
Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters



Abstract
:1. Introduction
2. Material and Methods
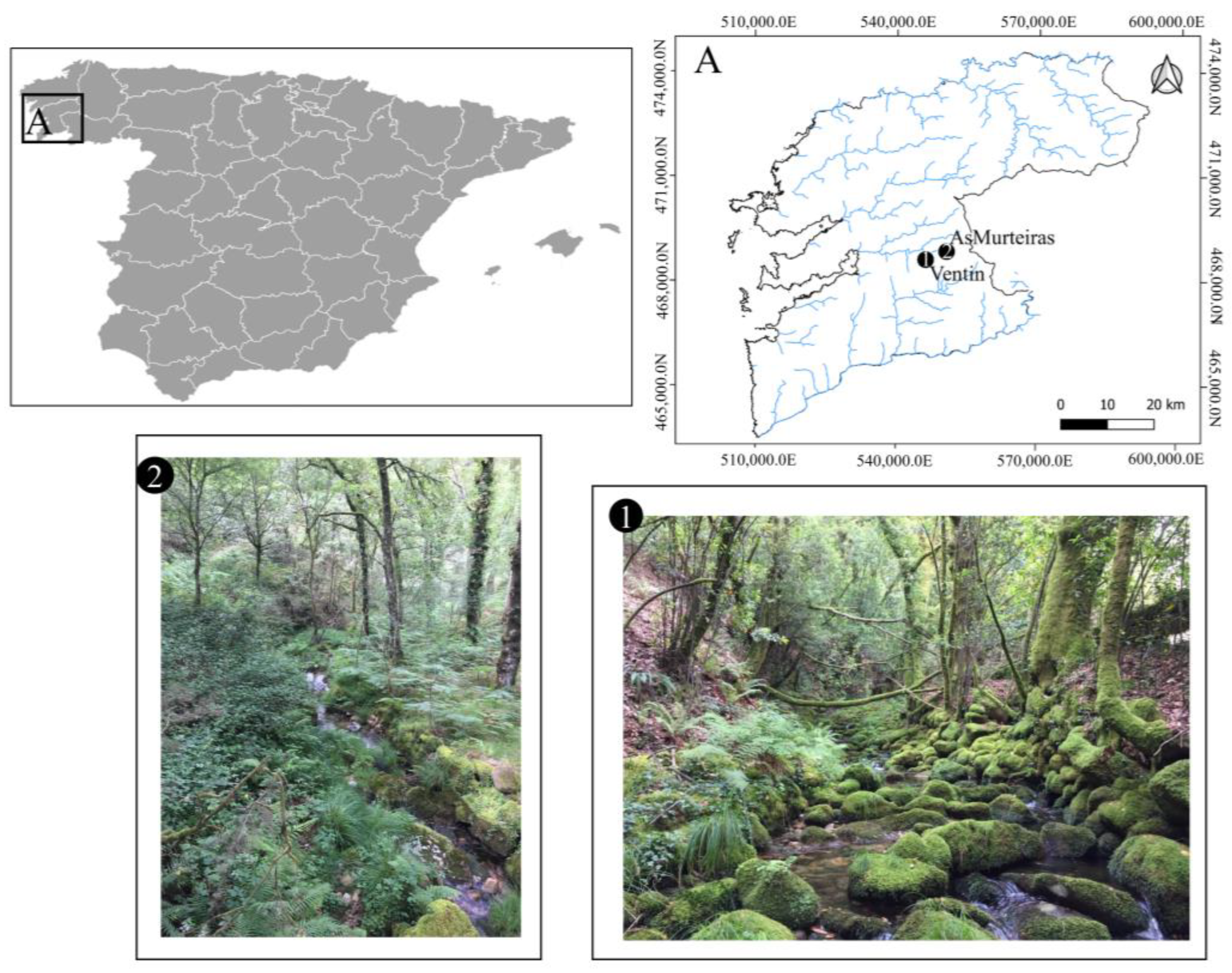
2.1. Studied Region
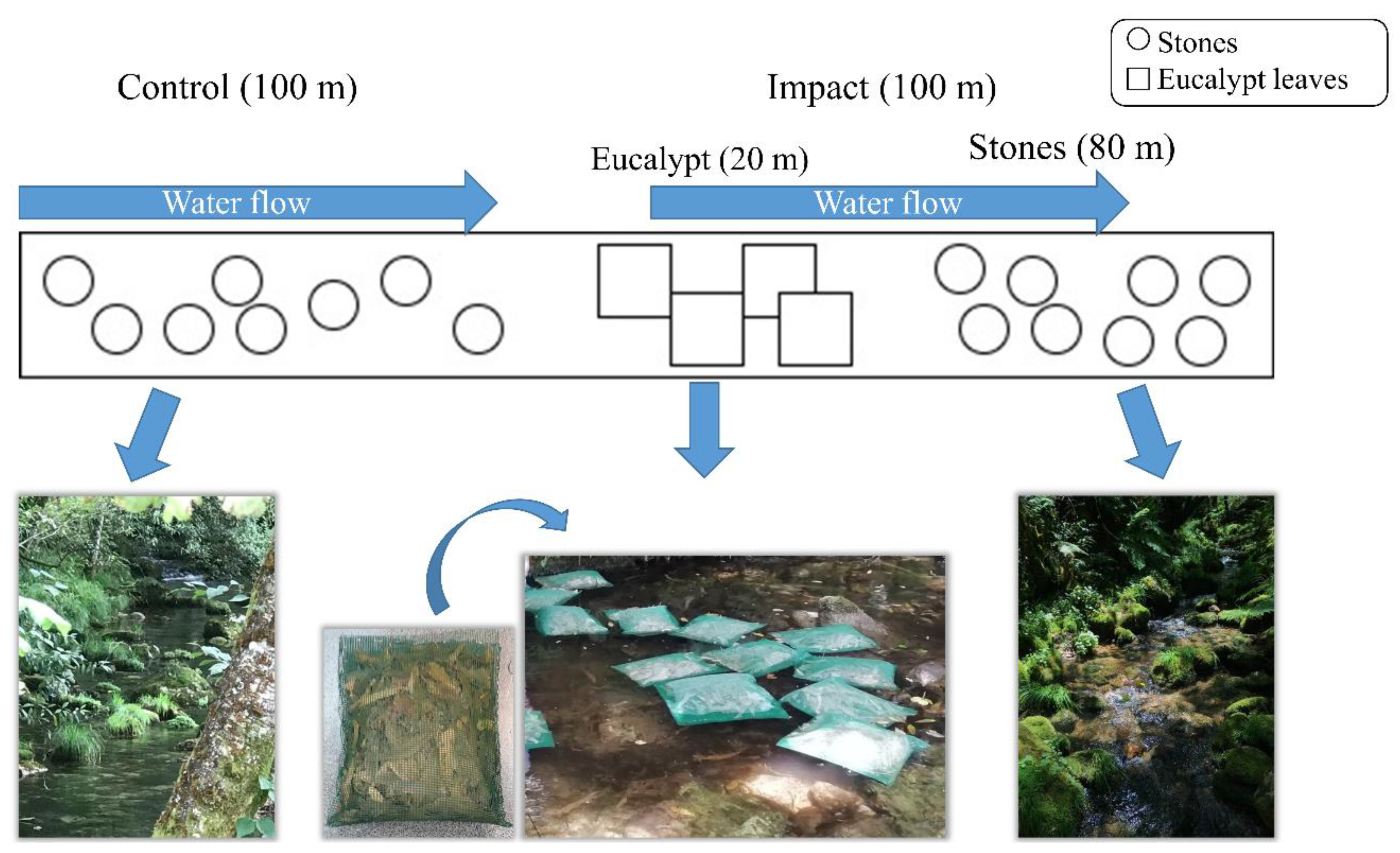
2.2. Experimental Design
2.3. Field and Laboratory Work
2.4. Statistical Analyses
3. Results
3.1. Physico-Chemical Variables
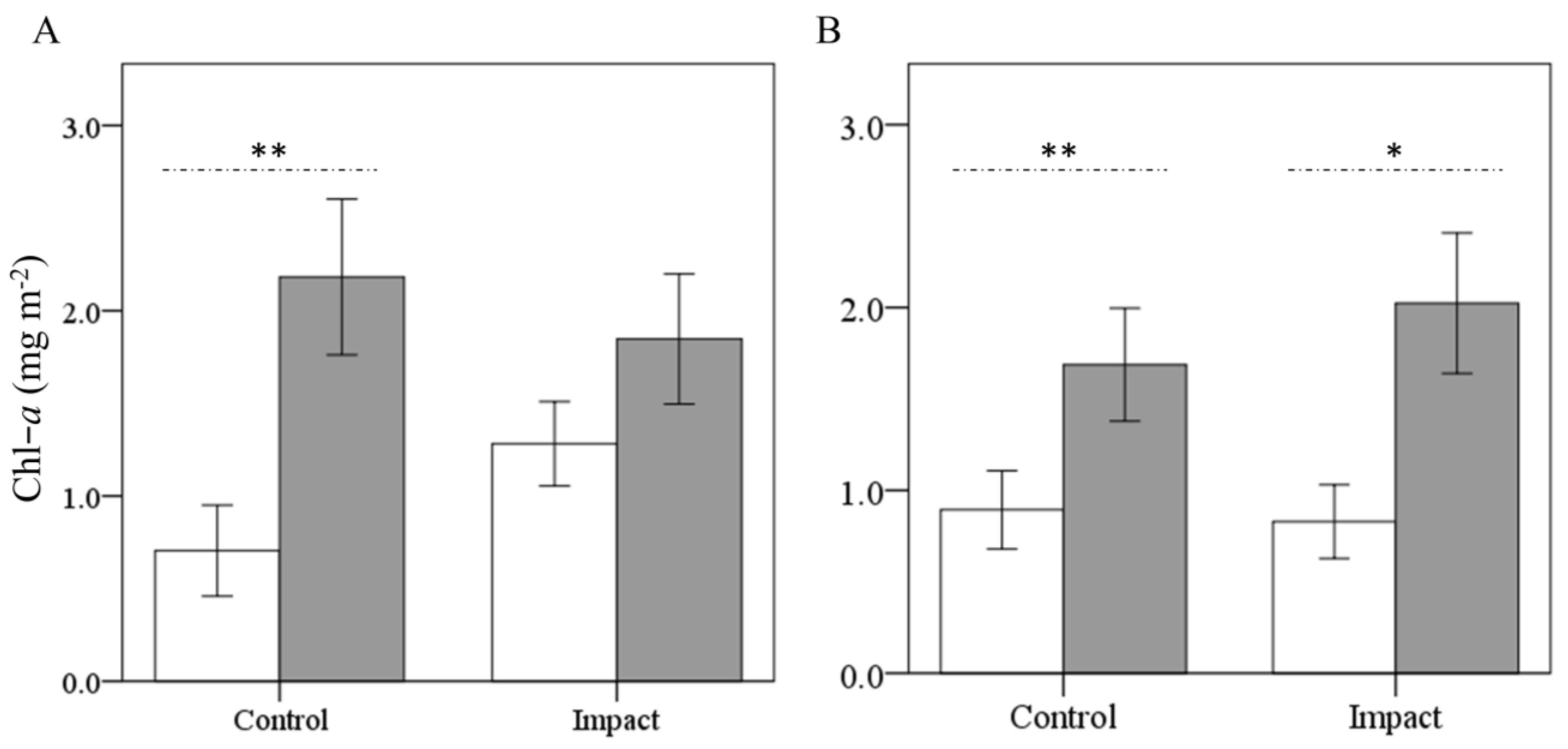
3.2. Chlorophyll a
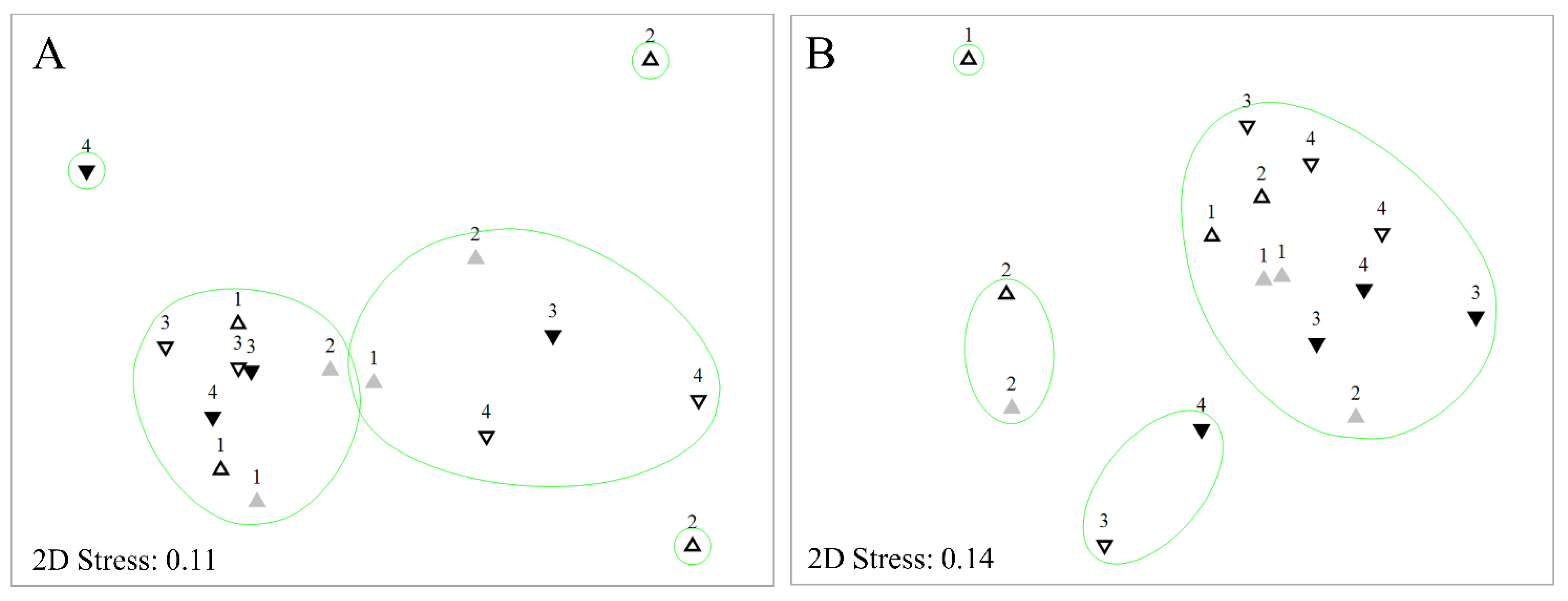
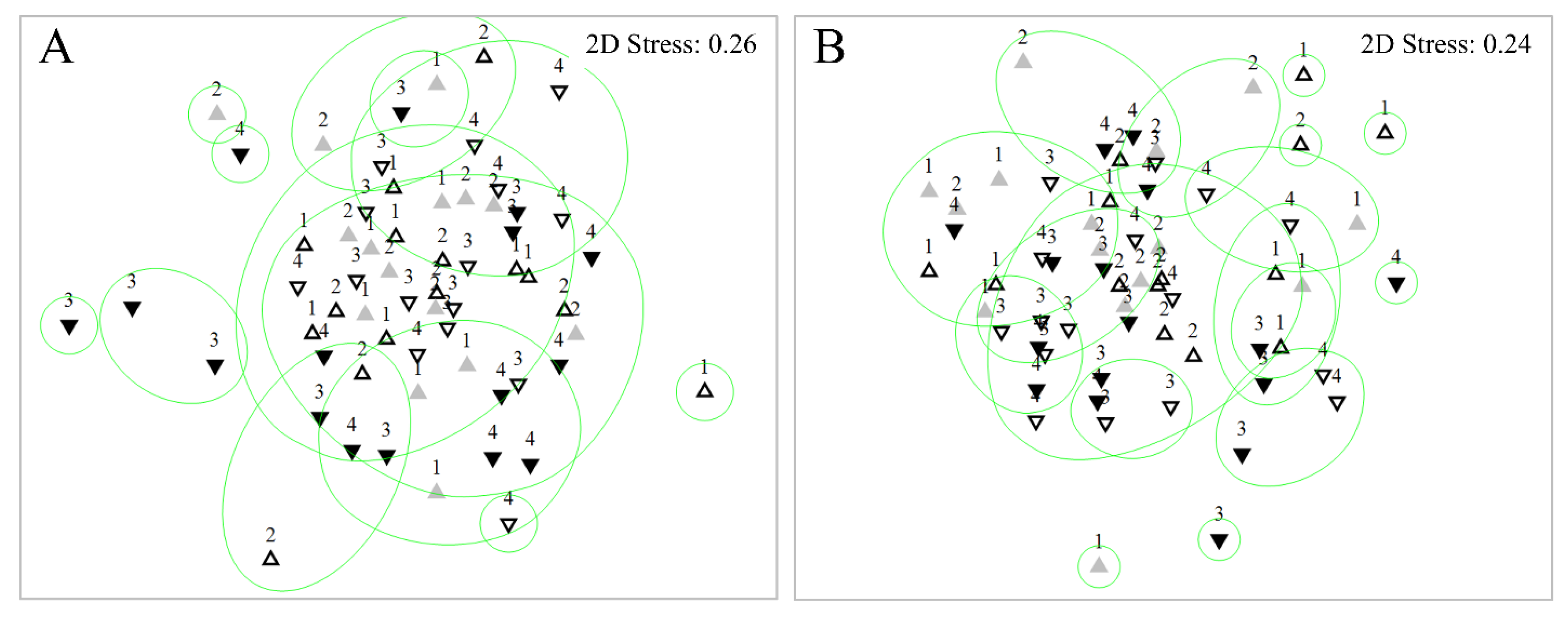
3.3. Diatom Assemblages
3.3.1. Diatom Composition
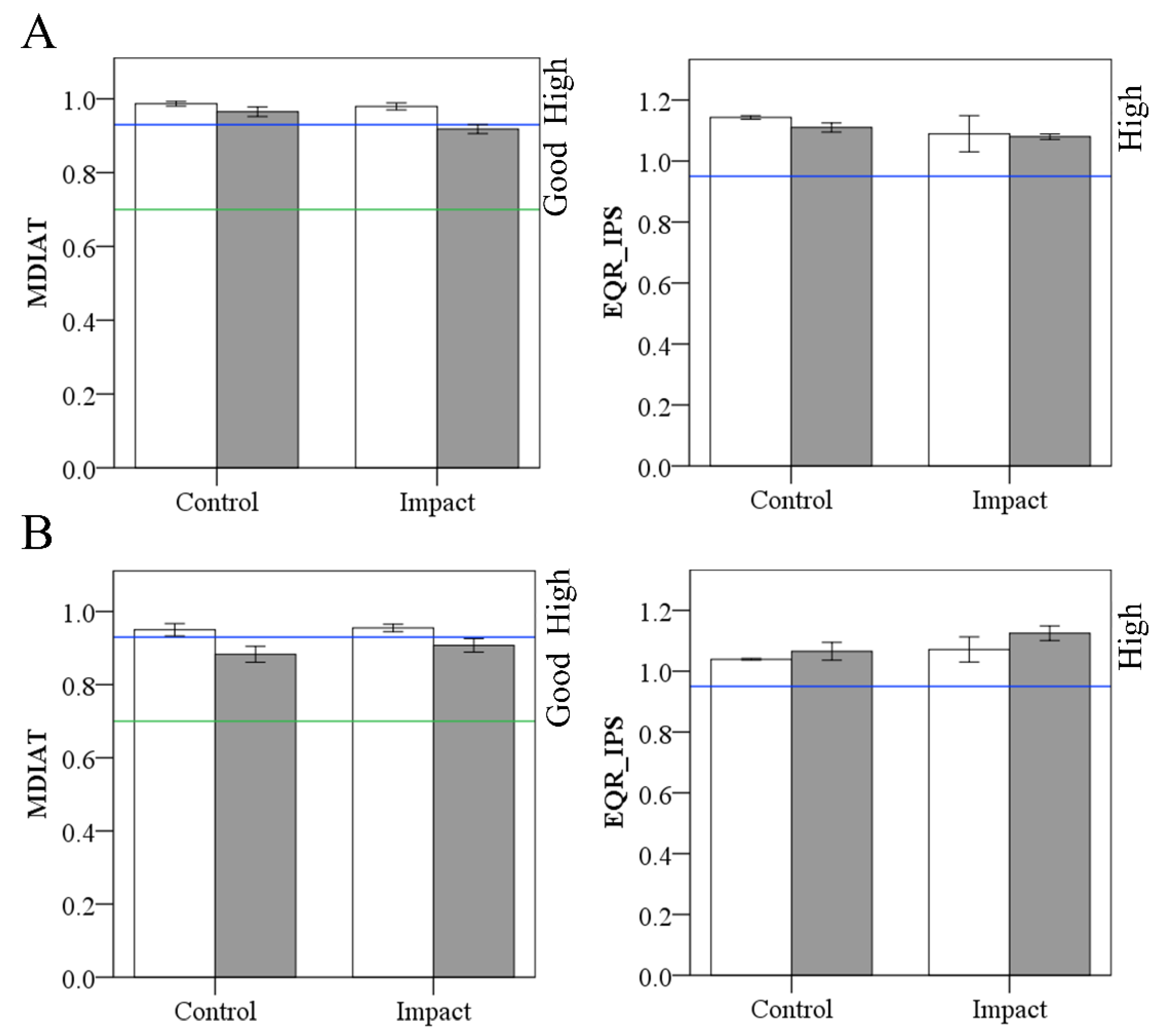
3.3.2. Ecological Status
3.4. Invertebrate Assemblages
3.4.1. Invertebrate Composition
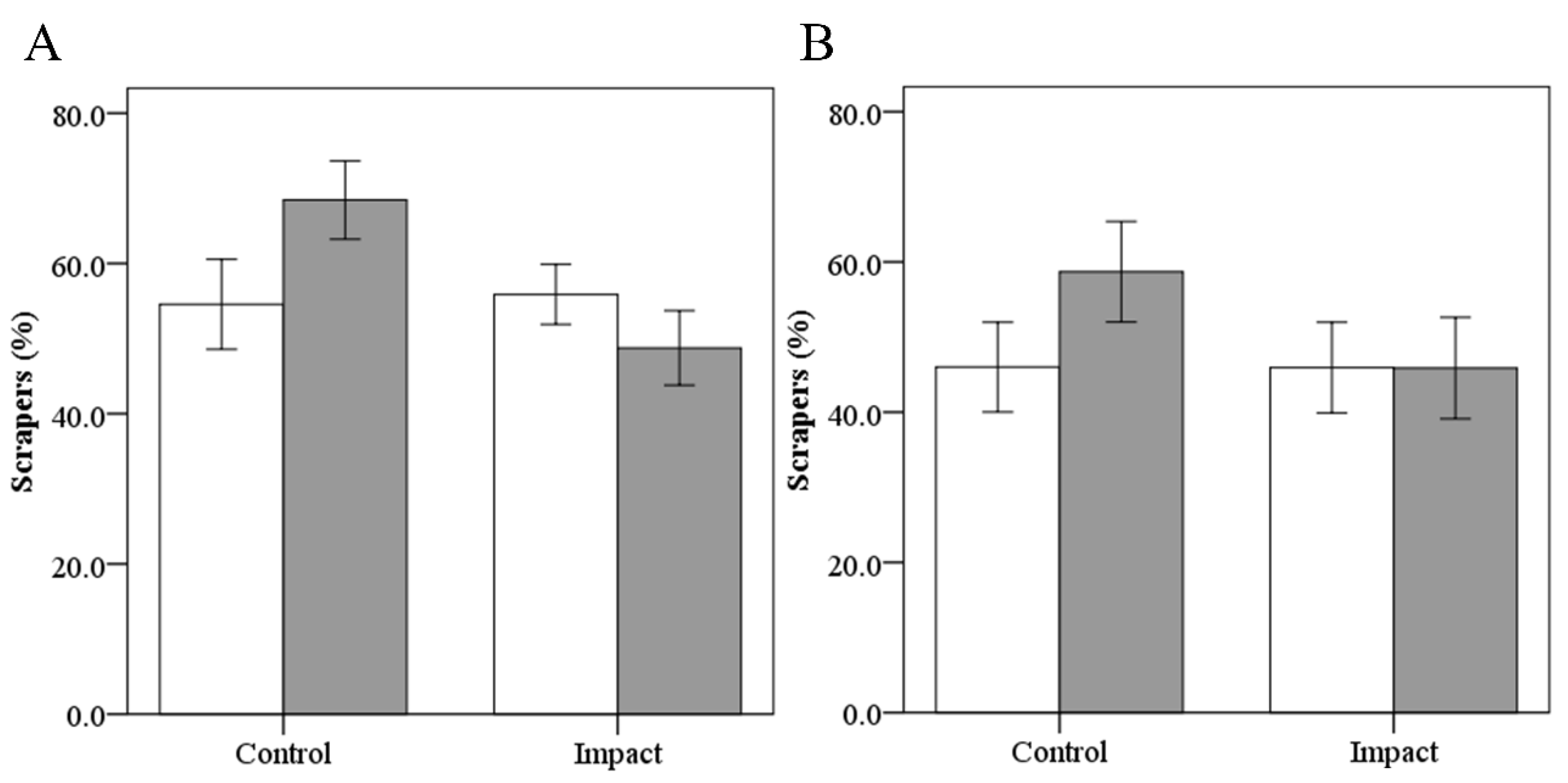
3.4.2. Scraper’s Abundance
4. Discussion
4.1. Eucalypt Influence on Physical-Chemical Variables
4.2. Eucalypt Effect on Biomass Accrual (Chl-a)
4.3. Eucalypt Leachates Influencing Diatom Species
4.4. Eucalypt Effect on Benthic Invertebrates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hagen, E.M.; Mctammany, M.E.; Webster, J.R.; Benfield, E.F. Shifts in allochthonous input and autochthonous production in streams along an agricultural land-use gradient. Hydrobiologia 2010, 655, 61–77. [Google Scholar] [CrossRef]
- Cordone, G.; Salinas, V.; Marina, T.I.; Doyle, S.R.; Pasotti, F.; Saravia, L.A.; Morno, F.R. Green vs brown food web: Effects of habitat type on multidimensional stability proxies for a highly-resolved Antarctic food web. Food Webs 2020, 25, e00166. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, S.G.; Likens, G.E. Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism. Ecol. Monogr. 1973, 43, 421–439. [Google Scholar] [CrossRef]
- Rosas, K.G.; Colón-Gaud, C.; Ramírez, A. Trophic basis of production in tropical headwater streams, Puerto Rico: An assessment of the importance of allochthonous resources in fueling food webs. Hydrobiologia 2020, 847, 1961–1975. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, J.; Lu, Y.; Yang, T.; Lyu, S.; Shi, K.; Zhou, F.; Yu, G. Determining dominating control mechanisms of inland water carbon cycling processes and associated gross primary productivity on regional and global scales. Earth Sci. Rev. 2021, 213, 103497. [Google Scholar] [CrossRef]
- Díaz Villa, M.V.E.; Cristiano, P.M.; De Diego, M.S.; Rodríguez, S.A.; Efron, S.T.; Bucci, S.J.; Scholz, F.; Goldstein, G. Do selective logging and pine plantations in humid subtropical forests affect aboveground primary productivity as well as carbon and nutrients transfer to soil? For. Ecol. Manag. 2022, 503, 119736. [Google Scholar] [CrossRef]
- Pozo, J.; Basaguren, A.; Elósegui, A.; Molinero, J.; Fabre, E.; Chauvet, E. Afforestation with Eucalyptus globulus and leaf litter decomposition in streams of northern Spain. Hydrobiologia 1998, 373–374, 101–110. [Google Scholar] [CrossRef]
- García, L.; Pardo, I. On the way to overcome some ecological riddles of forested headwaters. Hydrobiologia 2012, 696, 123–136. [Google Scholar] [CrossRef]
- Nambiar, E.K.S. Plantation forests: Their scope and a perspective on plantation nutrition. In Nutrition on Plantation Forests; Bowen, G.D., Nambiar, E.K.S., Eds.; Academic Press: London, UK, 1984; pp. 1–15. [Google Scholar]
- Molinero, J.; Pozo, J. Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics of coarse particulate organic matter (CPOM) in a small stream. Hydrobiologia 2004, 528, 143–165. [Google Scholar] [CrossRef]
- Quartucci, F.; Schweier, J.; Jaeger, D. Environmental analysis of Eucalyptus timber production from short rotation forestry in Brazil. Int. J. For. Eng. 2015, 26, 225–239. [Google Scholar] [CrossRef]
- Zaiton, S.; Sheriza, M.R.; Ainishifaa, R.; Alfred, K.; Norfaryanti, K. Eucalyptus in Malaysia: Review on environmental impacts. J. Landsc. Ecol. 2020, 13, 79–94. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Pozo, J.; Canhoto, C.; Elosegi, A. Effects of eucalyptus plantations on detritus, decomposers, and detritivores in streams. Sci. World 2002, 2, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Boulet, A.; Prats, S.A.; Malvar, M.C.; González-Pelayo, O.; Coelho, C.O.; Ferreira, A.J.; Keizer, J.J. Surface and Subsurface Flow in Eucalyptus Plantations in North-Central Portugal. J. Hydrol. Hydromech. 2015, 63, 197. [Google Scholar] [CrossRef] [Green Version]
- Goded, S.; Ekroos, J.; Domínguez, J.; Azcárate, J.G.; Guitián, J.A.; Smith, H.G. Effects of eucalyptus plantations on avian and herb species richness and composition in North-West Spain. Glob. Ecol. Conserv. 2019, 19, e00690. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Fernandes, F.; Ferreira, M.T. Effects of forest management on physical habitats and fish assemblages in Iberian eucalypt streams. For. Ecol. Manag. 2016, 363, 1–10. [Google Scholar] [CrossRef]
- Simões, S.; Gonçalves, A.L.; Canhoto, J.M.; Gonçalves, G.; Canhoto, C. Eucalyptus spp. Leaf traits determine litter processing by fungi and invertebrates. Freshw. Biol. 2021, 66, 968–977. [Google Scholar] [CrossRef]
- Bañuelos, R.; Larranaga, S.; Elosegi, A.; Pozo, J. Effects of Eucalyptus Plantations on CPOM Dynamics in Headwater Streams: A Manipulative Approach. Archiv Für Hydrobiologie 2004, 159, 211–228. [Google Scholar] [CrossRef]
- Pozo, J. Leaf litter processing of alder and eucalyptus in the Agüera stream system (North Spain) I. Chemical changes. Arch. Hydrobiol. 1993, 127, 299–317. [Google Scholar] [CrossRef]
- López, E.; Pardo, I.; Felpeto, N. Seasonal differences in green leaf breakdown and nutrient content of deciduous and evergreen tree species and grass in a granitic headwater stream. Hydrobiologia 2001, 464, 51–61. [Google Scholar] [CrossRef]
- Pozo, J.; González, E.; Díez, J.R.; Molinero, J.; Elósegui, A. Inputs of particulate organic matter to streams with different riparian vegetation. J. N. Am. Benthol. Soc. 1997, 16, 602–611. [Google Scholar] [CrossRef]
- Reichert, J.M.; de Deus Junior, J.C.; Borges Junior, N.; Cavalcante, R.B.L. Experimental catchments in the pampa biome: Database on hydrology in grasslands and eucalyptus plantations in subtropical Brazil. Hydrol. Proc. 2021, 35, e14285. [Google Scholar] [CrossRef]
- Martello, F.; de Bello, F.; de Castro Morini, M.S.; Silva, R.R.; Souza-Campana, D.R.D.; Ribeiro, M.C.; Carmona, C.P. Homogenization and impoverishment of taxonomic and functional diversity of ants in eucalyptus plantations. Sci. Rep. 2018, 8, 3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmigid, H.M.; Morsi, M.M. Cytotoxic and Molecular Impacts of Allelopathic Effects of Leaf Residues of Eucalyptus globulus on Soybean (Glycine max). J. Genet. Eng. Biotechnol. 2017, 15, 297–302. [Google Scholar] [CrossRef]
- Nega, F.; Gudeta, T.B. Allelopathic effect of Eucalyptus globulus Labill. On seed germination and seedling growth of highland teff (Eragrostis tef (Zuccagni) Trotter)) and barley (Hordeum vulgare L.). J. Exp. Agric. Int. 2019, 30, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Morsi, M.M.; Abdelmigid, H.M. Allelopathic activity of ‘Eucalyptus globulus’ leaf aqueous extract on ‘Hordeum vulgare’ growth and cytogenetic behaviour. Aust. J. Crop. Sci. 2016, 11, 1551–1556. [Google Scholar] [CrossRef]
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the bioherbicide potential of Eucalyptus globulus Labill: Biochemistry and effects of its aqueous extract. PloS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [Green Version]
- Puig, C.G.; Gonçalves, R.F.; Valentao, P.; Andrade, P.B.; Reigosa, M.J.; Pedrol, N. The consistency between phytotoxic effects and the dynamics of allelochemicals release from Eucalyptus globulus leaves used as biherbicide green manure. J. Chem. Ecol. 2018, 44, 658–670. [Google Scholar] [CrossRef]
- Souto, X.C.; Bolaño, J.C.; González, L.; Reigosa, M.J. Allelopathic effects of tree species on some soil microbial populations and herbaceous plants. Biol. Plan. 2001, 44, 269–275. [Google Scholar] [CrossRef]
- Puig, C.G.; Revilla, P.; Barreal, M.E.; Reigosa, M.J.; Pedrol, N. On the suitability of Eucalyptus globulus green manure for field weed control. Crop. Protection 2019, 121, 57–65. [Google Scholar] [CrossRef]
- European Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy (Water Framework Directive). Off. J. Eur. Union 2000, 43, 1–73. Available online: https://eur-lex.europa.eu/eli/dir/2000/60/oj (accessed on 4 October 2021).
- Palviainen, M.; Finér, L.; Laurén, A.; Mattsson, T.; Högbom, L. A method to estimate the impact of clear-cutting on nutrient concentrations in boreal headwater streams. Ambio 2015, 44, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marttila, H.; Kløve, B. Dynamics of erosion and suspended sediment transport from drained peatland forestry. J. Hydrol. 2010, 388, 414–425. [Google Scholar] [CrossRef]
- Kukuła, K.; Bylak, A. Synergistic impacts of sediment generation and hydrotechnical structures related to forestry on stream fish communities. Sci. Total Environ. 2020, 737, 139751. [Google Scholar] [CrossRef]
- González-Paz, L.; Delgado, C.; Pardo, I. Understanding divergences between ecological status classification systems based on diatoms. Sci. Total Environ. 2020, 734, 139418. [Google Scholar] [CrossRef]
- Debenest, T.; Silvestre, J.; Coste, M.; Delmas, F.; Pinelli, E. Herbicide Effects on Freshwater Benthic Diatoms: Induction of Nucleus Alterations and Silica Cell Wall Abnormalities. Aquat. Toxicol. 2008, 88, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.J.; Mitrovic, S.M.; Lim, R.P.; Warne, M.S.J.; Dunlop, J.; Kefford, B.J. Benthic diatoms as indicators of herbicide toxicity in rivers–A new SPEcies at risk (SPEARherbicides) index. Ecol. Indic. 2019, 99, 203–213. [Google Scholar] [CrossRef]
- Friberg, N.; Winterbourn, M.J. Interactions between riparian leaves and algal/microbial activity in streams. Hydrobiologia 1996, 341, 51–56. [Google Scholar] [CrossRef]
- Pinheiro, C.; Oliveira, U.; Rodrigues, T.; Oliva-Teles, L.; Vieira, M.N. Assessing the ecological status of a Mediterranean river: Benthic invertebrates and diatoms as complementary bioindicators. Limnetica 2020, 39, 299–315. [Google Scholar] [CrossRef]
- Canhoto, C.; Laranjeira, C. Leachates of Eucalyptus globulus in Intermittent Streams Affect Water Parameters and Invertebrates. Int. Rev. Hydrobiol. 2007, 92, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Andriotti, J.; Petrucio, M.M.; Silva, A.L.L.D. Exploring the impacts of non-native leaf litter on invertebrate community and leaf decomposition in a Atlantic Forest stream. Acta Limnol. Bras. 2022, 34, 1–11. [Google Scholar] [CrossRef]
- Eimil-Fraga, C.; Álvarez-Rodríguez, E.; Rodríguez-Soalleiro, R.; Fernández-Sanjurjo, M.J. Influence of Parent Material on the Aluminium Fractions in Acidic Soils Under Pinus pinaster in Galicia (NW Spain). Geoderma 2015, 255–256, 50–57. [Google Scholar] [CrossRef]
- Pardo, I. Comparative water quality characterization by PCA of an unperturbed and a polluted stream. Arch. FÜR Hydrobiol. 1994, 132, 95–114. [Google Scholar] [CrossRef]
- Delgado, C.; Pardo, I. Comparison of Benthic Diatoms from Mediterranean and Atlantic Spanish Streams: Community Changes in Relation to Environmental Factors. Aquat. Bot. 2015, 120, 304–314. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products Against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- González-Paz, L.; Almeida, S.F.P.; Pardo, I.; Delgado, C. Periphyton colonization and changes in the diatom assemblages of an artificial urban pond. Fundam. Appl. Limnol. 2020, 193, 313–326. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water, Sewage, and Wastewater, 17th ed.; American Public Health Association: New York, NY, USA, 1989. [Google Scholar]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations 1. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- BS EN 14407:2004; Water Quality. Guidance Standard for the Identification, and Enumeration of Benthic Diatom Samples from Rivers, and Their Interpretation. British Standards Institution: London, UK, 2004.
- Kelly, M.G.; Cazaubon, A.; Coring, E.; Dell’Uomo, A.; Ector, L.; Goldsmith, B.; Guasch, H.; Hürlimann, J.; Jarlman, A.; Kawecka, B.; et al. Recommendations for the routine sampling of diatoms for water quality assessments in Europe. J. Appl. Phycol. 1998, 10, 215–224. [Google Scholar] [CrossRef]
- AFNOR. Qualité de L’eau—Guide Pour L’échantillonnage en Routine et le Prétraitement des Diatomées Benthiques de Rivières. Norme Française NF EN 13946: 2003. Available online: www.boutique.afnor.org/fr-fr/norme/nf-en-13946/qualite-de-leau-guide-pour-lechantillonnage-en-routine-et-le-pretraitement-/fa102567/21845#AreasStoreProductsSummaryView (accessed on 10 June 2022).
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae, Süsswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1988; Volume 2–2, pp. 1–596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. Süsswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1991; Volume 2–3, pp. 1–600. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. Gesamtliteraturverzeichnis, Susswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1991; Volume 2–4, pp. 1–437. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 1. Allgemeiner und Encyonema part. Bibl. Diatomol. 1997, 36, 1–382. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 2. Encyonema part, Encyonopsis und Cymbellopsis. Bibl. Diatomol. 1997, 37, 1–469. [Google Scholar]
- Krammer, K. Cymbella. In Diatoms of the European Inlands Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Koeltz Botanical Books: Königstein, Germany, 2002; Volume 3, p. 584. [Google Scholar]
- Lange-Bertalot, H. 85 neue Taxa und über 100 weitere neu definierte Taxa ergänzend zur Süsswasserflora von Mitteleuropa 2/1–4. Biblioth. Diatomol. 1993, 27, 1–454. [Google Scholar]
- Lange-Bertalot, H. (Ed.) Navicula sensu stricto. 10 genera separated from Navicula sensu lato. Frustulia. In Diatoms of Europe 2; Verlag: Stuttgart, Germany, 2001; p. 526. [Google Scholar]
- Prygiel, J.; Coste, M. Guide Méthodologique Pour la Mise en Œuvre de L’INDICE Biologique Diatomées. NF T90-394Ï; Agence de L’Eau Artois-Picardie: Douai, France, 2000; p. 134. [Google Scholar]
- Trobajo, R.; Rovira, L.; Ector, L.; Wetzel, C.E.; Kelly, M.; Mann, D.G. Morphology and identity of some ecologically important small Nitzschia species. Diatom Res. 2013, 28, 37–59. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Ector, L.; Van der Vijver, B.; Compère, P.; Mann, D.G. Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea 2015, 15, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Death, R.G.; Winterbourn, M.J. Environmental Stability and Community Persistence: A Multivariate Perspective. J. N. Am. Benthol. Soc. 1994, 13, 125–139. [Google Scholar] [CrossRef]
- Matthaei, C.D.; Arbuckle, C.J.; Townsend, C.R. Stable surface stones as refugia for invertebrates during disturbance in a New Zealand stream. J. N. Am. Benthol. Soc. 2000, 19, 82–93. [Google Scholar] [CrossRef]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’Eau Douce, 3rd ed.; CNRS: Paris, France, 2010; p. 607. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design Gleneden Beach: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-Based Redundancy Analysis: Testing Multispecies Responses in Multifactorial Ecological Experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.; Clarke, K.P. For PRIMER: Guide to Software and Statistical Methods; Primer-e: Plymouth, UK, 2008. [Google Scholar]
- Akaike, H. Factor Analysis and the AIC. Psychometrika 1987, 52, 317–332. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER V7. PRIMER-E: Plymouth; Plymouth Marine Laboratory: Plymouth, UK, 2015; Volume 20. [Google Scholar]
- Cemagref, M. Etude Des Méthodes Biologiques D’Appréciation Quantitative De La Qualité Des Eaux. In Rapport Cemagref QE Lyon-AF; Bassin Rhône Méditerranée Corse: Lyon, France, 1982. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. Omnidia 3.2. Diatom Index Software Including Diatom Database with Taxonomic Names, References and Codes of 11645 Diatom Taxa. Hydrobiologia 2003, 269, 509–513. [Google Scholar]
- Delgado, C.; Pardo, I.; García, L. A multimetric diatom index to assess the ecological status of coastal Galician rivers (NW Spain). Hydrobiologia 2010, 644, 371–384. [Google Scholar] [CrossRef]
- Kelly, M.G.; Gómez-Rodríguez, C.; Kahlert, M.; Almeida, S.F.; Bennett, C.; Bottin, M.; Delmas, F.; Descy, J.P.; Dörflinger, G.; Kennedy, B.; et al. Establishing expectations for pan-european diatom based ecological status assessments. Ecol. Indic. 2012, 20, 177–186. [Google Scholar] [CrossRef]
- Ferreira, V.; Larrañaga, A.; Gulis, V.; Basaguren, A.; Elosegi, A.; Graça, M.A.S.; Pozo, J. The Effects of Eucalypt Plantations on Plant Litter Decomposition and Macroinvertebrate Communities in Iberian Streams. For. Ecol. Manage. 2015, 335, 129–138. [Google Scholar] [CrossRef]
- Dodds, W.K.; Jones, J.R.; Welch, E.B. Suggested Classification of Stream Trophic State: Distributions of Temperate Stream Types by Chlorophyll, Total Nitrogen, and Phosphorus. Water Res. 1998, 32, 1455–1462. [Google Scholar] [CrossRef]
- Drummond, J.D.; Bernal, S.; von Schiller, D.; Martí, E. Linking in-Stream Nutrient Uptake to Hydrologic Retention in Two Headwater Streams. Freshw. Sci. 2016, 35, 1176–1188. [Google Scholar] [CrossRef]
- Hoellein, T.J.; Tank, J.L.; Rosi-Marshall, E.J.; Entrekin, S.A.; Lamberti, G.A. Controls on spatial and temporal variation of nutrient uptake in three Michigan headwater streams. Limnol. Oceanogr. 2007, 52, 1964–1977. [Google Scholar] [CrossRef]
- Pardo, I.; Kuglerová, L.; García, L.; Martí, E. Nutrient availability modulates the effect of water abstraction on the metabolism of 2 lowland forested streams. Freshw. Sci. 2022, 41, 299–314. [Google Scholar] [CrossRef]
- Grimm, N.B.; Fisher, S.G.; Minckley, W.L. Nitrogen and phosphorus dynamics in hot desert streams of southwestern USA. Hydrobiologia 1981, 83, 303–312. [Google Scholar] [CrossRef]
- Hill, W.R.; Mulholland, P.J.; Marzolf, E.R. Stream ecosystem responses to forest leaf emergence in spring. Ecology 2001, 82, 2306–2319. [Google Scholar] [CrossRef]
- Pardo, I.; Álvarez, M. Comparison of resource and consumer dynamics in Atlantic and Mediterranean streams. Limnetica 2006, 25, 271–286. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Yu, J.Q. Allelochemicals and photosynthesis. In Allelopathy: A physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 127–156. [Google Scholar]
- Jayakumar, M.; Eyini, M. Allelopathic effect of Eucalyptus globulus labil in groundnut and corn. Comp. Physiol. Ecol. 1990, 15, 109–113. [Google Scholar]
- Zhao, W.; Zheng, Z.; Zhang, J.; Roger, S.; Luo, X. Evaluation of the use of eucalyptus to control algae bloom and improve water quality. Sci. Total Environ. 2019, 667, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Sillett, S.C.; Van Pelt, R.; Kramer, R.D.; Carroll, A.L.; Koch, G.W. Biomass and growth potential of Eucalyptus regnans up to 100m tall. For. Ecol. Manage. 2015, 348, 78–91. [Google Scholar] [CrossRef]
- Vallotton, N.; Moser, D.; Eggen, R.I.L.; Junghans, M.; Chèvre, N. S-metolachlor pulse exposure on the alga Scenedesmus vacuolatus: Effects during exposure and the subsequent recovery. Chemosphere 2008, 73, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Delgado, C.; Abraín, R.; Gómez-Rodríguez, C.; García-Roselló, E.; García, L.; Reynoldson, T.B. A predictive diatom-based model to assess the ecological status of streams and rivers of Northern Spain. Ecol. Indic. 2018, 90, 519–528. [Google Scholar] [CrossRef]
- Paule, A.; Roubeix, V.; Lauga, B.; Duran, R.; Delmas, F.; Paul, E.; Rols, J.L. Changes in tolerance to herbicide toxicity throughout development stages of phototrophic biofilms. Aquat. Toxicol. 2013, 144–145, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Esteves, S.M.; Keck, F.; Almeida, S.F.P.; Figueira, E.; Bouchez, A.; Rimet, F. Can we Predict Diatoms Herbicide Sensitivities with Phylogeny? Influence of Intraspecific and Interspecific Variability. Ecotoxicology 2017, 26, 1065–1077. [Google Scholar] [CrossRef]
- Turunen, J.; Markkula, J.; Rajakallio, M.; Aroviita, J. Riparian forests mitigate harmful ecological effects of agricultural diffuse pollution in medium-sized streams. Sci. Total Environ. 2019, 649, 495–503. [Google Scholar] [CrossRef]
- Genkal, S.I.; Yarushina, M.I. To morphology, taxonomy and distribution of rare species of the genus Sellaphora (Bacillariophyta) in Russia. Int. J. Algae 2018, 20, 1–12. [Google Scholar] [CrossRef]
- Cantonati, M.; Zorza, R.; Bertoli, M.; Pastorino, P.; Salvi, G.; Platania, G.; Prearo, M.; Pizzul, E. Recent and Subfossil Diatom Assemblages as Indicators of Environmental Change (Including Fish Introduction) in a High-Mountain Lake. Ecol. Ind. 2021, 125, 107603. [Google Scholar] [CrossRef]
- Kazanci, N.; Ekingen, P.; Türkmen, G.; Ertunç, Ö.; Dügel, M.; Gültutan, Y. Assessment of ecological quality of Aksu Stream (Giresun, Turkey) in Eastern Black Sea Region by using Water Framework Directive (WFD) methods based on benthic macroinvertebrates. Rev. Hydrobiol. 2010, 3, 165–184. [Google Scholar]
- Farhadian, O.; Pouladi, M.; Vazirizadeh, A.; Sedaghat, R. A Study of Diatoms Seasonal Distribution and Biodiversity in Helleh River Estuary, Persian Gulf. Environ. Stud. Persian Gulf 2015, 2, 32–44. [Google Scholar]
- Andersen, D.K.; Larsen, S.E.; Johansson, L.S.; Alnøe, A.B.; Baattrup-Pedersen, A. Udvikling Af Biologisk Indeks for Bentiske Alger (Fytobenthos) i Danske Vandløb; Aarhus Universitet, DCE–Nationalt Center for Miljø og Energi: Roskilde, Denmark, 2018; Volume 42. [Google Scholar]
- González-Paz, L.; Delgado, C.; Pardo, I. How good is good ecological status? A test across river typologies, diatom indices and biological elements. Sci. Total Environ. 2022, 815, 152901. [Google Scholar] [CrossRef] [PubMed]
- García, L.; Pardo, I.; Richardson, J. A cross-continental comparison of stream invertebrate community assembly to assess convergence in forested headwater streams. Aquat. Sci. 2014, 76, 29–40. [Google Scholar] [CrossRef]
- Wasserman, R.J.; Sanga, S.; Buxton, M.; Dalu, T.; Cuthbert, R.N. Does invasive river red gum (Eucalyptus camaldulensis) alter leaf litter decomposition dynamics in arid zone temporary rivers? Inland Waters 2020, 11, 104–113. [Google Scholar] [CrossRef]
- Larrañaga, A.; Basaguren, A.; Pozo, J. Impacts of Eucalyptus globulus plantations on physiology and population densities of invertebrates inhabiting Iberian Atlantic streams. Int. Rev. Hydrobiol. 2009, 94, 497–511. [Google Scholar] [CrossRef]
- García, L.; Pardo, I. Food type and temperature constraints on the fitness of a dominant freshwater shredder. Ann. De Limnol. 2015, 51, 227–235. [Google Scholar] [CrossRef] [Green Version]
- García, L.; Richardson, J.S.; Pardo, I. Leaf quality influences invertebrate colonization and drift in a temperate rainforest stream. Can. J. Fish. Aquat. Sci. 2012, 69, 1663–1673. [Google Scholar] [CrossRef]
- Ledger, M.E.; Hildrew, A.G. The ecology of acidification and recovery: Changes in herbivore-algal food web linkages across a stream pH gradient. Environ. Pollut. 2005, 137, 103–118. [Google Scholar] [CrossRef]
- Kidd, K.A.; Paterson, M.J.; Rennie, M.D.; Podemski, C.L.; Findlay, D.L.; Blanchfield, P.J.; Liber, K. Direct and indirect responses of a freshwater food web to a potent synthetic oestrogen. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130578. [Google Scholar] [CrossRef] [Green Version]
- Dodds, W.K.; Smith, V.H. Nitrogen, Phosphorus, and Eutrophication in Streams. Inland Waters 2016, 6, 155–164. [Google Scholar] [CrossRef]
- Pandey, L.K.; Bergey, E.A. Metal toxicity and recovery response of riverine periphytic algae. Sci. Total Environ. 2018, 642, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Karaouzas, I.; Smeti, E.; Vourka, A.; Vardakas, L.; Mentzafou, A.; Tornés, E.; Sabater, S.; Muñoz, I.; Skoulikidis, N.T.; Kalogianni, E. Assessing the ecological effects of water stress and pollution in a temporary river-Implications for water management. Sci. Total Environ. 2018, 618, 1591–1604. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| As Murteiras | Ventín | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Impact | Significance | Control | Impact | Significance | |||||||
| B | Ar | B | A | B/A | C/I | B | A | B | A | B/A | C/I | |
| Conductivity (µs cm−1) | 19.0 ± 0.2 | 19.2 ± 0.1 | 18.1 ± 0.1 | 19.4 ± 0.7 | * | 19.0 ± 0.2 | 19.0 ± 0.6 | 21.2 ± 1.1 | 21.9 ± 1.7 | * | ||
| pH | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.7 ± 0.3 | 6.4 ± 0.0 | * | |||
| Dissolved oxygen (mg L−1) | 8.9 ± 0.0 | 8.8 ± 0.1 | 9.2 ± 0.1 | 8.9 ± 0.0 | * | * | 8.8 ± 0.3 | 9.1 ± 0.1 | 8.6 ± 0.0 | 9.0 ± 0.2 | * | |
| Oxygen saturation (%) | 97.9 ± 1.5 | 96.7 ± 1.1 | 100.0 ± 0.6 | 97.0 ± 0.6 | * | * | 95.4 ± 1.2 | 97.8 ± 0.7 | 94.6 ± 0.9 | 96.3 ± 1.5 | * | |
| Water temperature (°C) | 17.6 ± 0.8 | 16.5 ± 0.1 | 17.0 ± 0.7 | 16.9 ± 0.7 | 17.6 ± 0.8 | 16.5 ± 1.3 | 18.1 ± 0.4 | 16.5 ± 0.8 | * | |||
| Water temperature (°C) 24 h | 16.1 ± 0.0 | 15.8 ± 0.0 | * | 15.6 ± 0.0 | 15.3 ± 0.0 | 15.5 ± 0.0 | 15.3 ± 0.0 | * | * | |||
| Luminosity (lum ft−2) | 8.7 ± 0.8 | 9.4 ± 0.8 | 5.6 ± 0.5 | 3.7 ± 0.3 | 10.1 ± 0.9 | 7.0 ± 0.6 | * | * | ||||
| N-DIN (µg L−1) | 10.3 ± 2.4 | 9.2 ± 2.6 | 9.3 ± 0.9 | 7.6 ± 0.2 | 9.5 ± 3.7 | 4.5 ± 1.9 | 20.6 ± 1.2 | 15.5 ± 3.4 | * | * | ||
| P-PO43− (µg L−1) | 6.3 ± 0.2 | 10.9 ± 5.6 | 5.5 ± 0.0 | 8.7 ± 3.3 | 5.7 ± 0.7 | 12.9 ± 6.4 | 5.3 ± 0.4 | 12.3 ± 7.5 | * | |||
| Discharge (m3 s−1) | 0.09 ± 0.06 | 0.11 ± 0.01 | 0.40 ± 0.16 | 0.48 ± 0.12 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Paz, L.; Gestido, J.; Delgado, C.; Pedrol, N.; Pardo, I. Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters. Water 2023, 15, 115. https://doi.org/10.3390/w15010115
González-Paz L, Gestido J, Delgado C, Pedrol N, Pardo I. Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters. Water. 2023; 15(1):115. https://doi.org/10.3390/w15010115
Chicago/Turabian StyleGonzález-Paz, Lorena, Josué Gestido, Cristina Delgado, Nuria Pedrol, and Isabel Pardo. 2023. "Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters" Water 15, no. 1: 115. https://doi.org/10.3390/w15010115
APA StyleGonzález-Paz, L., Gestido, J., Delgado, C., Pedrol, N., & Pardo, I. (2023). Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters. Water, 15(1), 115. https://doi.org/10.3390/w15010115