
Quantitative Stable Isotope Tracing of Water Uptake in Cynanchum sibiricum during Its Growth Period in Arid Areas of Northwest China

Abstract
:1. Introduction
2. Materials and Methods
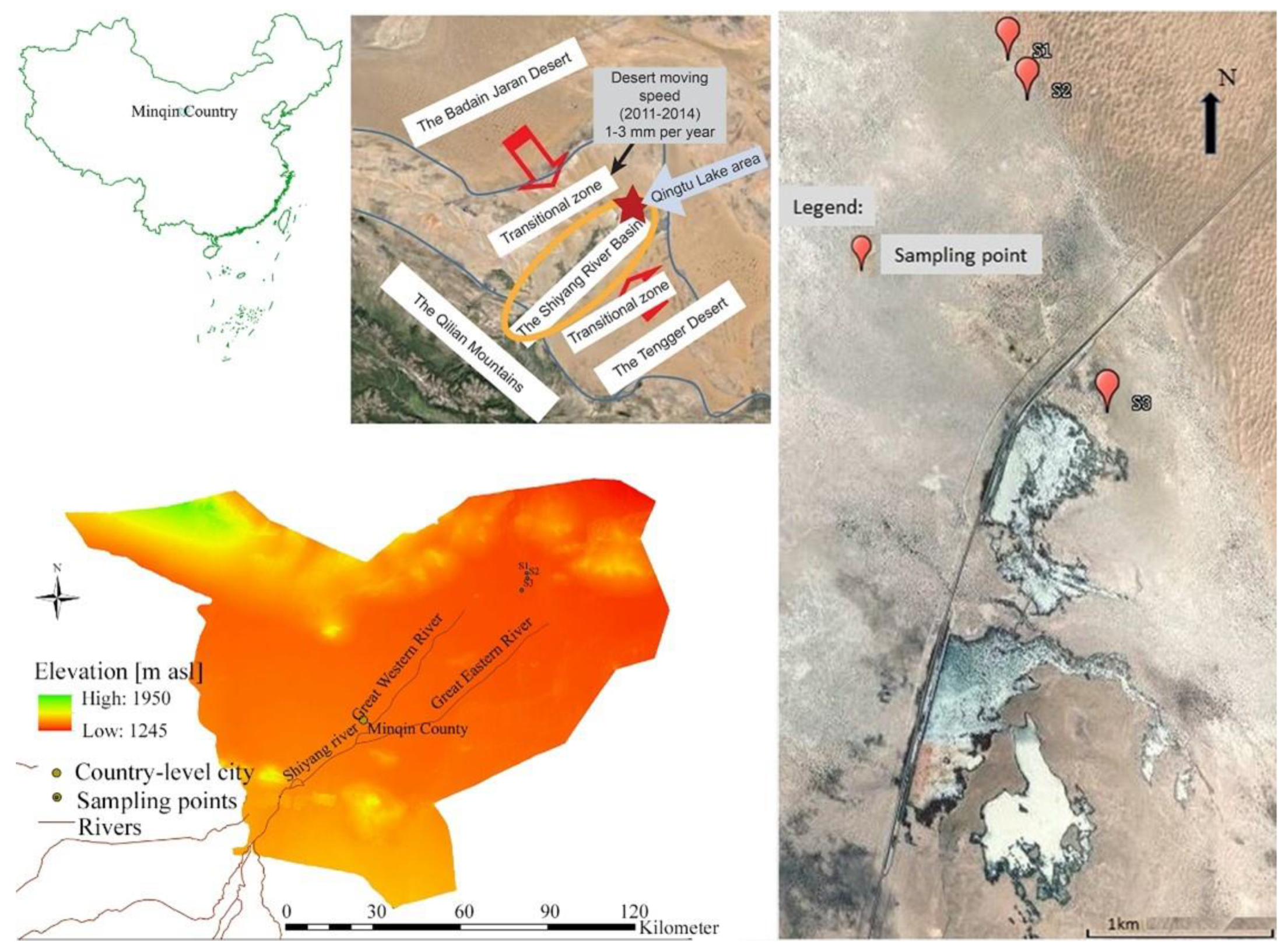
2.1. Study Area
2.2. Collection and Determination of Samples
2.3. Multivariate Linear Mixed Model Method
3. Results and Discussions
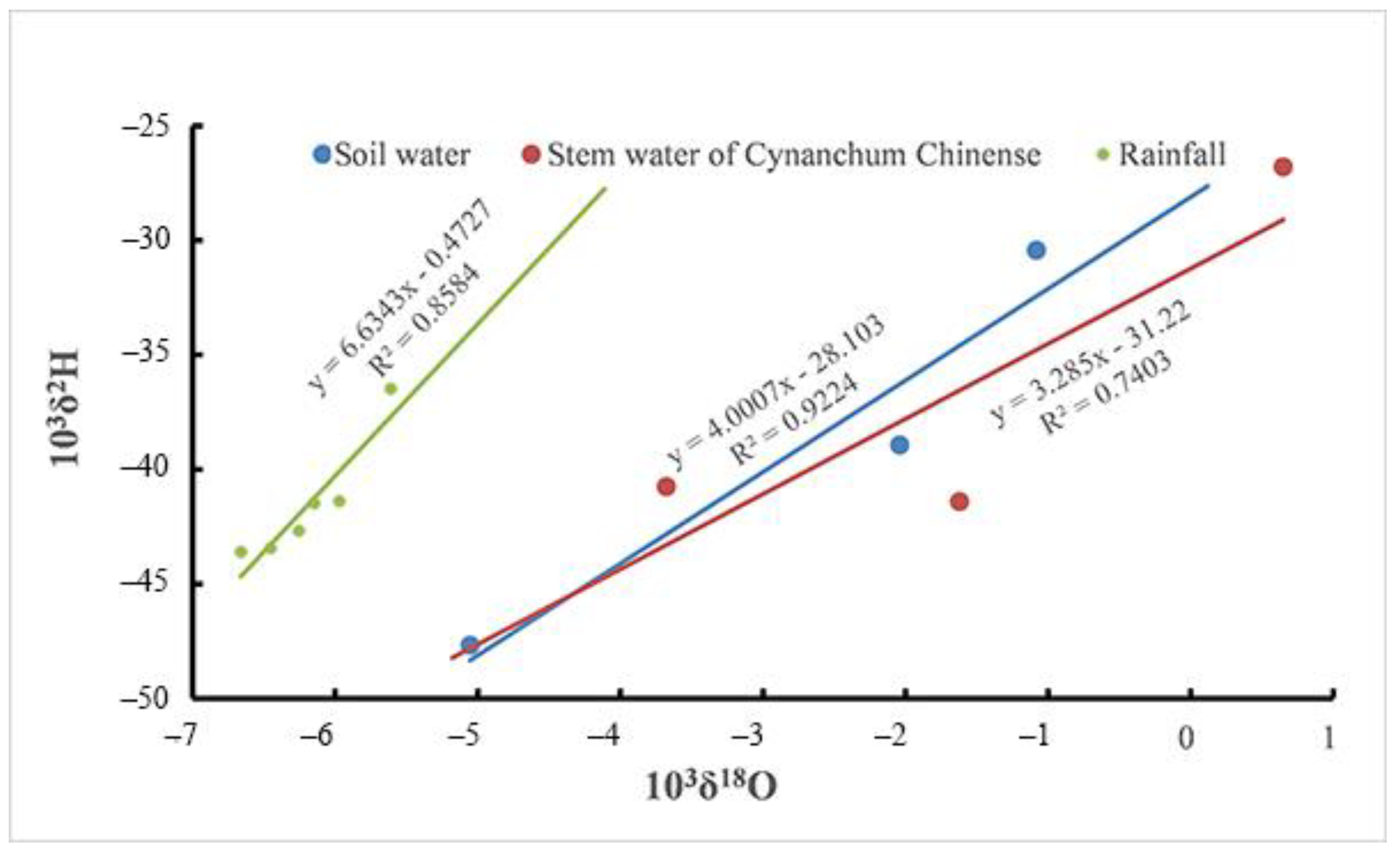
3.1. The Relationships between δ18O and δD in Rainfall, Soil Water, and the Xylem Water of Cynanchum sibiricum
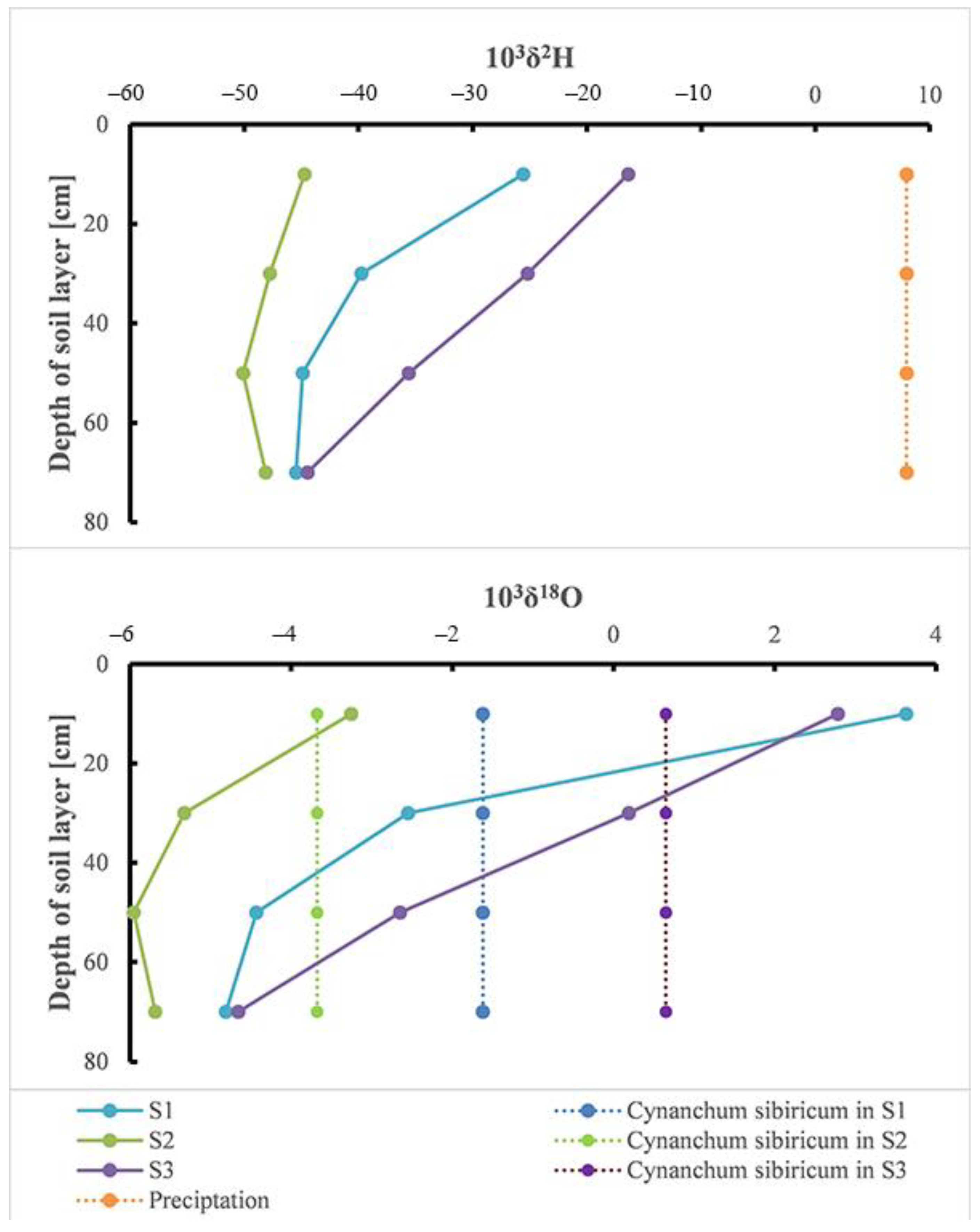
3.2. Vertical Distribution of δ18O and δD in Soil Moisture
3.3. The Water Sources and Contributions of Cynanchum sibiricum in the Growth Period
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beyer, M.; Koeniger, P.; Gaj, M.; Hamutoko, J.T.; Wanke, H.; Himmelsbach, T. A Deuterium-based labeling technique for the investigation of rooting depths, water uptake dynamics and unsaturated zone water transport in semiarid environments. J. Hydrol. 2016, 533, 627–643. [Google Scholar] [CrossRef]
- Youri, R.; Mathieu, J. Reviews and syntheses: Isotopic approaches to quantify root water uptake: A review and comparison of methods. Biogeosciences 2017, 14, 2199–2224. [Google Scholar]
- Germon, A.; Laclau, J.P.; Robin, A.; Jourdan, C. Tamm Review: Deep fine roots in forest ecosystems: Why dig deeper? For. Ecol. Manag. 2020, 466, 118135. [Google Scholar] [CrossRef]
- Eamus, D.; Zolfaghar, S.; Villalobos-Vega, R.; Cleverly, J.; Huete, A. Groundwater-dependent ecosystems: Recent insights, new techniques and an ecosystem-scale threshold response. Hydrol. Earth Syst. Sci. Discuss. 2015, 12, 4677–4754. [Google Scholar]
- Beyer, M.; Hamutoko, J.T.; Wanke, H.; Gaj, M.; Koeniger, P. Examination of deep root water uptake using anomalies of soil water stable isotopes, depth-controlled isotopic labeling and mixing models. J. Hydrol. 2018, 566, 122–136. [Google Scholar] [CrossRef]
- Barbeta, A.; Peñuelas, J. Relative contribution of groundwater to plant transpiration estimated with stable isotopes. Sci. Rep. 2017, 7, 10580. [Google Scholar] [CrossRef] [Green Version]
- Berry, Z.C.; Evaristo, J.; Moore, G.; Poca, M.; Steppe, K.; Verrot, L.; Asbjornsen, H.; Borma, L.S.; Bretfeld, M.; Hervé-Fernández, P.; et al. The two water worlds hypothesis: Addressing multiple working hypotheses and proposing a way forward. Ecohydrology 2017, 11, e1843. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.-L.; Clément, C.; Montoroi, J.-P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 4, mcw130. [Google Scholar] [CrossRef] [Green Version]
- Antunes, C.; Barradas, M.C.D.; Zunzunegui, M.; Vieira, S.; Pereira, Â.; Anjos, A.; Correia, O.; Pereira, M.J.; Máguas, C. Contrasting plant water-use responses to groundwater depth in coastal dune ecosystems. Funct. Ecol. 2018, 32, 1931–1943. [Google Scholar] [CrossRef]
- Jiang, S.X.; An, F.B.; Ma, J.P.; Zhao, P.; Liu, H.J.; Liu, S.J. Water sources of Nitraria tangutorum nebkhas and its response to ecological water transfer in Qingtu lake in lower reaches of Shiyang river. J. Arid Land Resour. Environ. 2019, 33, 176–182, (In Chinese with English Abctract). [Google Scholar]
- Zhou, T.H.; Zhao, C.Y.; Wu, G.L.; Jiang, S.W.; Yu, Y.X.; Wang, D.D. Application of stable isotopes in analyzing the water sources of Populus euphratica and tamarix ramosissima in the upstream of tarim river. J. Desert Res. 2017, 37, 124, (In Chinese with English Abctract). [Google Scholar]
- Li, Y.F.; Yu, J.J.; Lu, K.; Wang, P.; Zhang, Y.C.; Du, C.Y. Water sources of Populus euphratica and Tamarix ramosissima in Ejina Delta, the lower reaches of the Heihe River, China. Chin. J. Plant Ecol. 2017, 41, 519–528, (In Chinese with English Abctract). [Google Scholar]
- Debandi, G.; Rossi, B.E.; Villagra, P.E.; Giantomasi, M.A.; Mantován, N.G. Spatial and temporal synchronicity in the phenological events of Prosopis flexuosa in the Central Monte Desert. Rev. Fac. Cienc. Agrar. 2020, 52, 148–160. [Google Scholar]
- Phillips, D.L.; Gregg, J.W. Source partitioning using stable isotopes: Coping with too many sources. Oecologia 2003, 136, 261–269. [Google Scholar] [CrossRef]
- Duan, D.Y.; Ouyang, H. Application of stable hydrogen and oxygen isotope in analyzing plant water use sources. Ecol. Environ. 2007, 16, 655–660. [Google Scholar]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Xing, D.; Xiao, J.J.; Han, S.Y.; Peng, G.H.; Fu, W.T.; Jia, Y.L. Water absorption source analysis of mulberry roots based on stable isotopes in rocky desertification area. Trans. Chin. Soc. Agric. Eng. 2019, 35, 77–84, (In Chinese with English Abctract). [Google Scholar]
- Liu, J.R.; Song, X.F.; Yuan, G.F.; Sun, X.M.; Yang, L.H. Stable isotopic compositions of precipitation in China. Tellus B 2014, 66, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Chen, H.; Zhu, J.; Chen, T. Water sources of five dominant desert species in Nuomuhong area of Qaidam Basin. Acta Ecol. Sin. 2014, 34, 6277–6286, (In Chinese with English Abctract). [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Chen, Y.P. Research progress in water uptake models by plant roots. Acta Prataculturae Sin. 2017, 26, 214–225. [Google Scholar]
- Couvreur, V.; Rothfuss, Y.; Meunier, F.; Bariac, T.; Biron, P.; Durand, J.-L.; Richard, P.; Javaux, M. Disentangling temporal and population variability in plant root water uptake from stable isotopic analysis: When rooting depth matters in labeling studies. Hydrol. Earth Syst. Sci. 2020, 24, 3057–3075. [Google Scholar] [CrossRef]
- Arndt, P.; Maren, D.; Rolf, S.; Matthias, C.; Christiane, W. Quantification of dynamic soil–vegetation feedbacks following an isotopically labelled precipitation pulse. Biogeosciences 2017, 14, 2293–2306. [Google Scholar]
- Ceperley, N.; Mande, T.; Rinaldo, A.; Parlange, M.B. Evidence of hydraulic lift for pre-rainy season leaf out and dry-season stem water enrichment in Sclerocarya birrea, a tropical agroforestry tree. EGU Gen. Assem. Conf. Abstr. 2014, 16, 8261. [Google Scholar]
- Yuan, G.F.; Zhang, P.; Xue, S.S.; Zhuang, W. Change characteristics in soil water content in root zone and evidence of root hydraulic lift in Tamarix ramosissima thickets on sand dunes. Chin. J. Plant Ecol. 2012, 36, 1033–1042, (In Chinese with English Abctract). [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rainfall | 0–10 cm Soil Water | 10–30 cm Soil Water | 30–50 cm Soil Water | 50–70 cm Soil Water | |
|---|---|---|---|---|---|
| S1 | 0.14 ± 0.0010 | 0.33 ± 0.0005 | 0.21 ± 0.0015 | 0.16 ± 0.0011 | 0.16 ± 0.0011 |
| S2 | 0.03 ± 0.0010 | 0.83 ± 0.0004 | 0.05 ± 0.0015 | 0.04 ± 0.0011 | 0.04 ± 0.0013 |
| S3 | 0.06 ± 0.0008 | 0.56 ± 0.0017 | 0.21 ± 0.0026 | 0.10 ± 0.0013 | 0.07 ± 0.0009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Chen, Z.; Hao, J.; Wang, J.; Zhang, G. Quantitative Stable Isotope Tracing of Water Uptake in Cynanchum sibiricum during Its Growth Period in Arid Areas of Northwest China. Water 2022, 14, 1422. https://doi.org/10.3390/w14091422
Dong H, Chen Z, Hao J, Wang J, Zhang G. Quantitative Stable Isotope Tracing of Water Uptake in Cynanchum sibiricum during Its Growth Period in Arid Areas of Northwest China. Water. 2022; 14(9):1422. https://doi.org/10.3390/w14091422
Chicago/Turabian StyleDong, Haibiao, Zongyu Chen, Jing Hao, Jinzhe Wang, and Guanghui Zhang. 2022. "Quantitative Stable Isotope Tracing of Water Uptake in Cynanchum sibiricum during Its Growth Period in Arid Areas of Northwest China" Water 14, no. 9: 1422. https://doi.org/10.3390/w14091422
APA StyleDong, H., Chen, Z., Hao, J., Wang, J., & Zhang, G. (2022). Quantitative Stable Isotope Tracing of Water Uptake in Cynanchum sibiricum during Its Growth Period in Arid Areas of Northwest China. Water, 14(9), 1422. https://doi.org/10.3390/w14091422