treatments (WC) during vegetative (VP) and reproductive (RP) growth periods and corresponding control treatments (CK). There were six replicates per treatment. Natural precipitation was accepted before the jointing of the maize and appropriate water was added when there was not enough precipitation to ensure the normal growth of corn plants. At the jointing stage, six sample pots were selected to observe soil moisture, and the calculated water requirements were added to the pots to reach suitable soil moisture, i.e., a soil relative water content (SRWC) of 75%. After that, a large mobile waterproof shelter was used to prevent natural precipitation and the CK treatment was maintained at suitable soil moisture by water supplementation according to weather conditions and soil situations to ensure normal growth of corn.
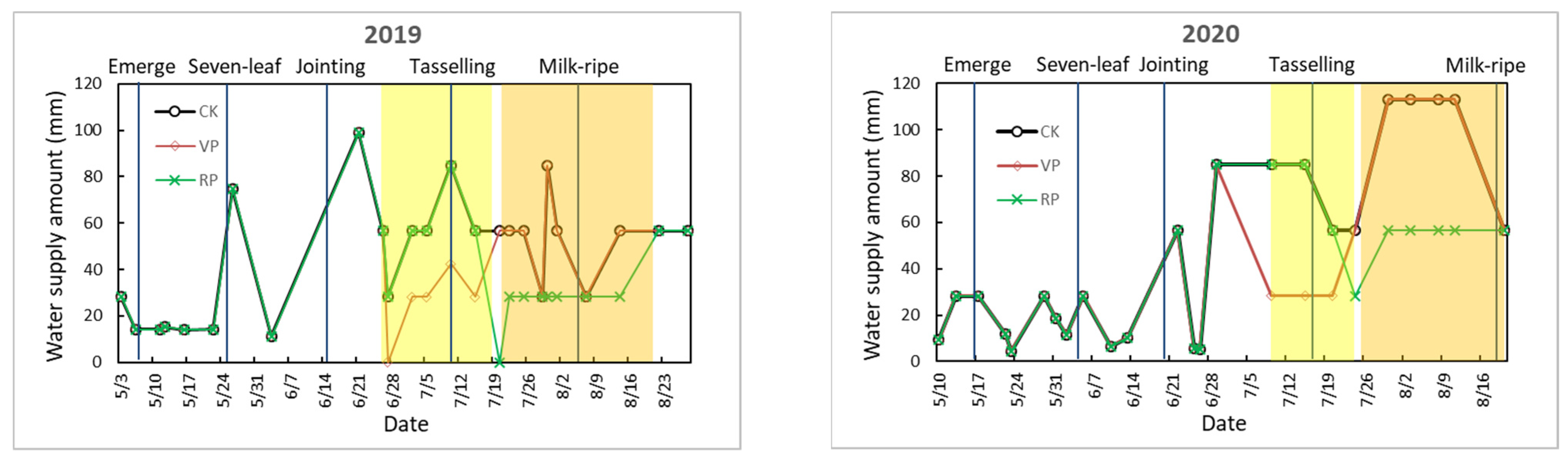
The VP and RP drought treatments were decreasingly irrigated from jointing to silking and from tasseling to milk ripening to form drought episodes, respectively. At the end of VP and RP drought treatments, dry matter weights (DMW) of different organs of maize were measured, and the real-time effects of drought were reflected by comparing the differences in the allometric relationships among various parts of the maize plant between CK and WC treatments. The corresponding WC and CK treatments were defined as VPWC, RPWC, and VPCK, RPCK, respectively. In addition, after the end of VPWC and RPWC, two treatments adopted the same water supply measure with the control until maturity and were respectively defined as VPAWC and RPAWC, and the CK treatment at the end of growth was named CKA. Assuming that the maximum transpiration of a single corn plant during VP and RP is approximately 2000 g/d and 1600 g/d [14], the depth of water replenishment was calculated according to the area of the pot section. The growth period of CK and the water supplement of each treatment are shown in Figure 1.
In 2019, sowing and jointing occurred on 30 April and 15 June, respectively. The VPWC was not irrigated on 27 June and was replenished with half of the water volume of CK from 2 July to 15 July, and irrigated similarly with CK after 20 July. The RPWC was not irrigated on 20 July and was irrigated with half of the water volume of CK from 22 July to 14 August, and irrigated similarly with CK after 22 August. In 2020, sowing and jointing dates were on 10 May and 20 June. The VPWC was irrigated with 33% of the water volume of CK from 9 July to 15 July, with half of the water volume of CK on 20 July, and similarly with CK from 24 July. The RPWC was irrigated with half of water volume of CK from 24 July to 11 August, and irrigated consistently with CK after 20 August. The abbreviations for the different experimental treatments and variables are summarized in Table 1.
2.2. Methods
2.2.1. Calculation of Irrigation Volume
According to the current soil water content, the expected water content can be achieved through water replenishment. The formula used was as follows.
where, stands for supply water quantity (mm), and are thickness (m) and bulk density (g·cm−3) of soil layer i, is the referenced soil relative water content or 75% in this study. and are field capacity (g·g−1) and soil weight water content (g·g−1) of soil layers i and n, equal to 9 in this research, refers to the number of soil layers, which is defined with a thickness of 10 cm per layer. Based on this method, the required volume of water replenishment for experimental samples at the jointing stage was calculated according to the measured soil moisture.
2.2.2. Soil Water Content Measurement
Soil water content was expressed by SRWC and was calculated as follows:
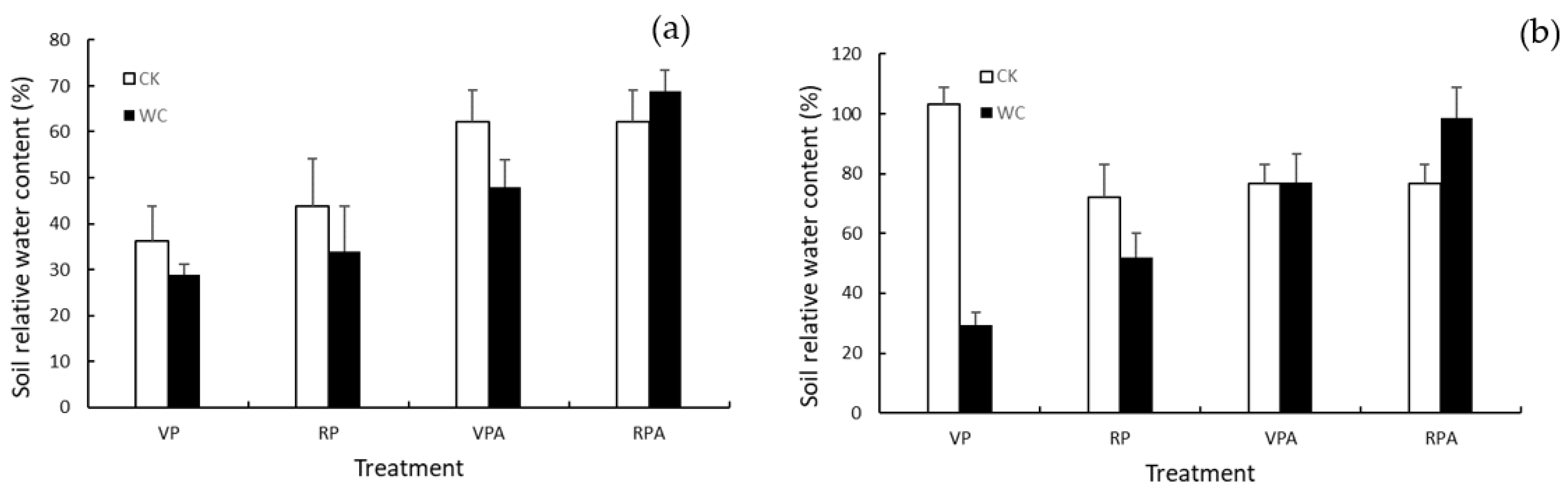
where is SRWC and was measured at the end of different treatments each year. As shown in Figure 2, the SRWC of CK was higher than those of WC after the drought in 2019 but failed to reach the floor level of 60%, due to a deficient water supply. Even after rehydration, the SRWC of VPAWC did not reach 60% of SRWC. Conversely, the SRWC of CK in 2020 reached the appropriate level after drought and rehydration. It is worth noting that the SRWC of RPAWC was higher than that of the corresponding CK in both years. The reason was that the physiological functions of plants were disrupted due to RP drought, and the plants withered after rewatering, which decreased the water consumption of transpiration. In addition, the measured soil samples were in the outer layer of the soil column, and were drier than those in the inside of the soil column, resulting in a lower value relative to the real condition.
2.2.3. Parameter Expression and Allometric Method
RSR and HI are expressed as Equations (3) and (4):
where DMr, DMe, and DMs represent the dry matter weight of root, ear, and aboveground organs. AR can be described as:
RSR = DMr/DMs
HI = DMe/DMs
To facilitate parameter fitting, the equation is logarithmically converted into a linear relationship.
where DMy and DMx represent the masses of organs y and x, respectively. is an allometric coefficient. reflects the relative growth ratio between y and x, equaling 1 when different organs are in a steady growth status, and deviation from 1 means that it is influenced by changes in the environment or developmental period [2].
2.3. Data Statistics
The observational data for each year were statistically analyzed using SPSS 26.0 software (SPSS Inc., Chicago, IL, USA, URL:https://www.ibm.com/cn-zh/spss?lnk=flatitem (accessed on 2 March 2021).
3. Results
3.1. ARs between Roots, Aboveground, and Total Biomass
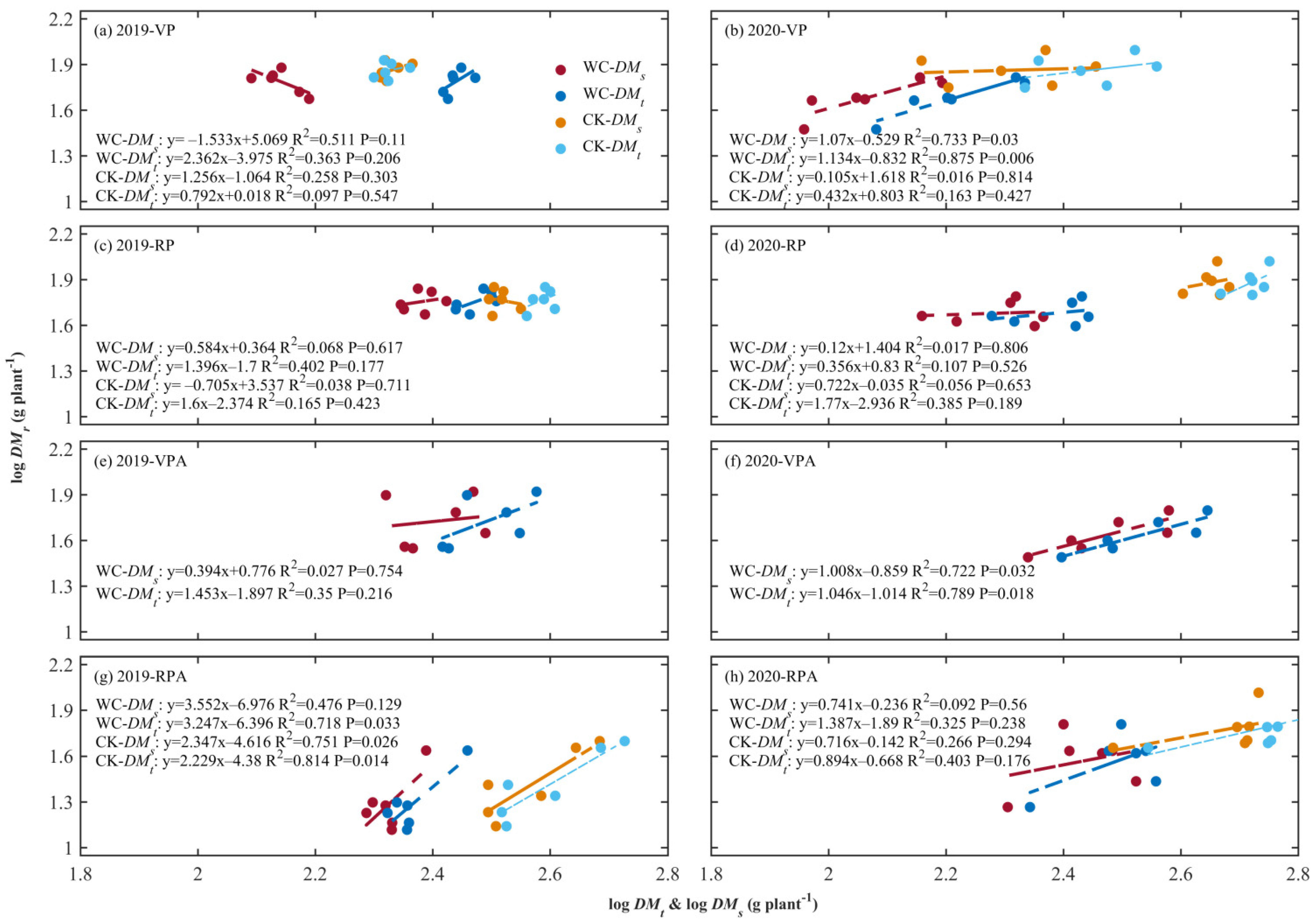
Logarithmic transformation of the dry matter weight of roots, and aboveground organs i.e. the shoot and total plant were conducted and named as logDMr, logDMs, and logDMt. Their relationships were established to reflect ARs in Figure 3.
In 2019, at the end of VPWC, the logDMr of CK showed a positive correlation with the logDMs and logDMt with a relative growth ratio larger than 1 for the former, meaning that the growth rate of roots was greater than that of shoot. There was a negative correlation between logDMr and logDMs for WC, indicating that they had opposite growth trends and the growth rate of the roots was higher than that of the shoot, but there was a positive correlation between logDMr and logDMt. In 2020, the logDMr and logDMt all showed a positive correlation for CK and WC, and the relationship for WC was more significant (p < 0.01) with a relative growth rate greater than 1, indicating that the root growth was faster than that of the total plant, while the growth rate of the roots was lower than that of total plant for CK. The relationship between logDMr and logDMs was not obvious for CK and significant for WC (p < 0.05). The DMt of VPCK in 2019 and 2020 was 213.3 ± 10.6 and 284.3 ± 57.7 g∙plant−1, respectively, indicating that the VPCK in 2019 was also affected by drought. The ARs for CK between roots and shoot and total plant in 2019 were stronger than those in 2020, indicating that drought promoted AR in this period. At the end of RPWC, there was a slight positive correlation between logDMr and logDMt for CK, and AR was more obvious for WC, but there was no reliable positive correlation between logDMr and logDMs for WC and CK in 2019, while only logDMr and logDMt for CK in 2020 had slightly positive correlativity.
At the end of the growth in 2019, DMs of 257.4 ± 40.8 g∙plant−1 was smaller than that in 2020 of 303.0 ± 65.9 g∙plant−1, reflecting that after effect of VPCK in 2019 was persistent. Specifically, in 2019, the ARs between roots and shoot and total plant were significant for CKA and the root-total plant AR was more significant (p < 0.05). There was no AR between root and shoot and positive AR between root and total plant was slight with a large variability for VPAWC. Inversely, the ARs of root-total plant and root-shoot both were obvious and the former (p < 0.05) was more significant than the latter for RPAWC. In 2020, the ARs between root, shoot and total plant both showed slightly positive correlativity for CKA and were strengthened by the previous VPWC with the AR of the root-total plant being more significant than that of root-shoot (p < 0.05), while the ARs were weakened by the previous RPWC.
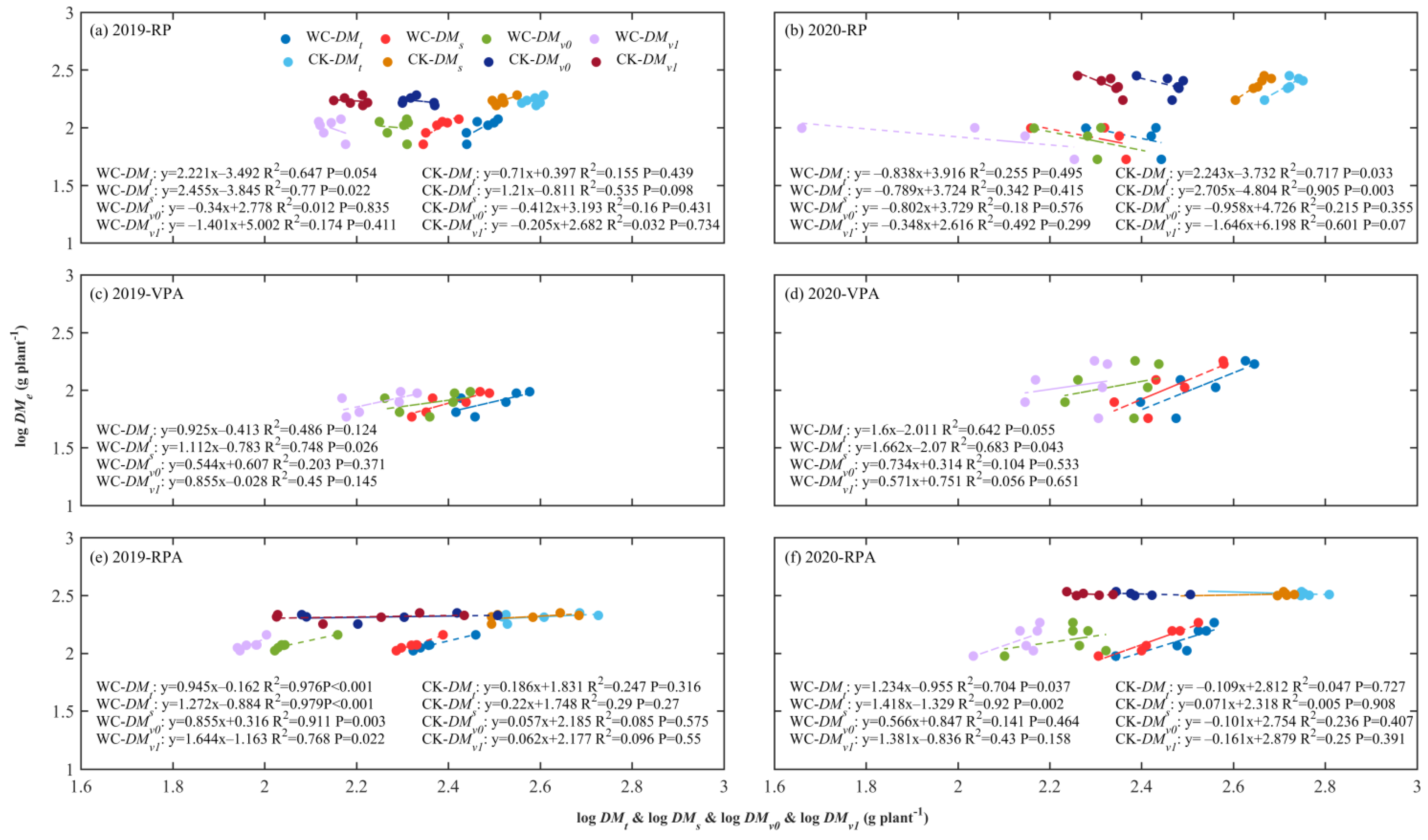
3.2. ARs between Ear and Various Parts of Maize
The ARs between the ear and various parts including total plant, shoot, total plant minus ear (DMv0) and shoot minus ear (DMv1) were shown in Figure 4. At the end of RPWC, the log-transformed ear (logDMe) was positively correlated with logDMs and logDMt in 2019 and 2020, and the relationships between logDMe and logDMs were very significant with p < 0.1 and p < 0.01 in 2019 and 2020, respectively. But logDMe was negatively correlated with log-transformed DMv0 and DMv1 for CK in 2019 and 2020, and the correlations were more significant in 2020 than in 2019. Under the impact of RPWC, the ear had significant ARs with the shoot (p < 0.05) and total plant (p < 0.1) and had no ARs with DMv0 and DMv1 in 2019, and had no ARs with four parts in 2020. In general, the AR between the ear and shoot was the most significant, but the AR between the ear and the vegetative organ was not significant.
At the end of growth, there were no obvious ARs between the ear and four parts in 2019 and 2020 for CK. Under the effects of VPWC and RPWC, the ear showed positive ARs with all four parts, and the ARs between the ear and shoot and total plant were the most significant in 2019 (p < 0.001 and p < 0.001 ) and 2020 (p < 0.001 and p < 0.05) for RPAWC, and were less significant for VPAWC than for RPAWC.
3.3. Size-Dependence of HI and Its Response to Drought and Rewatering
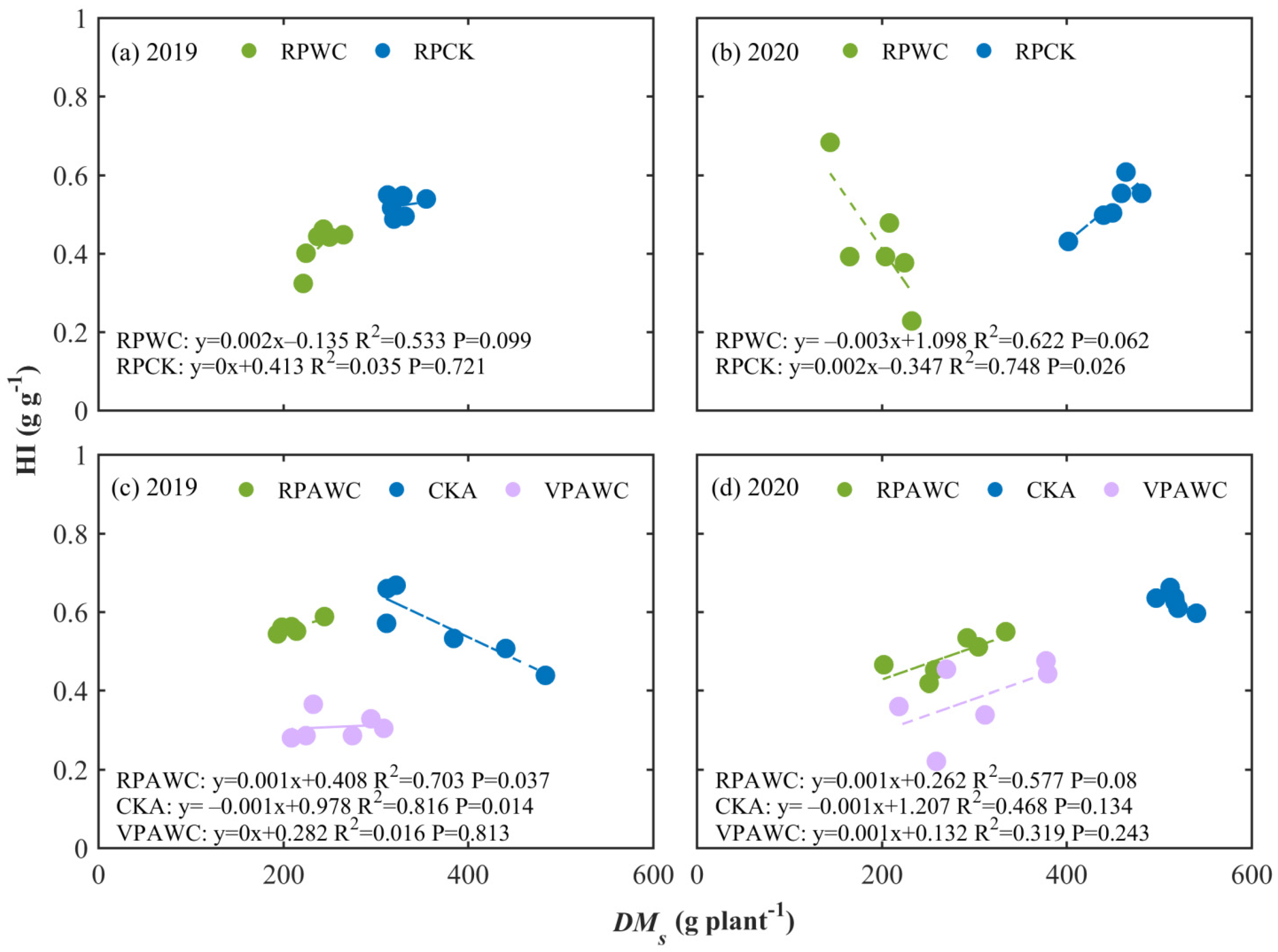
Figure 5 shows the relationships between HI and the size of maize plants under different water treatments. In terms of the condition at the end of the RPWC, the correlation between HI and DMs was not obvious for CK but positively significant for WC (p < 0.1) in 2019. The situation in 2020 was the opposite with the correlation for CK significantly positive (p < 0.05) and negative for WC (p < 0.1). In general, HI was not a constant and was related to plant size under different soil water situations and drought made HI smaller than the control value.
From rewatering after drought to the end of the growth period in the two years, the HI of plants experiencing RPWC was slightly smaller than that of CK and was larger than that affected by VPWC. The correlations between HI and DMs for CK were negative and significant in 2019 (p < 0.05) and they were significantly positive in 2019 (p < 0.05) and in 2020 (p < 0.1) for RPAWC and not significant for VPAWC.
4. Discussion
4.1. ARs between Maize Root and Shoot and Total Plant and Their Response to Drought
Some studies have argued that the correlation between vegetative organs and the total plant is incorrect because the latter includes the former [21] and some scholars believe that vegetative organs, reproductive organs, and the total plant are all related to each other. This study further clarified the above relationships by analyzing the ARs of root and shoot and total plant and their responses to drought. The AR of the root-total plant was stronger than that of the root-shoot in different treatments and periods, which was closely related to the fact that the root was included in the total plant.
Further analyzing showed that the ARs of root and shoot and total plant for VPCK and RPCK were weaker than those for CKA, reflecting that the ARs gradually strengthened as the growth process approached the end. The VP drought promoted the AR between roots and total plant, and the AR between roots and shoot for VPWC was negatively correlated in 2019, suggesting that a certain degree of drought promoted root growth while inhibiting shoots. In addition, the ARs for CKA in 2019 were stronger than those in 2020 because VPCK for 2019 suffered drought and the ARs for VPAWC in 2020 were more significant than those in 2019, which corresponded to the fact that drought was more severe in 2019 than in 2020, showing that severe drought can weaken the AR, but the drought in a certain range can increase AR. The AR for RPAWC was stronger in 2019 than in 2020, reflecting that the more severe previous drought played a more vital role in strengthening the ARs at the end of the growth period, which is also related to the occurrence time of drought during growth [14]. The root-shoot ARs showed that the growth rates of roots for most treatments were larger than those of shoot, which is different from the conclusion of Zhang et al. [10] for two reasons. The first is related to the differences in varieties and cultivation zones, and the second is that roots were only observed above the depth of 30 cm in the study without considering total roots.
Root-shoot ratio (RSR) is an important indicator of crop yield. Variation characteristics of RSR can be reflected by analyzing the AR between roots and shoot. The obvious ARs for VPWC and VPAWC in 2020 and for RPVWC and CKA in 2019 reflected that RSR was not constant under these conditions, but closely related to individual size. The shifting ARs for VPWC and RPAWC varied the size-dependence of RSR. These results indicated that the functional relationship between the dry matter weight of each part and plant size under drought was related to the difference in organ growth rate in response to drought [8], so plant growth and size should be considered to explain the dry matter distribution process relative to fixed allocation rates [10]. Specifically, differences in environmental conditions can be combined with biomass allocation patterns related to total mass to understand how plants allocate biomass in response to changes in individual size and environment [22].
4.2. ARs between Ear and Different Biomass and Their Drought Responses
Regardless of CK and WC at the end of drought treatment, the ARs between ear and DMv0 and DMv1 were significantly less than those between the ear and DMs and DMt, and the AR of ear–DMs was the most significant. Under normal water supply, the ARs were more significant during the growing process than at the end of growth, and ears and vegetative organs had negative ARs, reflecting that the two parts competed for photosynthate. However, drought enhanced ARs between the ear and DMs, DMt, and even vegetative organs, which reflected the contribution of dry matter redistribution of vegetative organs to the ear biomass accumulation [1]. At the end of growth, the ear biomass for CK showed negligible change with different parts’ biomasses and was not size-dependent, indicating that the ear was sufficiently supplied with photosynthate and the excess photosynthate was allocated to the vegetative organs, increasing the individual difference. Under the effect of the previous drought, the ARs were strengthened relative to those for CK, indicating that the limited photosynthate was supplied for vegetative organs according to a certain proportion after the accumulation of ear dry matter was ensured, showing that drought in different developmental periods can promote ARs. The significant ARs between ear and vegetative organs further reflected the contribution of dry matter redistribution from vegetative organs to ear at the late growth stage.
The constant HI is often used to estimate yield by multiplying it with aboveground dry matter weight and it is an important parameter to measure crop productivity [23]. However, it changed with the developmental stage [24]. In this study, under normal conditions, the AR between ear and DMs was significant during the growing process, reflecting the source–sink relationship of dry matter allocation of different organs [24], while it was nonexistent at the end of growth, meaning that source–sink relationship weakened, also indicating that the correlations between reproductive output and individual size varied with growth stages [24]. Compared with the conclusion in the literature [10], the ear–shoot ARs in this research are credible as a whole except for the smaller sample size, which further confirmed the size-dependence of the ear on maize plants. Under the effect of drought occurring at different growth periods, reproductive output was correlated with plant size, and its size-dependence was enhanced, reflecting that HI varied with individual size, which further proved that drought affected biomass allocation patterns [2,18].
Compared with the allocation ratio, the biomass allocation pattern can be better understood by considering growth and individual size [10] and AR will give a more reasonable explanation for the change of HI [8]. Under normal conditions, the relationships between HI and DMs were positively correlated during the growth process, meaning that the whole plant provided an important material basis for ear dry matter accumulation, while was negatively correlated at the later stage of growth, illustrating that the ratio of dry matter transformed from vegetative organs to reproductive organs decreased with increasing plant size and the greater the aboveground biomass, the smaller the HI. Drought varied the relationships between HI and plant size. Under RP drought during the growing process, the relationship obviously varied relative to CK, and HI decreased with the increase of DMs. At a later growth stage, under the effect of the previous drought during VP and RP, HIs were positively correlated with individual size and the relationship was more significant under RP drought than under VP drought, indicating that RP drought reinforced the role of size in increasing HI and an aftereffect was induced by the previous drought. Increasing plant size is beneficial to obtain higher yields under environmental stress [24,25].
5. Conclusions
The present study indicated that the root had a more significant AR with the total plant than with the shoot of maize. Under normal conditions, the ARs of root-total plant and root-shoot gradually increased with the growth process approaching an endpoint. The droughts at different growth stages exerted a stimulatory effect on the above ARs during the growth process and induced different aftereffects on the ARs at the end of growth after rewatering. The VP drought within a certain range intensified the ARs during the growth process and at the end of growth and increased the size-dependence of RSR, while severe drought broke the AR at the end of growth. The RP drought inhibited the above ARs and the size-dependence of RSR at the end of growth. The AR of the ear–shoot during the growth process was more significant than that at the end of growth under normal conditions. Drought strengthened the ear–shoot AR at the two periods and varied the size-dependence of HI. The RP drought played a more evident role in promoting this AR and the relationship between HI and aboveground biomass relative to VP drought at the end of growth.
Author Contributions
H.M., F.C. and Y.Z. conceived and designed the experiments; H.Z., X.Z. and Y.F. performed the experiments; H.M., F.C., N.M. and S.Z. analyzed the data; H.M. drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (Grant number: 41775110, 41975149, and 42275202), the LiaoNing Revitalization Talents program (Grant number: XLYC1807262), and the foundation of China meteorological administration Shenyang institute of atmospheric environment (Grant number: 2022SYIAEJY12).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors are grateful to Hang Wu, Bingbing Zhang, Quan Gao, Lili Gao, Hao Zhang and Likun Zhou for their work during the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weiner, J.; Campbell, L.G.; Pino, J.; Echarte, L. The allometry of reproduction within plant populations. J. Ecol. 2009, 97, 1220–1233. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phycol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Sun, Z.X.; Zheng, J.M.; Wang, W.B.; Bai, W.; Feng, L.S.; Yang, N.; Xiang, W.Y.; Zhang, Z.; Feng, C. Dry Matter Accumulation, Allocation, Yield and Productivity of Maize Soybean Intercropping Systems in the Semi-Arid Region of Western Liaoning Province. Sci. Agric. Sin. 2021, 54, 909–920. [Google Scholar]
- Lizaso, J.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.; Lorite, I.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crops Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Kruse, J.; Hänsch, R.; Mendel, R.R.; Renneberg, H. The role of root nitrate reduction in the systemic control of biomass partitioning between leaves and roots in accordance to the C/N-status of tobacco plants. Plant Soil. 2010, 332, 387–403. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Shipley, B.; Meziane, D. The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Funct. Ecol. 2002, 16, 326–331. [Google Scholar] [CrossRef]
- Qin, X.L.; Weiner, J.; Qi, L.; Xiong, Y.C.; Li, F.M. Allometric analysis of the effects of density on reproductive allocation and Harvest Index in 6 varieties of wheat (Triticum). Field Crops Res. 2013, 144, 162–166. [Google Scholar] [CrossRef]
- Ratjen, A.M.; Lemaire, G.; Kage, H.D.; Justes, P.E. Key variables for simulating leaf area and N status: Biomass based relations versus phenology driven approaches. Eur. J. Agron. 2018, 100, 110–117. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Yu, K.L.; Jin, X.L.; Nan, Z.B.; Wang, J.F.; Niu, X.L.; Whish, J.P.M.; Bell, L.W.; Siddique, K.H.M. Above- and belowground dry matter partitioning of four warm-season annual crops sown on different dates in a semiarid region. Eur. J. Agron. 2019, 109, 125918. [Google Scholar] [CrossRef]
- Kage, H.; Kochler, M.; Stützel, H. Root growth and dry matter partitioning of cauliflower under drought stress conditions: Measurement and simulation. Eur. J. Agron. 2004, 20, 379–394. [Google Scholar] [CrossRef]
- Kumar, R.; Sarawgi, A.K.; Ramos, C.; Amarante, S.T.; Ismail, A.M.; Wade, L.J. Partitioning of dry matter during drought stress in rainfed lowland rice. Field Crops Res. 2006, 96, 455–465. [Google Scholar] [CrossRef]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.S.; Mi, N.; Ming, H.Q.; Zhang, S.J.; Zhang, H.; Zhao, X.L. Maize (Zea mays L.) physiological responses to drought and rewatering, and the associations with water stress degree. Agric. Water Manag. 2020, 241, 106479. [Google Scholar] [CrossRef]
- Djaman, K.; Irmak, S.; Rathje, W.R.; Martin, D.L.; Eisenhauer, D.E. Maize evapotranspiration, yield production functions, biomass, grain yield, harvest index, and yield response factors under full and limited irrigation. Trans. ASABE 2013, 56, 373–393. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.S.; Mi, N.; Ming, H.Q.; Zhang, S.J.; Zhang, H.; Zhao, X.L.; Zhang, B.B. The Effect of Drought and Sowing Date on Dry Matter Accumulation and Partitioning in the Above-Ground Organs of Maize. Atmosphere 2022, 13, 677. [Google Scholar] [CrossRef]
- Mi, N.; Cai, F.; Zhang, Y.S.; Ji, R.P.; Zhang, S.J.; Wang, Y. Differential responses of maize yield to drought at vegetative and reproductive stages. Plant Soil Environ. 2018, 64, 260–267. [Google Scholar]
- Wang, X.C.; Yang, W.Y.; Deng, X.Y.; Zhang, Q.; Yong, T.W.; Liu, W.G.; Yang, F.; Mao, S.M. Differences of dry matter accumulation and distribution of maize and their responses to nitrogen fertilization in maize/soybean and maize/sweet potato relay intercropping systems. Plant Nutr. Fertil. Sci. 2015, 21, 46–57. [Google Scholar]
- Wei, T.B.; Hu, F.L.; Zhao, C.; Feng, F.X.; Yu, A.Z.; Liu, C.; Chai, Q. Response of dry matter accumulation and yield components of maize under n-fertilizer postponing application in oasis irrigation areas. Sci. Agric. Sin. 2017, 50, 2916–2927. [Google Scholar]
- Anothai, J.; Soler, C.M.T.; Green, A. Evaluation of two evapotranspiration approaches simulated with the CSM-CERES Maize model under different irrigation strategies and the impact on maize growth, development and soil moisture content for semi-arid conditions. Agric. For. Meteorol. 2013, 176, 64–76. [Google Scholar] [CrossRef]
- Brett, M.T. When is a correlation between non-independent variables “spurious”? Oikos 2004, 105, 647–656. [Google Scholar] [CrossRef]
- McCarthy, M.C.; Enquist, B.J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Maddonni, G.A. Analysis of the climatic constraints to maize production in the current agricultural region of Argentina—A probabilistic approach. Theor. Appl. Climatol. 2012, 107, 325–345. [Google Scholar] [CrossRef]
- Bonelli, L.E.; Monzon, J.P.; Cerrudo, A.; Rizzalli, R.H. Maize grain yield components and source-sink relationship as affected by the delay in sowing date. Field Crops Res. 2016, 198, 215–225. [Google Scholar] [CrossRef]
- Cerrudo, A.; Di Matteo, J.; Fernandez, E.; Robles, M.; Pico, L.O.; Andrade, F.H. Yield components of maize as affected by short shading periods and thinning. Crop Pasture Sci. 2013, 64, 580–587. [Google Scholar] [CrossRef]
Figure 1.
Dates (month/day) of maize growth periods and irrigation regimes for the different experimental treatments in 2019 and 2020. The light and deep yellows stand for the periods of VPWC and RPWC, respectively.
Figure 1.
Dates (month/day) of maize growth periods and irrigation regimes for the different experimental treatments in 2019 and 2020. The light and deep yellows stand for the periods of VPWC and RPWC, respectively.

Figure 2.
Soil moisture of WC and CK for different treatments in 2019 (a) and 2020 (b).
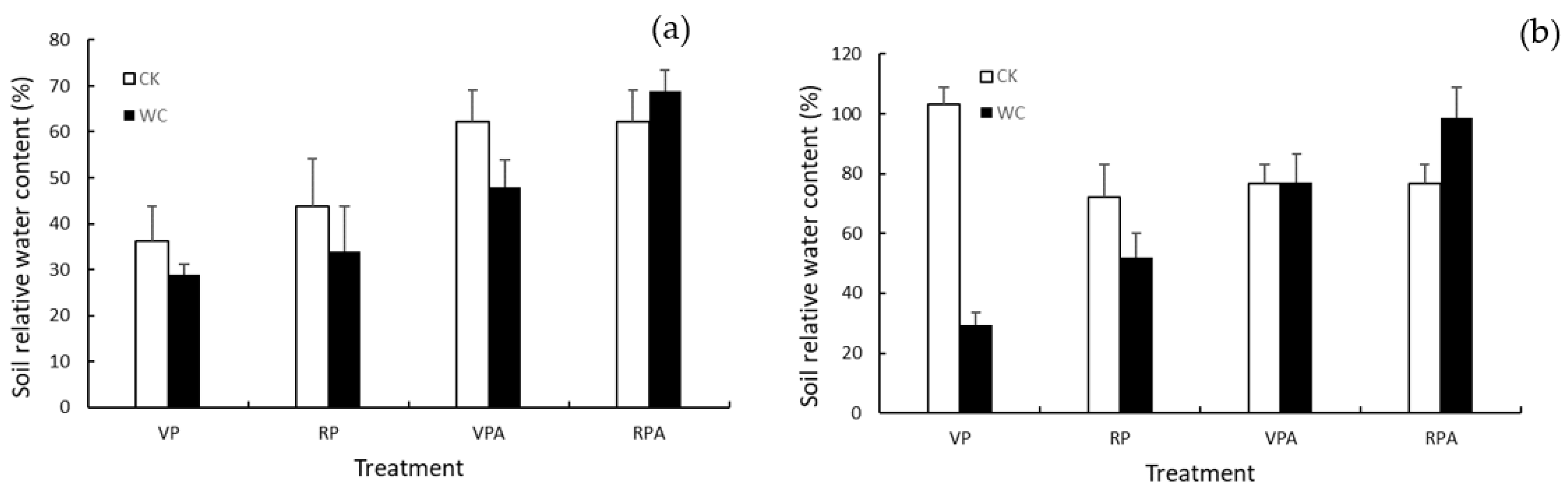
Figure 3.
The ARs between root and shoot and total plant in 2019 and 2020.
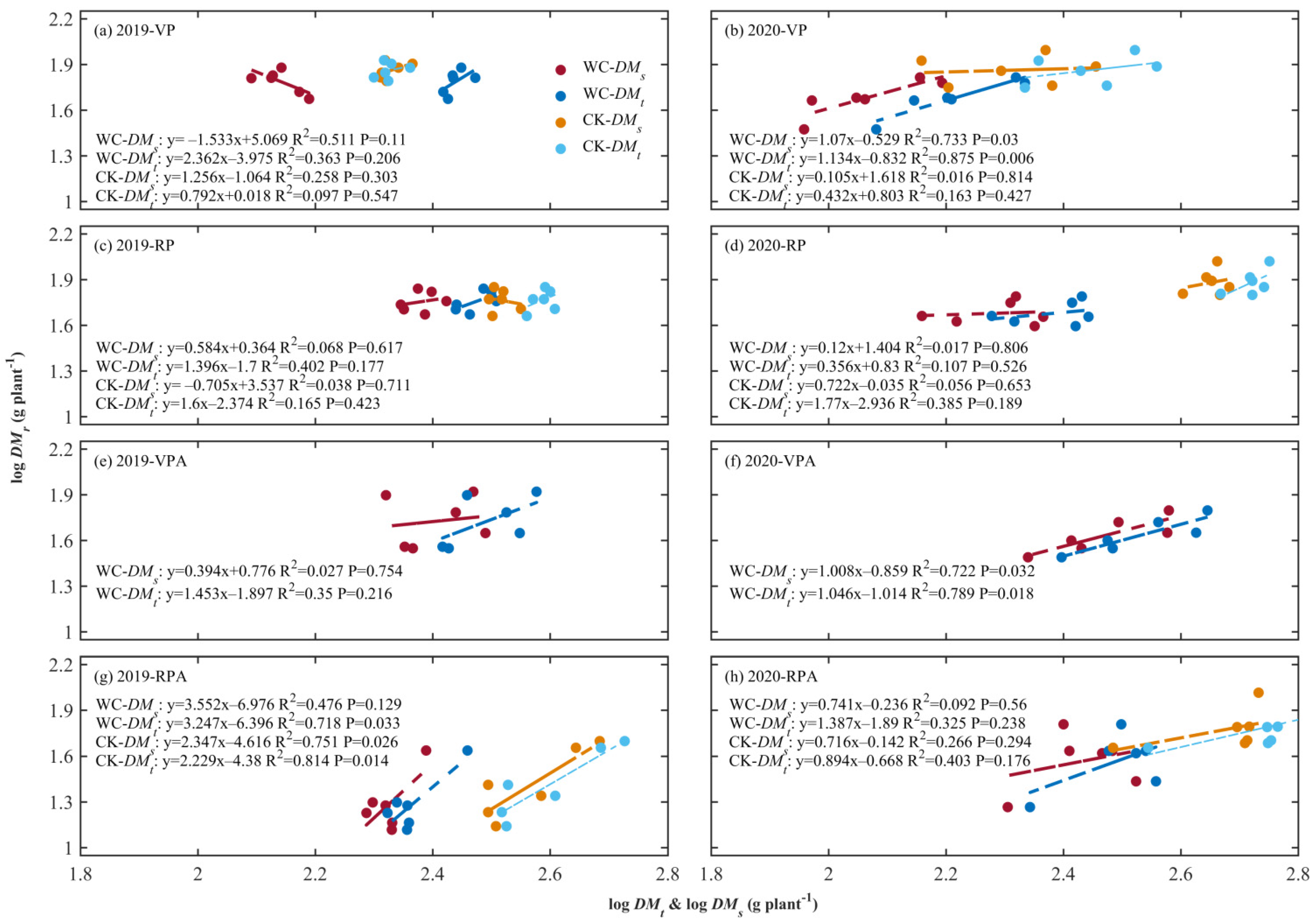
Figure 4.
The ARs between ear and various parts of maize in 2019 and 2020.

Figure 5.
Relationships between HI and shoot during growth (a,b) and at the end of growth (c,d) in 2019 and 2020.
Figure 5.
Relationships between HI and shoot during growth (a,b) and at the end of growth (c,d) in 2019 and 2020.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Abbreviations used to denote each parameter and treatment.
| Abbreviation | Description |
|---|---|
| AR | Allometric relationship |
| DMP | Dry matter partitioning |
| DMW | Dry matter weight |
| RSR | Root-shoot ratio |
| HI | Harvest index |
| SRWC | Soil relative water content |
| CK | The control treatment |
| WC | Water control treatment |
| VP | Vegetative period of maize |
| RP | Reproductive period of maize |
| VPWC, VPCK | WC during VP and its CK |
| RPWC, RPCK | WC during RP and its CK |
| VPAWC, RPAWC | VPWC and RPWC irrigated until maturity |
| CKA | The CK for VPAWC and RPAWC |
| DMr, DMe, DMs, and DMt | DMWs of root, ear, aboveground organ, and total plant |
| DMv0, DMv1 | DMWs of total plant minus ear and shoot minus ear |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ming, H.; Cai, F.; Mi, N.; Zhang, Y.; Zhang, H.; Zhang, S.; Zhao, X.; Feng, Y. Allometric Relationships between Dry Matter Weights of Maize Organs and Their Responses to Drought. Water 2023, 15, 99. https://doi.org/10.3390/w15010099
AMA Style
Ming H, Cai F, Mi N, Zhang Y, Zhang H, Zhang S, Zhao X, Feng Y. Allometric Relationships between Dry Matter Weights of Maize Organs and Their Responses to Drought. Water. 2023; 15(1):99. https://doi.org/10.3390/w15010099
Chicago/Turabian StyleMing, Huiqing, Fu Cai, Na Mi, Yushu Zhang, Hui Zhang, Shujie Zhang, Xianli Zhao, and Yanrui Feng. 2023. "Allometric Relationships between Dry Matter Weights of Maize Organs and Their Responses to Drought" Water 15, no. 1: 99. https://doi.org/10.3390/w15010099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.