
Diversity and Vertical Distribution of Sedimentary Bacterial Communities and Its Association with Metal Bioavailability in Three Distinct Mangrove Reserves of South China

Abstract
:1. Introduction
2. Materials and Methods
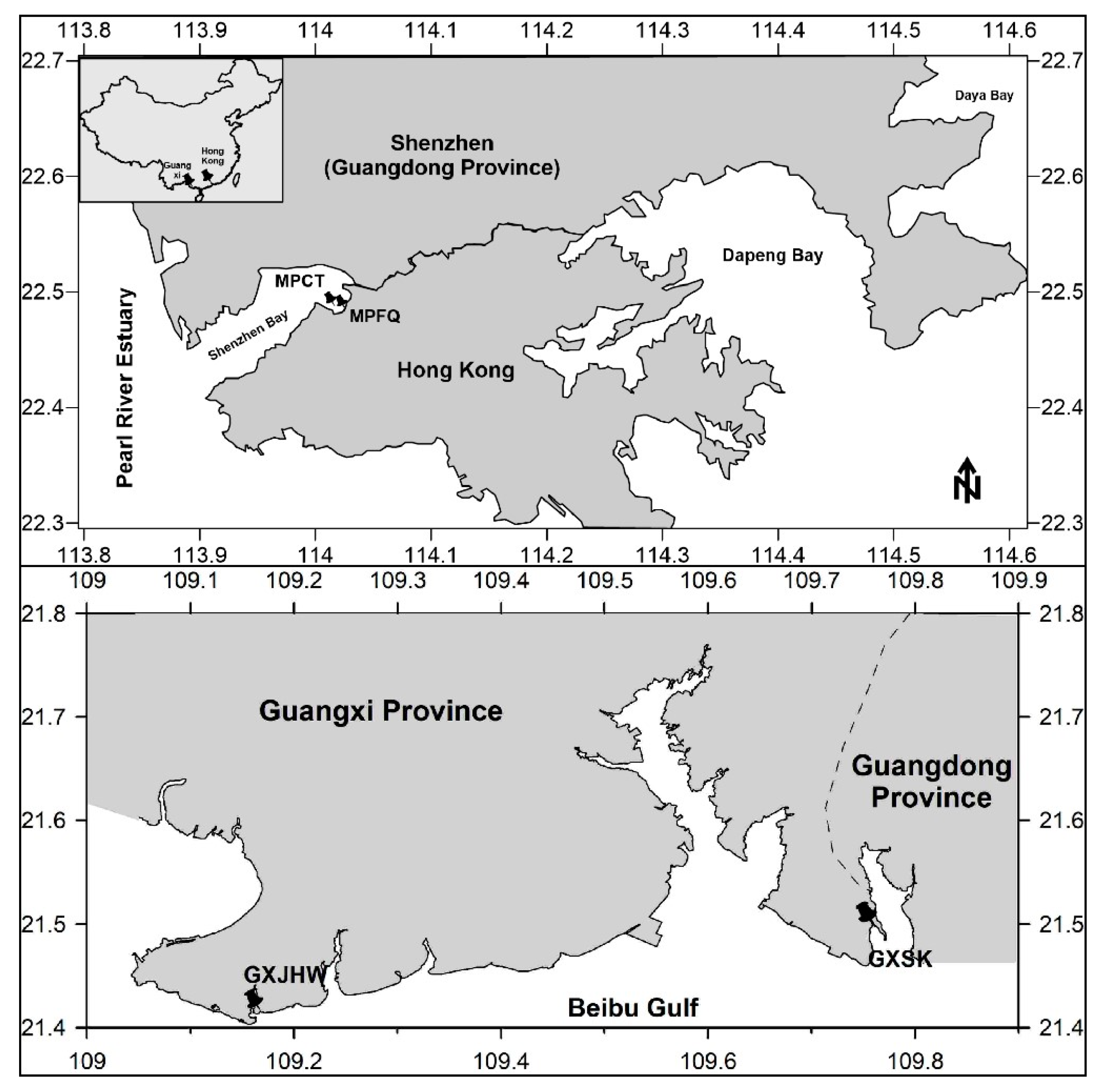
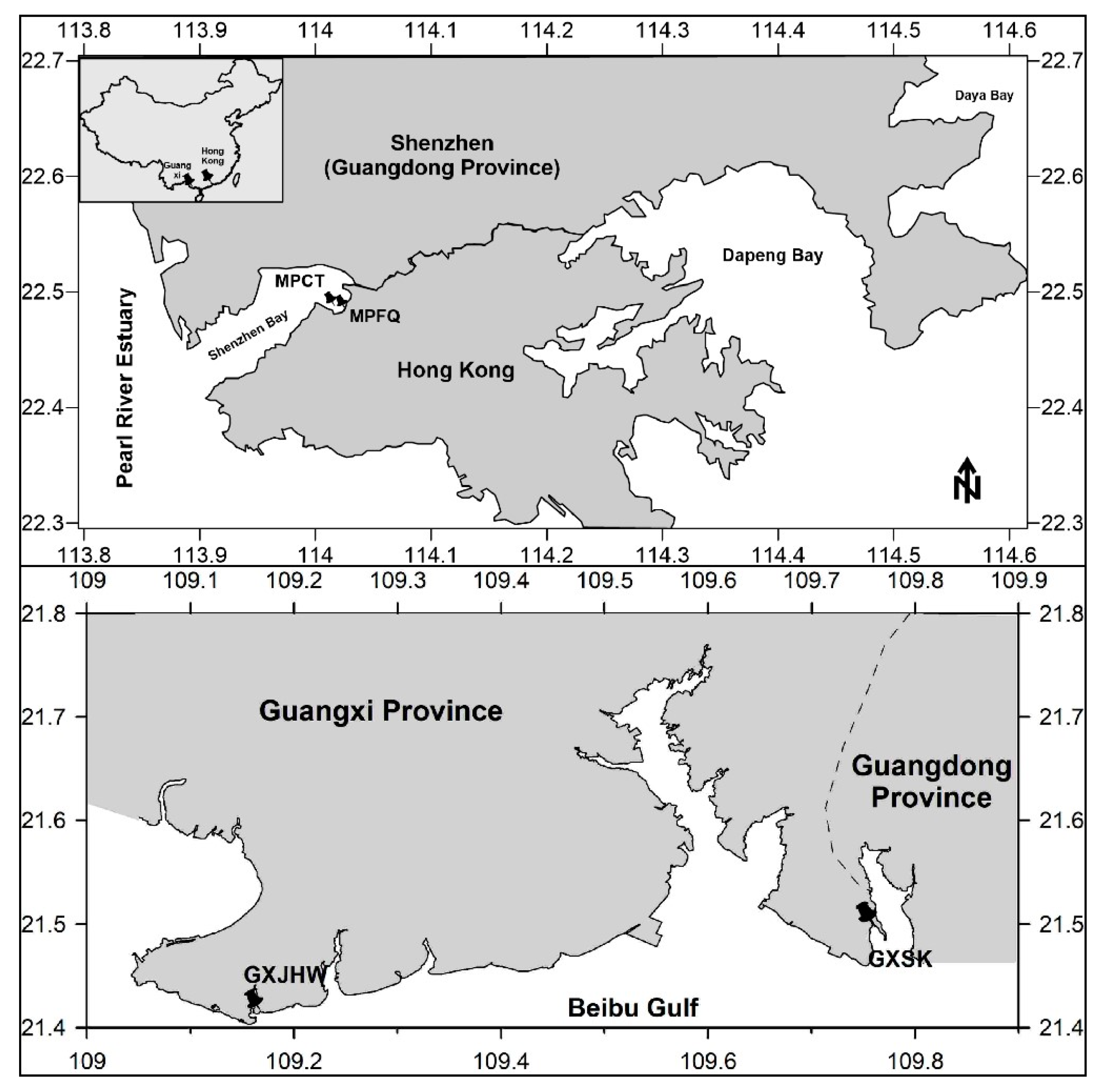
2.1. Site Description
2.2. Sampling
2.3. Analytical Methods
2.3.1. Sedimentary Physicochemical Properties
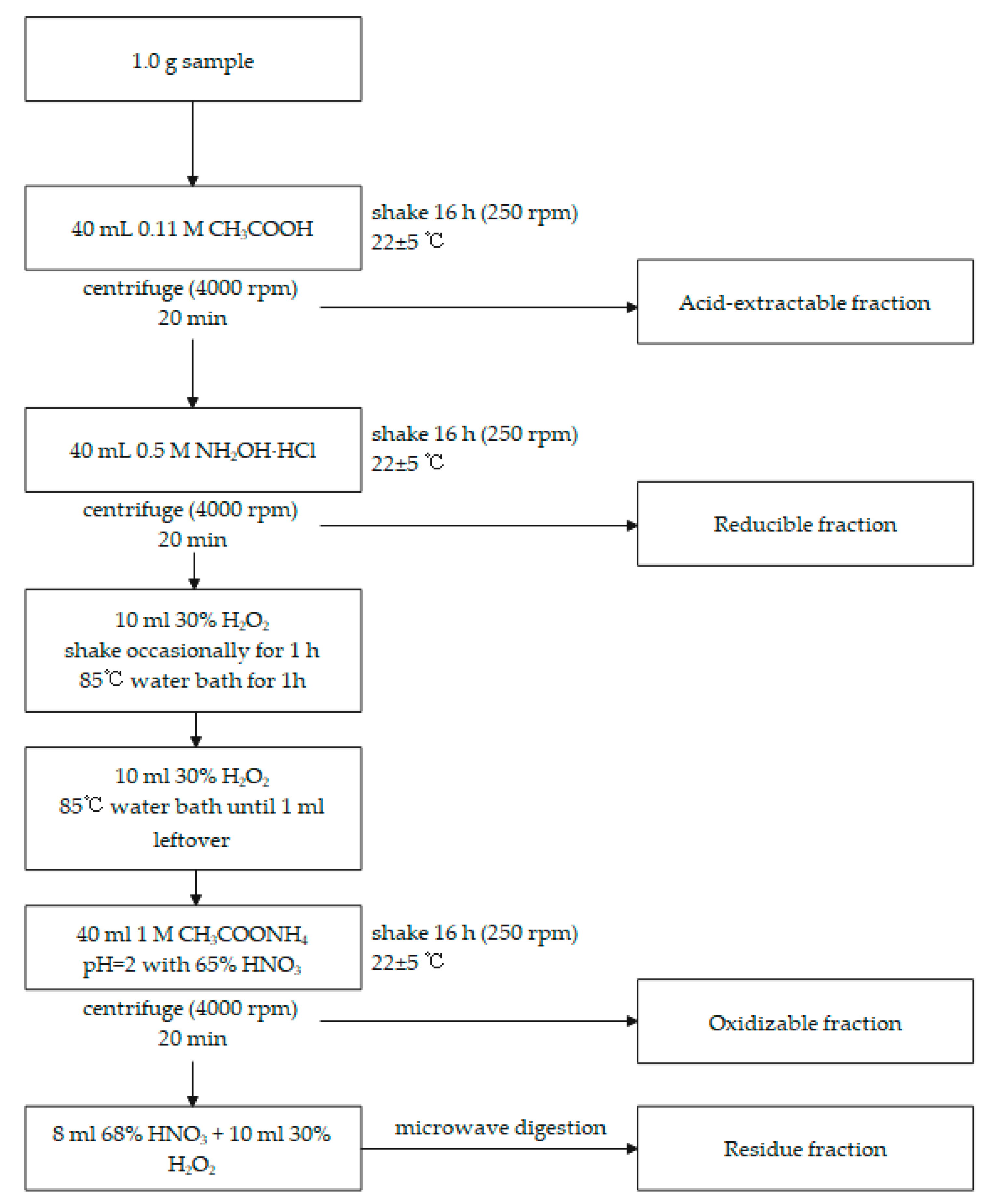
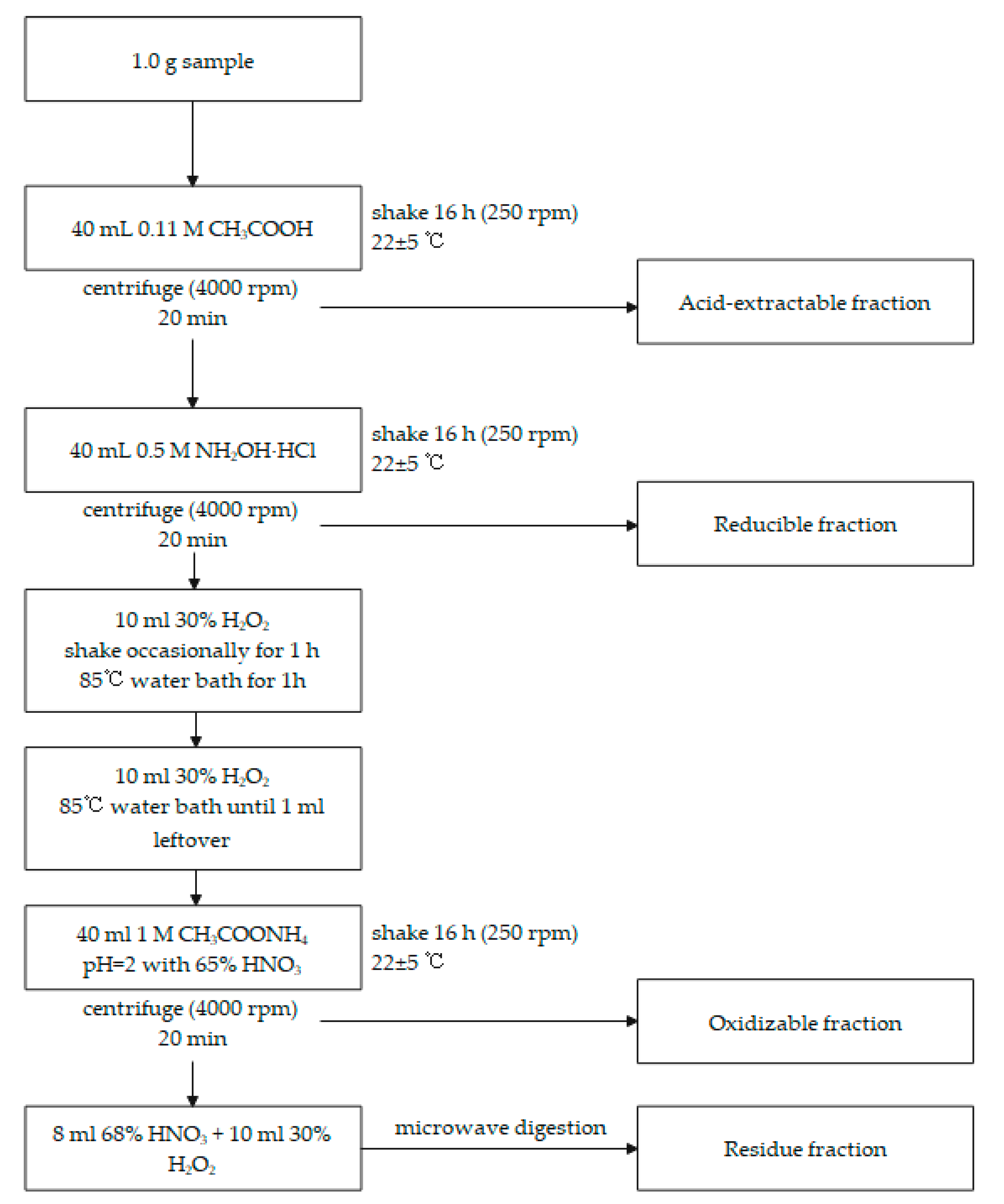
2.3.2. Metal Speciation
2.3.3. DNA Extraction and Illumina HiSeq Sequencing
2.3.4. Sedimentation Rate Measurement
2.4. Quality Assurance and Quality Control
2.5. Methods to Evaluate Heavy Metal Pollution
2.6. Statistical Analysis
3. Results and Discussion
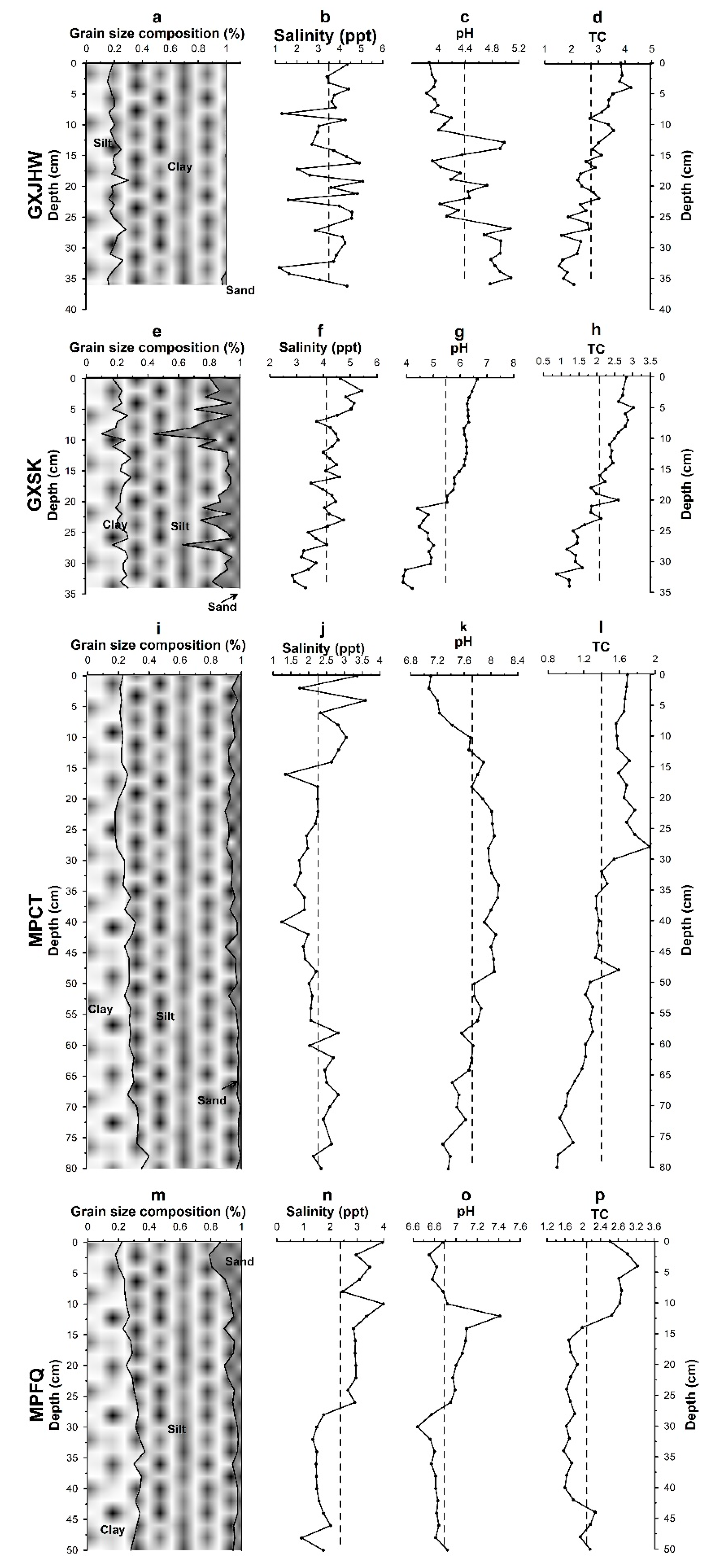
3.1. Physicochemical Properties and Metal Speciation Profiles in Cores from Mangrove Sediments
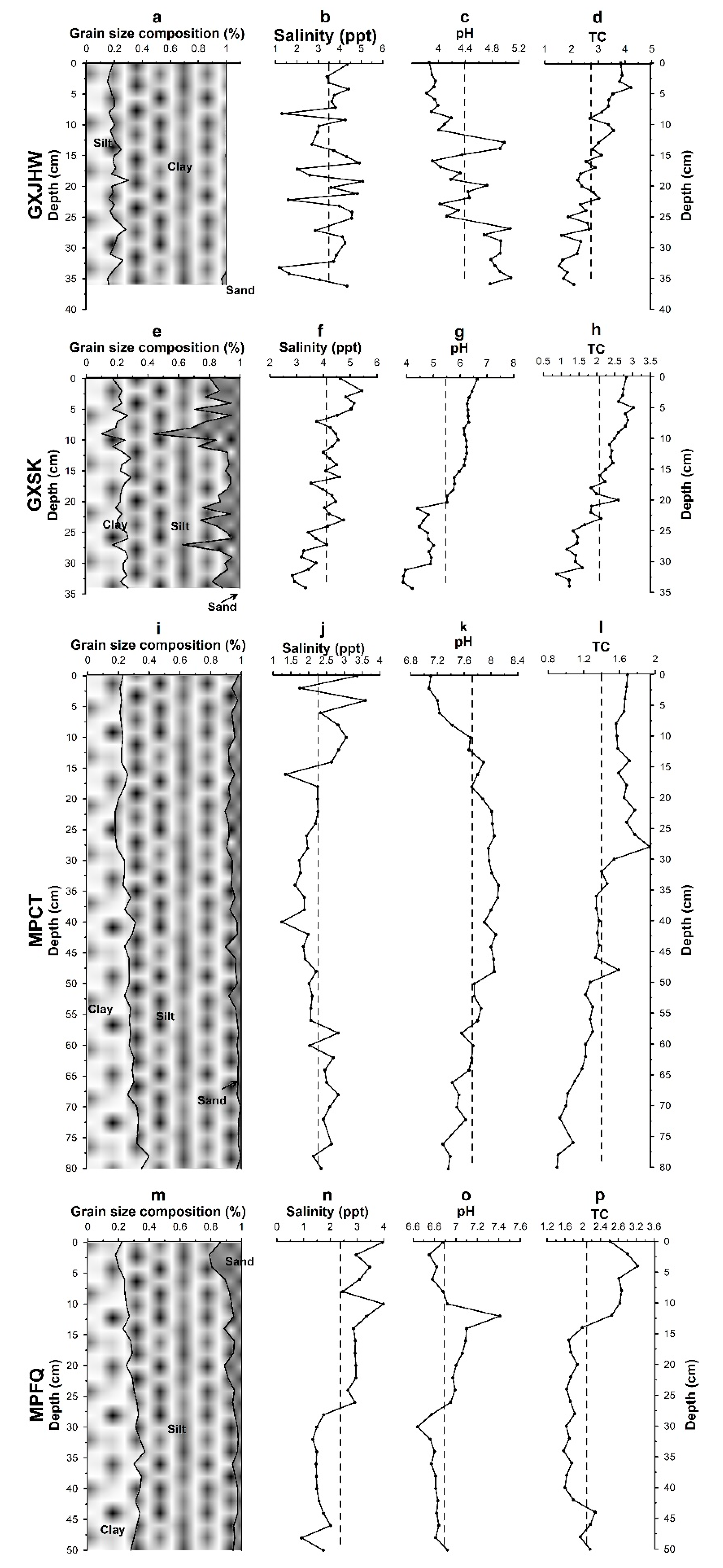
3.1.1. Vertical Profiles of Physicochemical Properties of Sediment Cores
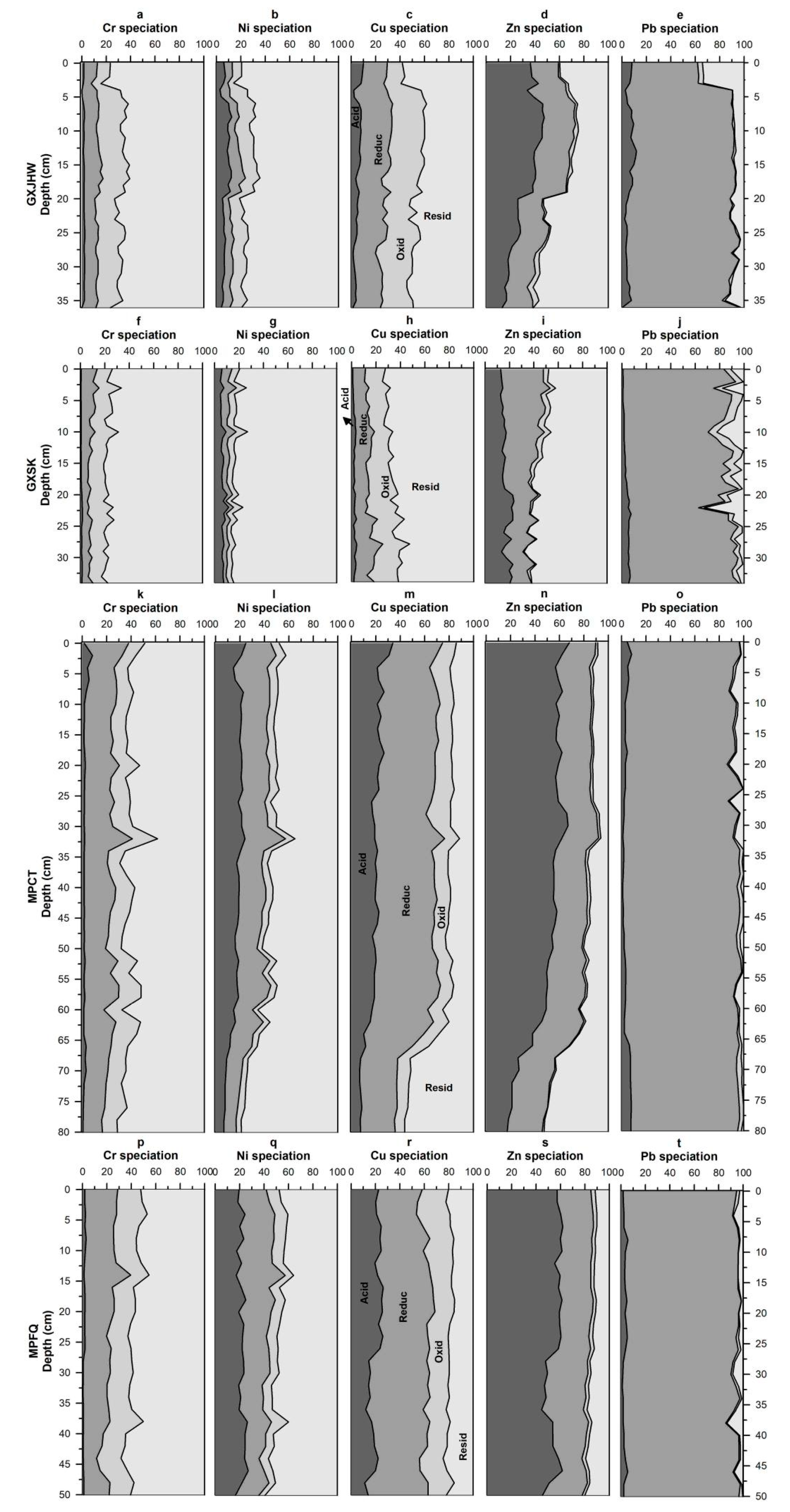
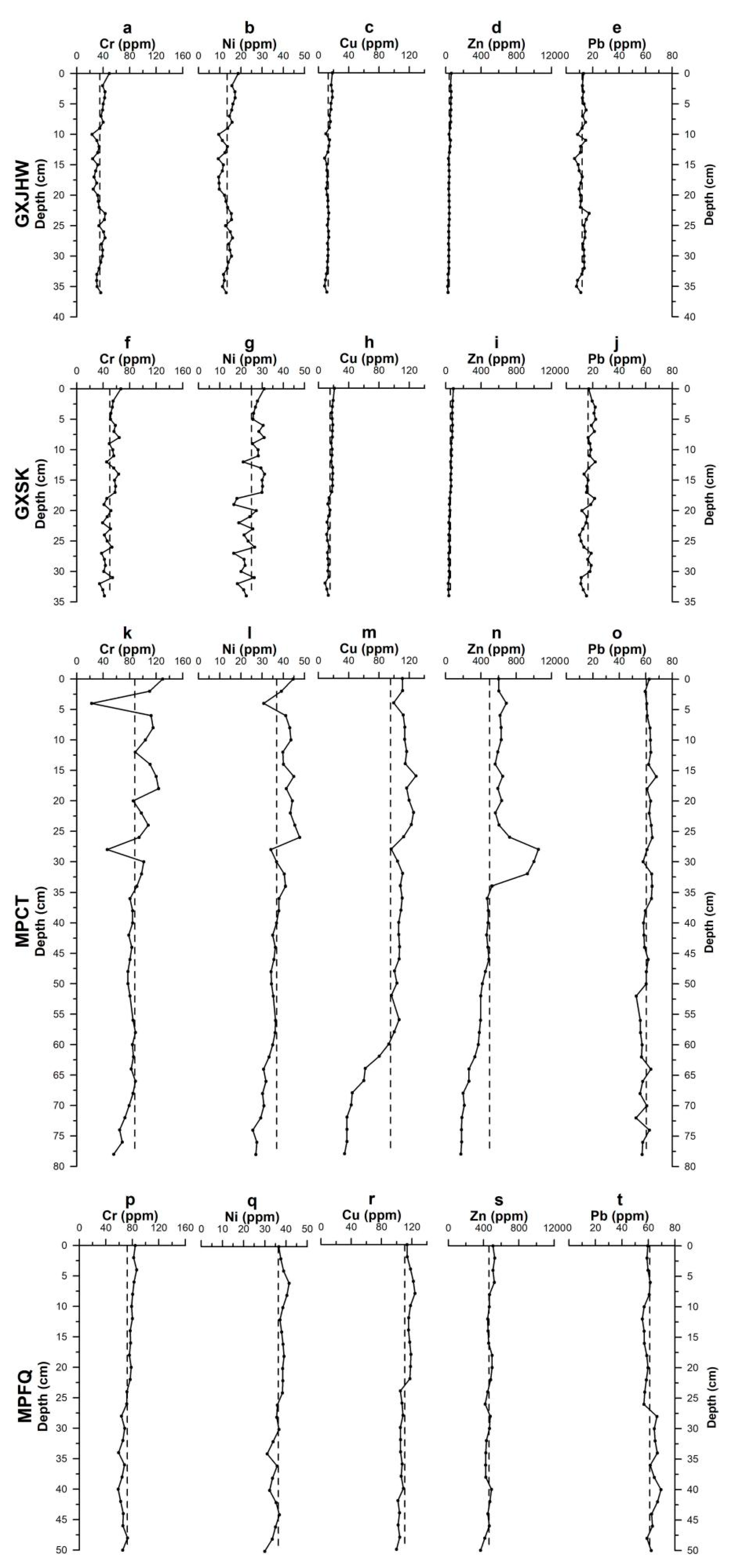
3.1.2. Vertical Profiles of Metal Speciation in Sediment Cores
Acid-Extractable Fraction
Reducible Fraction
Oxidizable and Residue Fractions
3.2. Bacterial Community Structures in Sediment Cores
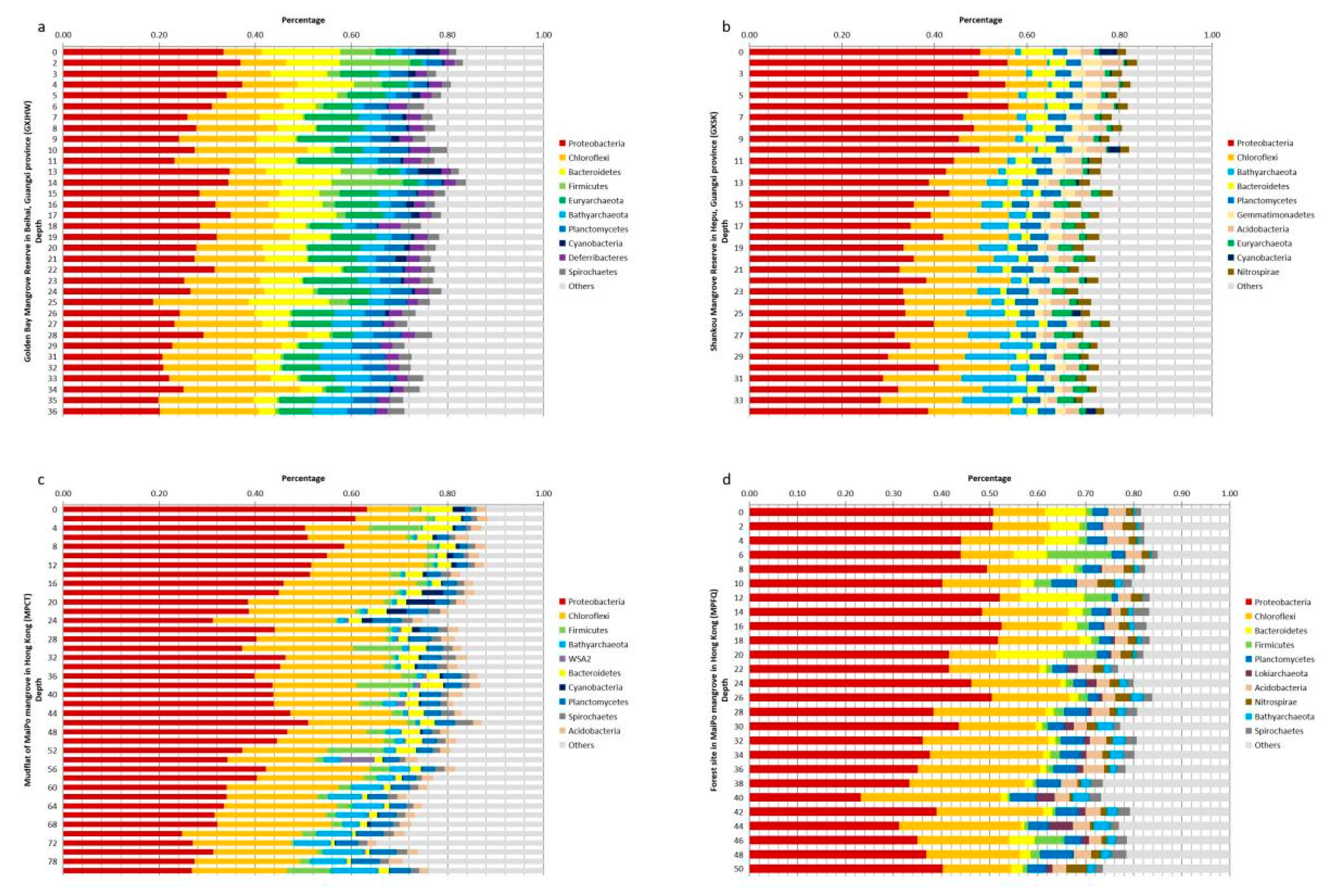
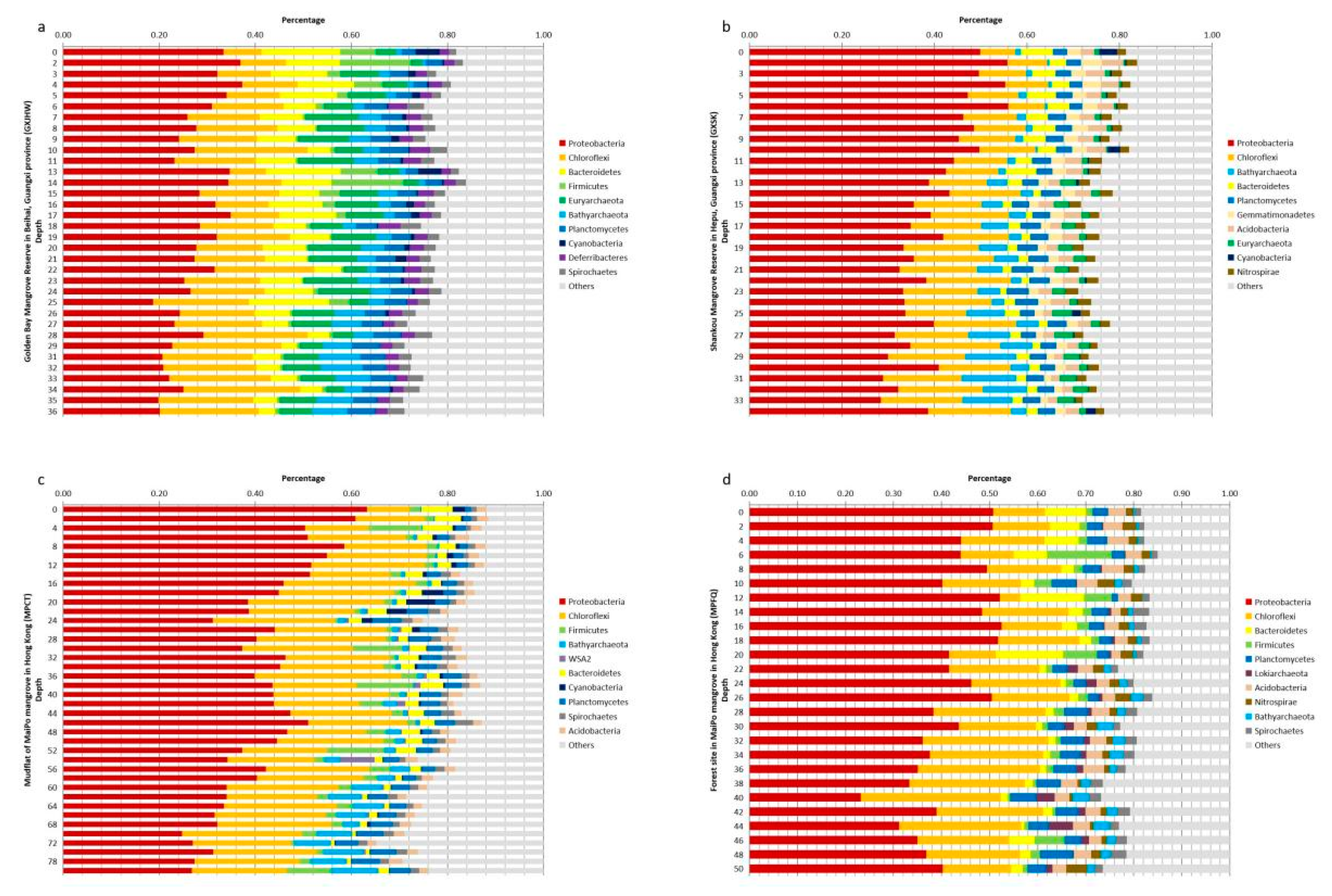
3.2.1. Vertical Profiles of Top 10 Phyla in Sediment Cores
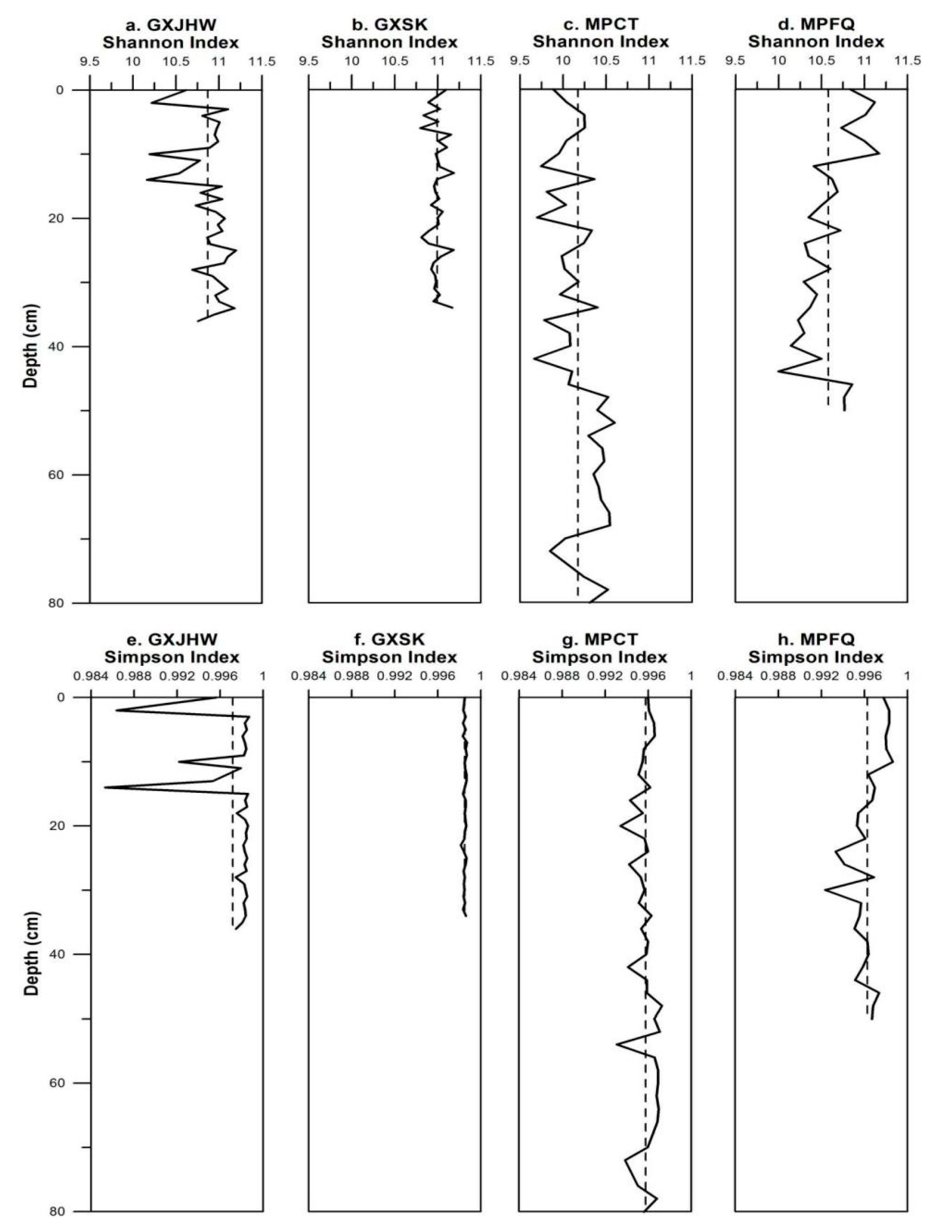
3.2.2. Vertical Variations in Alpha Diversity
3.3. The Influence of Environmental Factors on Bacterial Community Structures in Sediment Cores
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Meaning |
|---|---|
| TCr | Total amount of Cr |
| CrAcid | Acid extractable fraction of Cr |
| CrReduc | Reducible fraction of Cr |
| CrOxid | Oxidizable fraction of Cr |
| CrResid | Residue fraction of Cr |
| Cr_RSP | RSP of Cr |
| Cr_RAC | RAC of Cr |
| TNi | Total amount of Ni |
| NiAcid | Acid extractable fraction of Ni |
| NiReduc | Reducible fraction of Ni |
| NiOxid | Oxidizable fraction of Ni |
| NiResid | Residue fraction of Ni |
| Ni_RSP | RSP of Ni |
| Ni_RAC | RAC of Ni |
| TCu | Total amount of Cu |
| CuAcid | Acid extractable fraction of Cu |
| CuReduc | Reducible fraction of Cu |
| CuOxid | Oxidizable fraction of Cu |
| CuResid | Residue fraction of Cu |
| Cu_RSP | RSP of Cu |
| Cu_RAC | RAC of Cu |
| TZn | Total amount of Zn |
| ZnAcid | Acid extractable fraction of Zn |
| ZnReduc | Reducible fraction of Zn |
| ZnOxid | Oxidizable fraction of Zn |
| ZnResid | Residue fraction of Zn |
| Zn_RSP | RSP of Zn |
| Zn_RAC | RAC of Zn |
| TPb | Total amount of Pb |
| PbAcid | Acid extractable fraction of Pb |
| PbReduc | Reducible fraction of Pb |
| PbOxid | Oxidizable fraction of Pb |
| PbResid | Residue fraction of Pb |
| Pb_RSP | RSP of Pb |
| Pb_RAC | RAC of Pb |
| GXJHW | GXSK | MPCT | MPFQ | |
|---|---|---|---|---|
| Clay | 0.09 | −0.17 | −0.84 | −0.79 |
| Silt | 0.07 | −0.06 | 0.68 | 0.22 |
| pH | −0.47 | 0.95 | 0.24 | 0.47 |
| Eh | 0.46 | −0.84 | −0.16 | −0.91 |
| Salinity | 0.06 | 0.68 | −0.11 | 0.88 |
| TCr | −0.22 | 0.69 | 0.14 | 0.91 |
| CrAcid | −0.04 | −0.93 | 0.36 | 0.90 |
| CrReduc | 0.46 | 0.84 | 0.38 | 0.53 |
| CrOxid | 0.92 | 0.26 | 0.00 | −0.01 |
| CrResid | 0.42 | 0.10 | 0.10 | −0.50 |
| Cr_RSP | 0.35 | 0.35 | 0.19 | 0.66 |
| Cr_RAC | 0.15 | −0.91 | 0.15 | 0.87 |
| TNi | −0.17 | 0.59 | 0.48 | 0.83 |
| NiAcid | 0.61 | −0.38 | 0.77 | −0.12 |
| NiReduc | 0.24 | 0.95 | 0.72 | 0.60 |
| NiOxid | 0.95 | 0.76 | 0.87 | 0.63 |
| NiResid | 0.41 | 0.23 | −0.28 | −0.70 |
| Ni_RSP | 0.42 | 0.34 | 0.71 | 0.77 |
| Ni_RAC | 0.58 | −0.63 | 0.34 | −0.51 |
| TCu | 0.22 | 0.91 | 0.70 | 0.89 |
| CuAcid | 0.40 | 0.02 | 0.65 | 0.81 |
| CuReduc | 0.88 | 0.54 | 0.56 | −0.19 |
| CuOxid | 0.97 | 0.44 | 0.89 | 0.76 |
| CuResid | 0.39 | 0.89 | −0.34 | −0.24 |
| Cu_RSP | 0.71 | −0.77 | 0.68 | 0.57 |
| Cu_RAC | 0.43 | −0.43 | 0.57 | 0.71 |
| TZn | 0.41 | 0.95 | 0.94 | 0.58 |
| ZnAcid | 0.80 | 0.42 | 0.89 | 0.87 |
| ZnReduc | 0.94 | 0.96 | 0.83 | 0.08 |
| ZnOxid | 0.32 | 0.83 | 0.88 | 0.68 |
| ZnResid | −0.34 | 0.67 | −0.41 | −0.93 |
| Zn_RSP | 0.82 | 0.79 | 0.83 | 0.96 |
| Zn_RAC | 0.76 | −0.87 | 0.70 | 0.89 |
| TPb | −0.01 | 0.74 | 0.34 | −0.73 |
| PbAcid | 0.54 | −0.92 | −0.43 | 0.53 |
| PbReduc | 0.96 | 0.80 | −0.14 | 0.26 |
| PbOxid | −0.33 | 0.78 | −0.56 | −0.38 |
| PbResid | −0.14 | 0.13 | 0.36 | 0.05 |
| Pb_RSP | 0.07 | 0.23 | −0.15 | −0.32 |
| Pb_RAC | 0.43 | −0.94 | −0.45 | 0.58 |
| OC | 0.50 | 0.86 | 0.89 | 0.58 |
| Nitrogen | 0.07 | 0.19 | −0.18 | −0.47 |
| Location | Variable | r | p | |
|---|---|---|---|---|
| GXJHW | CrOxid | 0.3007 | 0.002 | ** |
| NiOxid | 0.1352 | 0.036 | * | |
| CuOxid | 0.1571 | 0.008 | ** | |
| ZnAcid | 0.1956 | 0.002 | ** | |
| ZnReduc | 0.1547 | 0.025 | * | |
| PbReduc | 0.1737 | 0.003 | ** | |
| OC | 0.2686 | 0.001 | ** | |
| GXSK | pH | 0.4095 | 0.001 | ** |
| Eh | 0.2646 | 0.002 | ** | |
| Salinity | 0.1942 | 0.008 | ** | |
| CrAcid | 0.3428 | 0.001 | ** | |
| CrReduc | 0.346 | 0.002 | ** | |
| Cr_RAC | 0.2804 | 0.001 | ** | |
| NiReduc | 0.3436 | 0.001 | ** | |
| NiOxid | 0.3935 | 0.001 | ** | |
| Ni_RAC | 0.1386 | 0.033 | * | |
| TCu | 0.3446 | 0.001 | ** | |
| CuResid | 0.3629 | 0.001 | ** | |
| Cu_RSP | 0.305 | 0.001 | ** | |
| TZn | 0.5063 | 0.001 | ** | |
| ZnReduc | 0.5704 | 0.001 | ** | |
| ZnOxid | 0.5144 | 0.001 | ** | |
| ZnResid | 0.1883 | 0.002 | ** | |
| Zn_RSP | 0.461 | 0.001 | ** | |
| Zn_RAC | 0.2047 | 0.011 | * | |
| TPb | 0.3145 | 0.001 | ** | |
| PbAcid | 0.4432 | 0.001 | ** | |
| PbReduc | 0.3459 | 0.001 | ** | |
| PbOxid | 0.3465 | 0.001 | ** | |
| OC | 0.4144 | 0.001 | ** | |
| MPCT | Clay | 0.401 | 0.001 | ** |
| Silt | 0.3096 | 0.001 | ** | |
| NiAcid | 0.4125 | 0.001 | ** | |
| NiReduc | 0.2778 | 0.002 | ** | |
| NiOxid | 0.4014 | 0.001 | ** | |
| Ni_RSP | 0.223 | 0.008 | ** | |
| TCu | 0.3546 | 0.002 | ** | |
| CuReduc | 0.1713 | 0.022 | * | |
| CuOxid | 0.3708 | 0.001 | ** | |
| Cu_RSP | 0.2331 | 0.004 | ** | |
| TZn | 0.4478 | 0.001 | ** | |
| ZnAcid | 0.4121 | 0.001 | ** | |
| ZnReduc | 0.2724 | 0.002 | ** | |
| ZnOxid | 0.329 | 0.001 | ** | |
| Zn_RSP | 0.3961 | 0.001 | ** | |
| Zn_RAC | 0.3337 | 0.002 | ** | |
| PbOxid | 0.1504 | 0.01 | ** | |
| OC | 0.4864 | 0.001 | ** | |
| MPFQ | Clay | 0.4482 | 0.001 | ** |
| Eh | 0.3135 | 0.001 | ** | |
| Salinity | 0.3716 | 0.001 | ** | |
| TCr | 0.3819 | 0.001 | ** | |
| CrAcid | 0.2751 | 0.003 | ** | |
| Cr_RSP | 0.2924 | 0.008 | ** | |
| Cr_RAC | 0.2008 | 0.014 | * | |
| NiReduc | 0.314 | 0.012 | * | |
| TCu | 0.3308 | 0.001 | ** | |
| CuOxid | 0.4424 | 0.002 | ** | |
| ZnAcid | 0.2166 | 0.018 | * | |
| ZnOxid | 0.2334 | 0.039 | * | |
| ZnResid | 0.1962 | 0.008 | ** | |
| Zn_RSP | 0.3419 | 0.001 | ** | |
| OC | 0.5321 | 0.001 | ** | |
References
- Duke, N.C.; Meynecke, J.O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; et al. A World Without Mangroves? Science 2007, 317, 41–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological role and services of tropical mangrove ecosystems: A reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Mille, G.; Guiliano, M.; Asia, L.; Malleret, L.; Jalaluddin, N. Sources of hydrocarbons in sediments of the Bay of Fort de France (Martinique). Chemosphere 2006, 64, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M.; Christoffersen, P.; Tirendi, F. The influence of forest type on microbial-nutrient relationships in tropical mangrove sediments. J. Exp. Mar. Biol. Ecol. 1993, 171, 201–223. [Google Scholar] [CrossRef]
- Kathiresan, K.; Selvam, M.M. Evaluation of beneficial bacteria from mangrove soil. Bot. Mar. 2006, 49, 86–88. [Google Scholar] [CrossRef]
- Piccini, C.; Garcíaalonso, J. Bacterial diversity patterns of the intertidal biofilm in urban beaches of Rio de la Plata. Mar. Pollut. Bull. 2015, 91, 476–482. [Google Scholar] [CrossRef]
- Holguin, G.; Vazquez, P.; Bashan, Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: An overview. Biol. Fert. Soils 2001, 33, 265–278. [Google Scholar] [CrossRef]
- Hou, L.J.; Liu, M.; Xu, S.Y.; Ou, D.N.; Yu, J.; Cheng, S.B.; Liu, X.; Yang, Y. The effects of semi-lunar spring and neap tidal change on nitrification, denitrification, and N2O vertical distribution in the intertidal sediments of the Yangtze Estuary, China. Estuar. Coast. Shelf Sci. 2007, 73, 607–616. [Google Scholar] [CrossRef]
- Abraham, T.J.; Ghosh, S.; Nagesh, T.S.; Sasmal, D. Distribution of bacteria involved in nitrogen and sulphur cycles in shrimp culture systems of west bengal, india. Aquaculture 2004, 239, 275–288. [Google Scholar] [CrossRef]
- Angela, M.G.; Mendonça-Hagler, L.C.S.; Andrew, M. Microbial diversity in Brazilian mangrove sediments: A mini review. Braz. J. Microbiol. 2012, 43, 1242–1254. [Google Scholar]
- Alongi, D.M. Carbon cycling and storage in mangrove forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.J.; Krichman, D.L. Bacterial diversity, community structure and potential growth rates along an estuarine salinity gradient. ISME J. 2013, 7, 210–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, T.; Sorci, J.; Ika, R. Interactions between metals and microbial communities in New Bedford Harbor, Massachusetts. Environ. Health Perspect. Suppl. 1998, 106, 1033–1039. [Google Scholar]
- Gomes, N.C.M.; Borges, L.R.; Paranhos, R.; Pinto, N.F.; Leda, C.S.; Smalla, K. Exploring the diversity of bacterial communities in sediments of urban mangrove forests. FEMS Microbiol. Ecol. 2008, 66, 96–109. [Google Scholar] [CrossRef] [PubMed]
- LaMontagne, M.G.; Leifer, I.; Bergmann, S.; Van, D.W.L.C.; Holden, P.A. Bacterial diversity in marine hydrocarbon seep sediments. Environ. Microbiol. 2004, 6, 799–808. [Google Scholar] [CrossRef]
- Basak, P.; Pramanik, A.; Sengupta, S.; Nag, S.; Bhattacharyya, M. Bacterial diversity assessment of pristine mangrove microbial community from Dhulibhashani, Sundarbans using 16S rRNA gene tag sequencing. Genom. Data 2016, 7, 76–78. [Google Scholar] [CrossRef] [Green Version]
- Dudani, S.N.; Lakhmapurkar, J.; Gavali, D.; Patel, T. Heavy metal accumulation in the mangrove ecosystem of south gujarat coast, India. Turk. J. Fish. Aquat. Sci. 2017, 17, 755–766. [Google Scholar]
- Bernard, D.; Pascaline, H.; Jeremie, J.J. Distribution and origin of hydrocarbons in sediments from lagoons with fringing mangrove communities. J. Mar. Pollut. Bull. 1996, 32, 734–739. [Google Scholar] [CrossRef]
- Kleskowski, E.J.; Corredor, J.E.; Morell, J.M.; Del Castillo, C.A. Petroleum pollution and mutation in mangroves. J. Mar. Pollut. Bull. 1994, 28, 166–169. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: Impact on microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, M.; Nagumo, T. Effects of heavy metal accumulation in apple orchard soils on microbial biomass and microbial activities. Soil Sci. Plant Nutr. 1997, 43, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Brookes, P.C.; McGrath, S.P. Effects of metal toxicity on the size of the soil microbial biomass. Soil Sci. 1984, 35, 341–346. [Google Scholar] [CrossRef]
- Ni, D.; Horton, D.J.; Rui, J.; Henson, M.W.; Jiang, W.; Huang, X.; Learman, D.R. High concentrations of bioavailable heavy metals impact freshwater sediment microbial communities. Ann. Microbiol. 2016, 66, 1003–1012. [Google Scholar] [CrossRef]
- Thavamani, P.; Malik, S.; Beer, M.; Megharaj, M.; Naidu, R. Microbial activity and diversity in long-term mixed contaminated soils with respect to polyaromatic hydrocarbons and heavy metals. J. Environ. Manag. 2012, 99, 10–17. [Google Scholar] [CrossRef]
- Dell’Anno, A.; Mei, M.L.; Ianni, C.; Danovaro, R. Impact of bioavailable heavy metals on bacterial activities in coastal marine sediments. World J. Microbiol. Biotechnol. 2003, 19, 93–100. [Google Scholar] [CrossRef]
- Chen, S.; Liao, J.; Zhang, Q.; Ding, S.; He, M.; Huang, D. Spatial Variability and Influencing Factors of Cr, Ni, Cu, Zn and Pb in Five Mangroves Reserves of South China. IOP Conf. Ser. Earth Environ. Sci. 2021, 690, 012017. [Google Scholar] [CrossRef]
- Feng, J.X.; Zhu, X.S.; Wu, H.; Ning, C.X.; Lin, G.H. Distribution and ecological risk assessment of heavy metals in surface sediments of a typical restored mangrove–aquaculture wetland in Shenzhen, China. Mar. Pollut. Bull. 2017, 124, 1033–1039. [Google Scholar] [CrossRef]
- Tessier, A.; Campbell, P.; Bisson, M. Sequential extraction procedure for the speciation of pareticulate trace-metrals. Anal. Chem. 1979, 51, 844–851. [Google Scholar] [CrossRef]
- Hall, G.; Vaive, J.E.; Beer, R.; Hoashi, M. Selective leaches revisited, with emphasis on the amorphous Fe oxyhydroxide phase extraction. J. Geochem. Explor. 1996, 56, 59–78. [Google Scholar] [CrossRef]
- Hall, G.E.M.; Pelchat, P. Comparibility of results obtained by the use of different selective extraction schemer for the determination of element forms in soils. Water Air Soil Pollut. 1999, 112, 41–53. [Google Scholar] [CrossRef]
- Rauret, G.; Lopez-Sanchez, J.; Sahuquillo, A.; Rubio, R.; Davidson, C.; Ure, A. Improvement of the BCR three step sequential extraction procedure prior to the certification of new sediment and soil reference materials. J. Environ. Monit. 1999, 1, 57–61. [Google Scholar] [CrossRef] [PubMed]
- IAEA. Measurement of Radionuclides in Food and Environment; Technical Reports Series, no. 295; IAEA: Vienna, Austria, 1989. [Google Scholar]
- Hakanson, L. An ecological risk index for aquatic pollution-control—A sedimentological approach. Water Res. 1980, 14, 975–1001. [Google Scholar] [CrossRef]
- Zhang, H.; Walker, T.; Davis, E.; Ma, G. Ecological risk assessment of metals in small craft harbour sediments in Nova Scotia, Canada. Mar. Pollut. Bull. 2019, 146, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Micó, C.; Recatalá, L.; Peris, M.; Sánchez, J. Assessing heavy metal sources in agricultural soils of an European Mediterranean area by multivariate analysis. Chemosphere 2006, 65, 863–872. [Google Scholar] [CrossRef]
- Akcay, H.; Oguz, A.; Karapire, C. Study of heavy metal pollution and speciation in Buyak Menderes and Gediz river sediments. Water Res. 2003, 37, 813–822. [Google Scholar] [CrossRef]
- Gusiatin, Z.M.; Kulikowska, D. The usability of the IR, RAC and MRI indices of heavy metal distribution to assess the environmental quality of sewage sludge composts. Waste Manag. 2014, 34, 1227–1236. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.P.; Xu, C.X.; An, Z.Y.; Wang, S.M. Analysis and evaluation of the source of heavy metals in water of the River Changjiang. Environ. Monit. Assess. 2011, 173, 301–313. [Google Scholar] [CrossRef]
- Harikumar, P.S.; Nasir, R.P. Ecotoxicological impact assessment of heavy metals in core sediments of a tropical estuary. Ecotoxicol. Environ. Saf. 2010, 73, 1742–1747. [Google Scholar] [CrossRef]
- Islam, M.S.; Ahmed, M.K.; Raknuzzaman, M.; Habibullah-Al-Mamun, M.; Islam, M.K. Heavy metal pollution in surface water and sediment: A preliminary assessment of an urban river in a developing country. Ecol. Indic. 2015, 48, 282–291. [Google Scholar] [CrossRef]
- Maslennikova, S.; Larina, N.; Larin, S. The effect of sediment grain size on heavy metal content. Solid State Phenom. 2012, 6, 43–54. [Google Scholar]
- Murray, K.S.; Cauvet, D.; Lybeer, M.; Thomas, J.C. Particle size and chemical control of heavy metals in bed sediment from the Rouge River, Southeast Michigan. Environ. Sci. Technol. 1999, 33, 987–992. [Google Scholar] [CrossRef]
- Soto-Jimenez, M.F.; Paez-Osuna, F. Distribution and normalization of heavy metal concentrations in mangrove and lagoonal sediments from Mazatlan Harbor (SE Gulf of California). Estuar. Coast. Shelf Sci. 2001, 53, 259–274. [Google Scholar] [CrossRef]
- Horowitz, A.J.; Elrick, K.A. The relation of stream sediment surface area, grain size and composition to trace element chemistry. Appl. Geochem. 1987, 2, 437–451. [Google Scholar] [CrossRef]
- Ray, A.; Tripathy, S.; Patra, S.; Sanna, V. Assessment of Godavari estuarine mangrove ecosystem through trace metal studies. Environ. Int. 2006, 32, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Defew, L.H.; Mair, J.M.; Guzman, H.M. An assessment of metal contamination in mangrove sediments and leaves from Punta Mala Bay, Pacific Panama. Mar. Pollut. Bull. 2005, 50, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, S.Z.; Mohamat-Yusuff, F.; Arai, T.; Ismail, A.; Miyazaki, N. An assessment of selected trace elements in intertidal surface sediments collected from the Peninsular Malaysia. Environ. Monit. Assess. 2010, 169, 457–472. [Google Scholar] [CrossRef]
- Cheng, W.H.; Yap, C.K. Potential human health risks from toxic metals via mangrove snail consumption and their ecological risk assessments in the habitat sediment from Peninsular Malaysia. Chemosphere 2015, 135, 156–165. [Google Scholar] [CrossRef]
- Amin, B.; Ismail, A.; Arshad, A.; Yap, C.K.; Kamarudin, M.S. Anthropogenic impacts on heavy metal concentrations in the coastal sediments of Dumai, Indonesia. Environ. Monit. Assess. 2009, 148, 291–305. [Google Scholar] [CrossRef]
- Jayachandran, S.; Chakraborty, P.; Ramteke, D.; Chennuri, K.; Chakraborty, S. Effect of pH on transport and transformation of Cu-sediment complexes in mangrove systems. Mar. Pollut. Bull. 2018, 133, 920–929. [Google Scholar] [CrossRef]
- Ramos, L.; González, M.J.; Hernández, L.M. Sequential extraction of copper, lead, cadmium, and zinc in sediments from Ebro River (Spain): Relationship with levels detected in earthworms. Bull. Environ. Contam. Toxicol. 1999, 62, 301–308. [Google Scholar] [CrossRef]
- Tam, N.F.Y.; Yao, M.W.Y. Normalisation and heavy metal contamination in mangrove sediments. Sci. Total Environ. 1998, 216, 33–39. [Google Scholar] [CrossRef]
- Li, X.; Poon, C.; Liu, P.S. Heavy metal contamination of urban soils and street dusts in Hong Kong. Appl. Geochem. 2001, 16, 1361–1368. [Google Scholar] [CrossRef]
- Caregnato, F.F.; Koller, C.E.; MacFarlane, G.R.; Moreira, J. The glutathione antioxidant system as a biomarker suite for the assessment of heavy metal exposure and effect in the grey mangrove, Avicennia marina (Forsk.) Vierh. Mar. Pollut. Bull. 2008, 56, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, G.R.; Burchett, M.D. Photosynthetic pigments and peroxidase activity as indicators of heavy metal stress in the grey mangrove, Avicennia marina (Forsk.) Vierh. Mar. Pollut. Bull. 2001, 42, 233–240. [Google Scholar] [CrossRef]
- Power, A.E.; Peters, C.E.; Firman, C.J.; Gassman, J.N.; Richmond, H.R. Ecotoxicology of tropical marine ecosystems. Environ. Toxicol. Chem. 1997, 16, 12–40. [Google Scholar]
- Gu, Y.G.; Wang, Z.H.; Lu, S.H.; Jiang, S.J.; Mu, D.H.; Shu, Y.H. Multivariate statistical and GIS-based approach to identify source of anthropogenic impacts on metallic elements in sediments from the mid Guangdong coasts, China. Environ. Pollut. 2012, 163, 248–255. [Google Scholar] [CrossRef]
- Zhou, F.; Guo, H.; Hao, Z. Spatial distribution of heavy metals in Hong Kong’s marine sediments and their human impacts: A GIS-based chemometric approach. Mar. Pollut. Bull. 2007, 54, 1372–1384. [Google Scholar] [CrossRef]
- Tam, N.F.; Wong, Y.S. Spatial variation of heavy metals in surface sediments of Hong Kong mangrove swamps. Environ. Pollut. 2000, 110, 195–205. [Google Scholar] [CrossRef]
- Liu, J.; Ma, K.; Qu, L. Ecological risk assessments and context-dependence analysis of heavy metal contamination in the sediments of mangrove swamp in Leizhou Peninsula, China. Mar. Pollut. Bull. 2015, 100, 224–230. [Google Scholar] [CrossRef]
- Vane, C.H.; Harrison, I.; Kim, A.W.; Moss-Hayes, V.; Vickers, B.P.; Hong, K. Organic and metal contamination in surface mangrove sediments of South China. Mar. Pollut. Bull. 2009, 58, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, G.W.; Ding, S.L.; Zhang, Q.J.; Huang, D.D.; Shang, C.J. A thirty-year record of PTE pollution in mangrove sediments: Implications for human activities in two major chinese metropolises, Shenzhen and Hong Kong. Sustain. Cities Soc. 2021, 68, 102766. [Google Scholar] [CrossRef]
- Almeida, C.M.R.; Mucha, A.P.; Vasconcelos, M.T.S.D. The role of a salt marsh plant on trace metal bioavailability in sediments estimation by different chemical approaches. Environ. Sci. Pollut. Res. 2005, 12, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Turki, A. Distribution and speciation of heavy metals in surficial sediments from the Estuary, north-east England. Mar. Pollut. Bull. 1997, 34, 768–779. [Google Scholar] [CrossRef]
- Tessier, A.; Campbell, P.; Auclair, J.C.; Bisson, M. Relationship between the partitioning of the trace metals in sediments and their accumulations in the tissue of the freshwater mollusk elliptio complanata in a mining area. Can. J. Fish. Quat. Sci. 1984, 41, 1463–1472. [Google Scholar]
- Whitall, D.; Mason, A.; Pait, A.; Brune, L.; Fulton, M.; Wirth, E.; Vandiver, L. Organic and metal contamination in marine surface sediments of Guánica Bay, Puerto Rico. Mar. Pollut. Bull. 2014, 80, 293–301. [Google Scholar] [CrossRef]
- Astruc, T.M. Fractionation studies of trace elements in contaminated soils and sediments: A review of sequential extraction procedures. TrAC Trends Anal. Chem. 2002, 21, 451–467. [Google Scholar]
- Li, X.; Wang, Y.; Li, B.H.; Feng, C.H.; Chen, Y.X.; Shen, Z.Y. Distribution and speciation of heavy metals in surface sediments from the yangtze estuary and coastal areas. Environ. Earth Sci. 2012, 69, 1537–1547. [Google Scholar] [CrossRef]
- Chaoui, A.; Mazhoudi, S.; Ghorbal, M.H.; Ferjani, E.E. Cadmium and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in bean (Phaseolus vulgaris L.). Plant Sci. 1997, 127, 139–147. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Wang, W.P. Research on key service factors’ identification of producer service value network of China’s high-end manufacturing based on grey relational analysis. Adv. Mater. Res. 2014, 933, 807–812. [Google Scholar] [CrossRef]
- Li, Y.; Wen, H.; Chen, L.; Yin, T. Succession of bacterial community structure and diversity in soil along a chronosequence of reclamation and re-vegetation on coal mine spoils in China. PLoS ONE 2014, 9, e115024. [Google Scholar] [CrossRef]
- Zhou, Y.W.; Zhao, B.; Peng, Y.S.; Chen, G.Z. Influence of mangrove reforestation on heavy metal accumulation and speciation in intertidal sediments. Mar. Pollut. Bull. 2010, 60, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Noronha-D’Mello, C.A.; Nayak, G.N. Geochemical characterization of mangrove sediments of the Zuari estuarine system, West coast of India. Estuar. Coast. Shelf Sci. 2015, 167, 313–325. [Google Scholar] [CrossRef]
- Zhang, L.; Xin, Y.; Feng, H.; Jing, Y.; Tong, O.; Yu, X.; Liang, R.; Gao, C.; Chen, W. Heavy metal contamination in western Xiamen Bay sediments and its vicinity, China. Mar. Pollut. Bull. 2007, 54, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.W.; Yang, S.Y. Heavy metal enrichments in the Changjiang (Yangtze River) catchment and on the inner shelf of the East China Sea over the last 150 years. Sci. Total Environ. 2016, 543, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Prosi, F. Factors controlling biological availability and toxic effects of lead in aquatic organisms. Sci. Total Environ. 1989, 79, 157–169. [Google Scholar] [CrossRef]
- Shao, Z.Y.; Wang, Y.; Zhang, J.; Yang, C.; Zhou, G.; Yang, R.Y. The bacterial community structure associated with a copper-tolerant plant, Artemisia capillaries Thunb., and its influencing factors. Acta Ecol. Sin. 2017, 37, 7679–7688. [Google Scholar]
- Schneider, A.R.; Gommeaux, M.; Duclercq, J.; Fanin, N.; Conreux, A.; Alahmad, A.; Lacoux, J.; Roger, D.; Spicher, F.; Ponthieu, M.; et al. Response of bacterial communities to Pb smelter pollution in contrasting soils. Sci. Total Environ. 2017, 605, 436–444. [Google Scholar] [CrossRef]
- Xia, N.; Xia, X.; Zhu, B.; Zheng, S.; Zhuang, J. Bacterial diversity and community structure in the sediment of the middle and lower reaches of the Yellow River, the largest turbid river in the world. Aquat. Microb. Ecol. 2013, 71, 43–55. [Google Scholar] [CrossRef]
- Ghosh, A.; Dey, N.; Bera, A.; Tiwari, A.; Sathyaniranjan, K.B.; Chakrabarti, K.; Chattopadhyay, D. Culture independent molecular analysis of bacterial communities in the mangrove sediment of Sundarban, India. Saline Syst. 2010, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; Volume 46, pp. 7–45. [Google Scholar]
- Peng, Z.; Song, J.; Yuan, H. Persistent organic pollutant residues in the sediments and mollusks from the bohai sea coastal areas, north china: An overview. Environ. Int. 2009, 35, 632–646. [Google Scholar]
- Suhadolc, M.; Schroll, R.; Gattinger, A.; Schloter, M.; Munch, J.C.; Lestan, D. Effects of modified pb-, zn-, and cd- availability on the microbial communities and on the degradation of isoproturon in a heavy metal contaminated soil. Soil Biol. Biochem. 2004, 36, 1943–1954. [Google Scholar] [CrossRef]
- Hiroki, M. Effects of heavy metal contamination on soil microbial population. Soil Sci. Plant Nutr. 1992, 38, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Wasilkowski, D.; Mrozik, A.; Piotrowska-Seget, Z.; Krzyżak, J.; Pogrzeba, M.; Płaza, G. Changes in enzyme activities and microbial community structure in heavy metal-contaminated soil underin situaided phytostabilization. CLEAN Soil Air Water 2014, 42, 1618–1625. [Google Scholar] [CrossRef]
- Bisessar, S. Effect of heavy metals on microorganisms in soils near a secondary lead smelter. Water Air Soil Pollut. 1982, 17, 305–308. [Google Scholar] [CrossRef]
- Oliveira, A.; Pampulha, M.E. Effects of long-term heavy metal contamination on soil microbial characteristics. Biosci. Bioeng. 2006, 102, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Ma, Z.; Van der Kuijp, T.J.; Yuan, Z.; Huang, L. A review of soil heavy metal pollution from mines in China: Pollution and health risk assessment. Sci. Total Environ. 2014, 468, 843–853. [Google Scholar] [CrossRef]
- Clark, M.W.; Mcconchie, D.M.; Lewis, D.W.; Saenger, P. Redox stratification and heavy metal partitioning in Avicennia-dominated mangrove sediments: A geochemical model. Chem. Geol. 1998, 149, 147–171. [Google Scholar] [CrossRef]
- Fernandes, H.M. Heavy metal distribution in sediments and ecological risk assessment: The role of diagenetic processes in reducing metal toxicity in bottom sediments. Environ. Pollut. 1997, 97, 317–325. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Cheung, R.Y.H.; Chan, K.M.; Wong, M.H. Metal concentrations in sediments and Tilapia collected from inland water of Hong Kong. Water Res. 1998, 32, 3331–3340. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Y.; Shen, Z.; Niu, J.; Tang, Z. Distribution and speciation of heavy metals in sediments from the mainstream, tributaries, and lakes of the yangtze river catchment of wuhan, china. J. Hazard. Mater. 2009, 166, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Middelburg, J.J. Reviews and syntheses: To the bottom of carbon processing at the seafloor. Biogeosciences 2018, 15, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Cathalot, C.; Van Oevelen, D.; Cox, T.J.S.; Kutti, T.; Lavaleye, M.; Duineveld, G. Cold-water coral reefs and adjacent sponge grounds: Hotspots of benthic respiration and organic carbon cycling in the deep sea. Front. Mar. Sci. 2015, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Rice, D.; Rhoads, D. Early Diagenesis of Organic Matter and the Nutritional Value of Sediment. In Ecology of Marine Deposit Feeders; Springer: New York, NY, USA, 1989; Volume 31. [Google Scholar]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Tangherlini, M.; Lo Martire, M.; Nishizawa, M. Drivers of Bacterial alpha- and beta-Diversity Patterns and Functioning in Subsurface Hadal Sediments. Front. Microbiol. 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Marchand, C.; Allenbach, M.; Lallier-Verges, E. Relationships between heavy metals distribution and organic matter cycling in mangrove sediments (Conception Bay, New Caledonia). Geoderma 2011, 160, 444–456. [Google Scholar] [CrossRef] [Green Version]
- Bayen, S. Occurrence, bioavailability and toxic effects of trace metals and organic contaminants in mangrove ecosystems: A review. Environ. Int. 2012, 48, 84–101. [Google Scholar] [CrossRef]


| Sites | Cr | Ni | Cu | Zn | Pb | RI | |
|---|---|---|---|---|---|---|---|
| GXJHW | content (μg/g) | 35.02 ± 6.05 | 13.49 ± 2.42 | 12.86 ± 2.64 | 40.87 ± 11.21 | 11.98 ± 2.31 | 7.12 ± 1.41 |
| 1.17 ± 0.20 | 0.90 ± 0.16 | 2.14 ± 0.44 | 0.51 ± 0.14 | 2.40 ± 0.46 | |||
| GXSK | content (μg/g) | 49.99 ± 8.22 | 24.97 ± 4.44 | 15.40 ± 3.19 | 53.09 ± 16.47 | 16.51 ± 3.49 | 9.86 ± 2.01 |
| 1.67 ± 0.27 | 1.66 ± 0.30 | 2.57 ± 0.53 | 0.66 ± 0.21 | 3.30 ± 0.70 | |||
| MPCT | content (μg/g) | 85.68 ± 24.46 | 36.17 ± 7.02 | 94.73 ± 27.97 | 497.99 ± 211.92 | 59.90 ± 4.73 | 39.26 ± 9.54 |
| 2.86 ± 0.82 | 2.41 ± 0.47 | 15.79 ± 4.66 | 6.22 ± 2.65 | 11.98 ± 0.95 | |||
| MPFQ | content (μg/g) | 72.39 ± 8.08 | 36.40 ± 2.79 | 110.80 ± 7.09 | 459.87 ± 37.42 | 61.04 ± 3.78 | 41.26 ± 2.86 |
| 2.41 ± 0.27 | 2.43 ± 0.19 | 18.47 ± 1.18 | 5.75 ± 0.47 | 12.21 ± 0.76 | |||
| Singapore [45] | content (μg/g) | 16.61–32.07 | 7.44–11.65 | 7.06–32.00 | 51.24–120.23 | 12.28–30.98 | |
| Punta Mala Bay, Panama [46] | content (μg/g) | 23.3 | 27.3 | 56.3 | 105 | 78.2 | |
| Peninsular Malaysia [47,48] | content (μg/g) | 0.71–6.53 | 2.41–36.29 | 1.63–150.81 | 23.70–607.20 | 7.97–93.11 | |
| Dumai coast Indonesia [49] | content (μg/g) | - | 7.26–19.97 | 1.61–13.84 | 31.49–87.11 | 14.63–84.90 |
| Site | Variable | Shannon | Simpson | ||
|---|---|---|---|---|---|
| r | p | r | p | ||
| GXJHW | CrReduc | −0.415 | 0.006 * | −0.190 | 0.134 |
| CrOxid | −0.207 | 0.112 | −0.033 | 0.425 | |
| NiAcid | 0.303 | 0.036 * | −0.159 | 0.178 | |
| NiOxid | 0.311 | 0.033 * | −0.100 | 0.280 | |
| Ni_RAC | −0.277 | 0.051 | −0.145 | 0.199 | |
| CuOxid | 0.316 | 0.03 * | −0.101 | 0.278 | |
| Cu_RSP | −0.124 | 0.235 | −0.136 | 0.214 | |
| ZnAcid | 0.305 | 0.035 * | −0.078 | 0.326 | |
| ZnReduc | −0.366 | 0.014 * | −0.112 | 0.257 | |
| PbReduc | −0.229 | 0.090 | −0.002 | 0.496 | |
| OC | −0.239 | 0.080 | −0.053 | 0.379 | |
| GXSK | pH | 0.093 | 0.300 | 0.018 | 0.148 |
| Eh | −0.171 | 0.166 | −0.317 | 0.034 * | |
| Salinity | −0.204 | 0.124 | −0.103 | 0.281 | |
| TCr | 0.107 | 0.273 | 0.117 | 0.254 | |
| CrAcid | −0.198 | 0.130 | −0.323 | 0.031 | |
| CrReduc | 0.250 | 0.077 | 0.234 | 0.091 | |
| Cr_RAC | −0.175 | 0.161 | −0.250 | 0.077 | |
| NiReduc | 0.223 | 0.102 | 0.244 | 0.082 | |
| NiOxid | 0.032 | 0.428 | 0.193 | 0.137 | |
| Ni_RAC | −0.152 | 0.195 | −0.108 | 0.271 | |
| TCu | 0.071 | 0.345 | 0.119 | 0.252 | |
| CuResid | 0.198 | 0.131 | 0.251 | 0.076 | |
| Cu_RSP | −0.175 | 0.161 | −0.185 | 0.147 | |
| TZn | 0.078 | 0.330 | 0.137 | 0.220 | |
| ZnReduc | 0.146 | 0.205 | 0.223 | 0.102 | |
| ZnOxid | 0.129 | 0.233 | 0.195 | 0.135 | |
| ZnResid | 0.077 | 0.333 | 0.149 | 0.199 | |
| Zn_RSP | 0.048 | 0.393 | 0.095 | 0.296 | |
| Zn_RAC | −0.168 | 0.172 | −0.179 | 0.156 | |
| TPb | −0.015 | 0.465 | 0.068 | 0.351 | |
| PbAcid | −0.225 | 0.101 | −0.235 | 0.090 | |
| PbReduc | 0.050 | 0.390 | 0.138 | 0.218 | |
| PbOxid | −0.002 | 0.496 | 0.165 | 0.176 | |
| Pb_RAC | −0.141 | 0.213 | −0.159 | 0.184 | |
| OC | 0.076 | 0.335 | 0.171 | 0.167 | |
| MPCT | Clay | 0.248 | 0.061 | 0.127 | 0.217 |
| Silt | −0.152 | 0.175 | 0.019 | 0.454 | |
| NiAcid | −0.479 | 0.001 ** | −0.417 | 0.007 * | |
| NiReduc | −0.273 | 0.088 | −0.200 | 0.108 | |
| NiOxid | −0.432 | 0.005 * | −0.284 | 0.076 | |
| Ni_RSP | −0.323 | 0.021 * | −0.113 | 0.244 | |
| TCu | −0.474 | 0.002 * | −0.304 | 0.056 | |
| CuReduc | −0.186 | 0.125 | −0.211 | 0.096 | |
| CuOxid | −0.372 | 0.009 * | −0.302 | 0.058 | |
| Cu_RSP | −0.332 | 0.018 * | −0.166 | 0.153 | |
| TZn | −0.510 | 0.001 ** | −0.382 | 0.015 * | |
| ZnAcid | −0.538 | 0.0001 ** | −0.355 | 0.024 * | |
| ZnReduc | −0.409 | 0.009 * | −0.331 | 0.037 * | |
| ZnOxid | −0.417 | 0.007 * | −0.260 | 0.053 | |
| Zn_RSP | −0.523 | 0.001 ** | −0.310 | 0.052 | |
| Zn_RAC | −0.311 | 0.052 | −0.103 | 0.264 | |
| PbOxid | 0.042 | 0.398 | −0.018 | 0.456 | |
| OC | −0.397 | 0.011 * | −0.242 | 0.066 | |
| MPFQ | Clay | −0.663 | 0.0001 ** | −0.582 | 0.002 * |
| Eh | −0.606 | 0.001 ** | −0.590 | 0.002 * | |
| Salinity | 0.527 | 0.006 * | 0.466 | 0.016 * | |
| TCr | 0.566 | 0.003 * | 0.479 | 0.013 * | |
| CrAcid | 0.549 | 0.004 * | 0.410 | 0.038 * | |
| CrReduc | 0.443 | 0.023 * | 0.420 | 0.033 * | |
| Cr_RSP | 0.439 | 0.025 * | 0.525 | 0.006 * | |
| Cr_RAC | 0.502 | 0.009 * | 0.329 | 0.101 | |
| TNi | 0.331 | 0.099 | 0.246 | 0.226 | |
| NiReduc | 0.364 | 0.067 | 0.340 | 0.089 | |
| NiOxid | 0.255 | 0.209 | 0.326 | 0.104 | |
| NiResid | −0.523 | 0.006 * | −0.550 | 0.004 * | |
| Ni_RSP | 0.330 | 0.100 | 0.412 | 0.037 * | |
| TCu | 0.357 | 0.073 | 0.422 | 0.032 * | |
| CuAcid | 0.195 | 0.340 | 0.132 | 0.521 | |
| CuOxid | 0.694 | 0.0001 ** | 0.704 | 0.0001 ** | |
| Cu_RSP | 0.225 | 0.268 | 0.174 | 0.395 | |
| Cu_RAC | 0.136 | 0.509 | 0.032 | 0.875 | |
| TZn | 0.350 | 0.080 | 0.415 | 0.035 * | |
| ZnAcid | 0.406 | 0.04 * | 0.368 | 0.065 | |
| ZnOxid | 0.233 | 0.253 | 0.230 | 0.259 | |
| ZnResid | −0.510 | 0.008 * | −0.442 | 0.024 * | |
| Zn_RSP | 0.450 | 0.021 * | 0.426 | 0.03 * | |
| Zn_RAC | 0.247 | 0.223 | 0.163 | 0.426 | |
| TPb | −0.308 | 0.125 | 0.140 | 0.495 | |
| OC | 0.702 | 0.0001 ** | 0.714 | 0.0001 ** | |
| Site | Env 1 | Env 2 | Explained % | ||
|---|---|---|---|---|---|
| Env 1 | Env 2 | Unexplained | |||
| GXJHW | Clay, Eh, Salinity, OC | TCr CrAcid CrReduc Cr_RSP Cr_RAC TNi NiReduc NiOxid NiResid Ni_RSP TCu CuAcid CuOxid Cu_RSP Cu_RAC TZn ZnAcid ZnOxid ZnResid Zn_RSP Zn_RAC TPb | 43.7 | 61.5 | 19.4 |
| GXSK | pH, Eh, Salinity, OC | TCr CrAcid CrReduc Cr_RAC NiReduc NiOxid Ni_RAC TCu CuResid Cu_RSP TZn ZnReduc ZnOxid ZnResid Zn_RSP Zn_RAC TPb PbAcid PbReduc PbOxid | 50.9 | 79.9 | 12.6 |
| MPCT | Clay, Silt, OC | NiAcid NiReduc NiOxid Ni_RSP TCu CuReduc CuOxid Cu_RSP TZn ZnAcid ZnReduc ZnOxid Zn_RSP Zn_RAC PbOxid | 27.3 | 71.8 | 25.0 |
| MPFQ | Clay, Eh, Salinity, OC | TCr CrAcid CrReduc Cr_RSP Cr_RAC TNi NiReduc NiOxid NiResid Ni_RSP TCu CuAcid CuOxid Cu_RSP Cu_RAC TZn ZnAcid ZnOxid ZnResid Zn_RSP Zn_RAC TPb | 43.7 | 91.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, B.; Liao, J.; Zhang, Q.; Ding, S.; He, M.; Qiao, Y.; Zhang, Z.; Shang, C.; Chen, S. Diversity and Vertical Distribution of Sedimentary Bacterial Communities and Its Association with Metal Bioavailability in Three Distinct Mangrove Reserves of South China. Water 2022, 14, 971. https://doi.org/10.3390/w14060971
Hu B, Liao J, Zhang Q, Ding S, He M, Qiao Y, Zhang Z, Shang C, Chen S. Diversity and Vertical Distribution of Sedimentary Bacterial Communities and Its Association with Metal Bioavailability in Three Distinct Mangrove Reserves of South China. Water. 2022; 14(6):971. https://doi.org/10.3390/w14060971
Chicago/Turabian StyleHu, Bochao, Junxian Liao, Qijiong Zhang, Suli Ding, Mengyuan He, Yue Qiao, Zuye Zhang, Chenjing Shang, and Si Chen. 2022. "Diversity and Vertical Distribution of Sedimentary Bacterial Communities and Its Association with Metal Bioavailability in Three Distinct Mangrove Reserves of South China" Water 14, no. 6: 971. https://doi.org/10.3390/w14060971
APA StyleHu, B., Liao, J., Zhang, Q., Ding, S., He, M., Qiao, Y., Zhang, Z., Shang, C., & Chen, S. (2022). Diversity and Vertical Distribution of Sedimentary Bacterial Communities and Its Association with Metal Bioavailability in Three Distinct Mangrove Reserves of South China. Water, 14(6), 971. https://doi.org/10.3390/w14060971