
Dietary Changes in the Ark Clam (Anadara kagoshimensis) Is Associated with Phytoplankton Community Patterns in a Temperate Coastal Embayment



Abstract
:1. Introduction
2. Materials and Methods
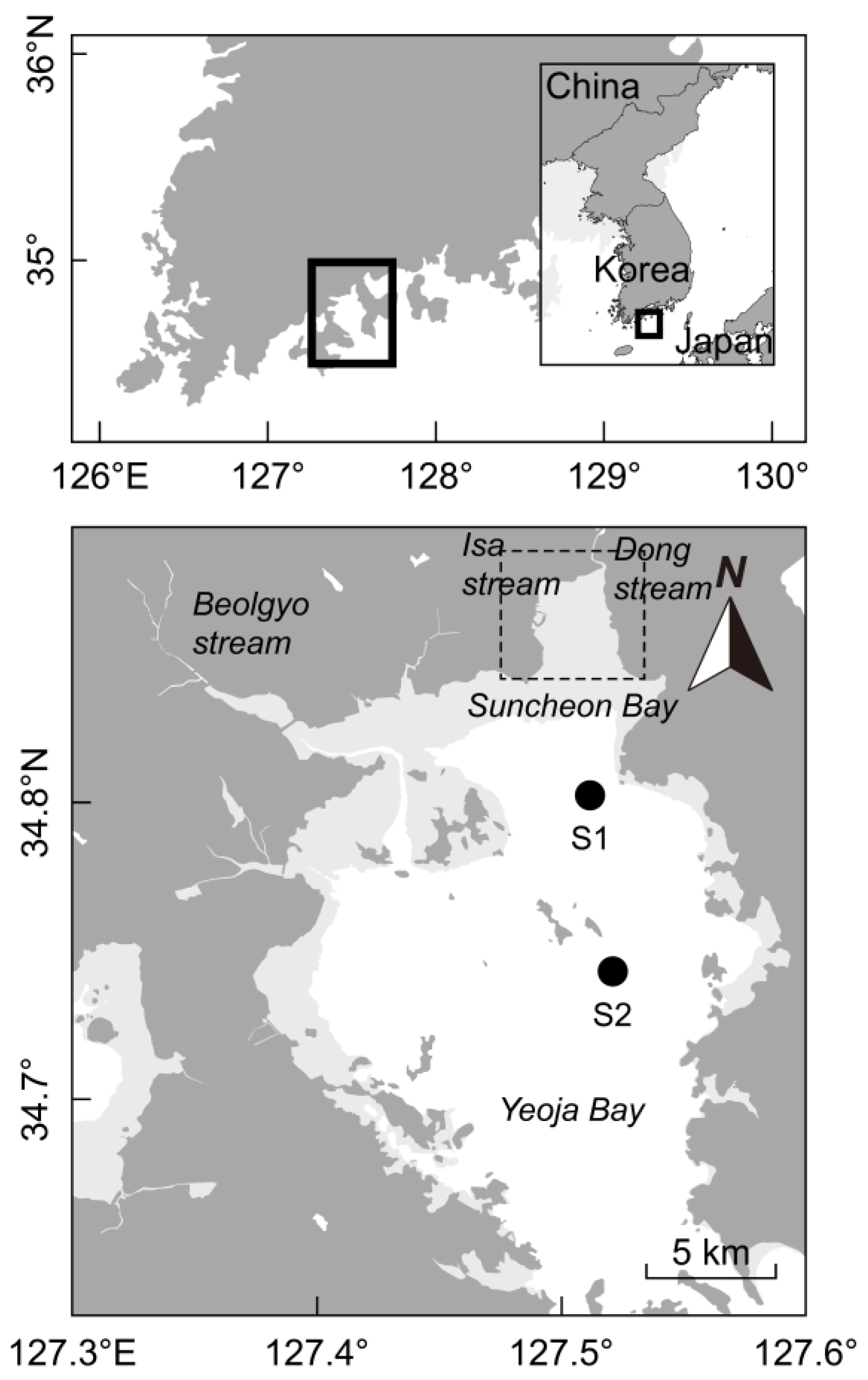
2.1. Site Description and Sampling Protocol
2.2. Collection and Treatment of Samples, and Analyses of Environmental Parameters
2.3. Pigment Analysis and Chemotaxonomic Identification of Phytoplankton
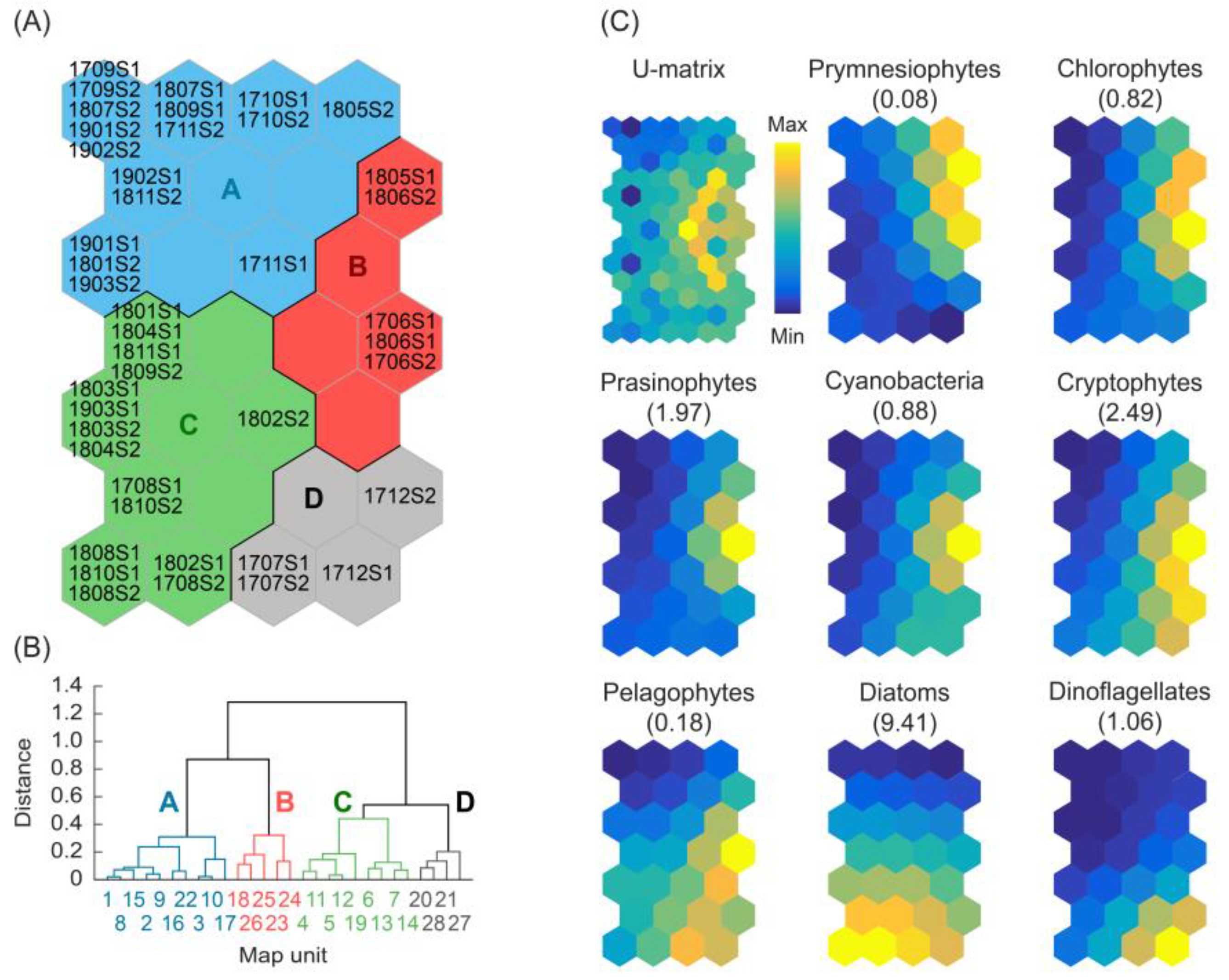
2.4. Self-Organizing Map Analysis and Clustering of the Phytoplankton Communities
2.5. Stable Isotope Analyses
2.6. Statistical Analyses
2.7. Isotope Mixing Model
3. Results
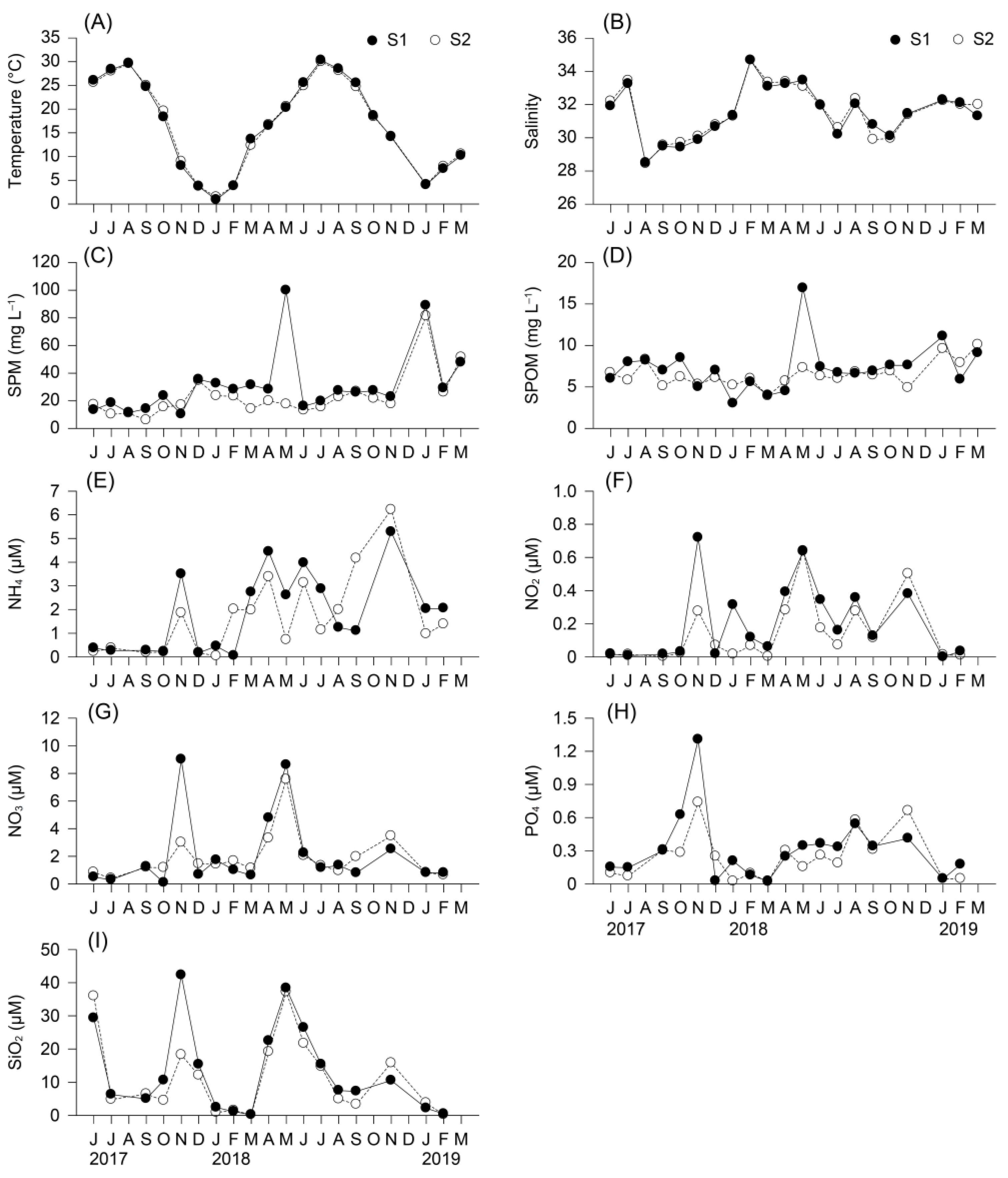
3.1. Environmental Conditions
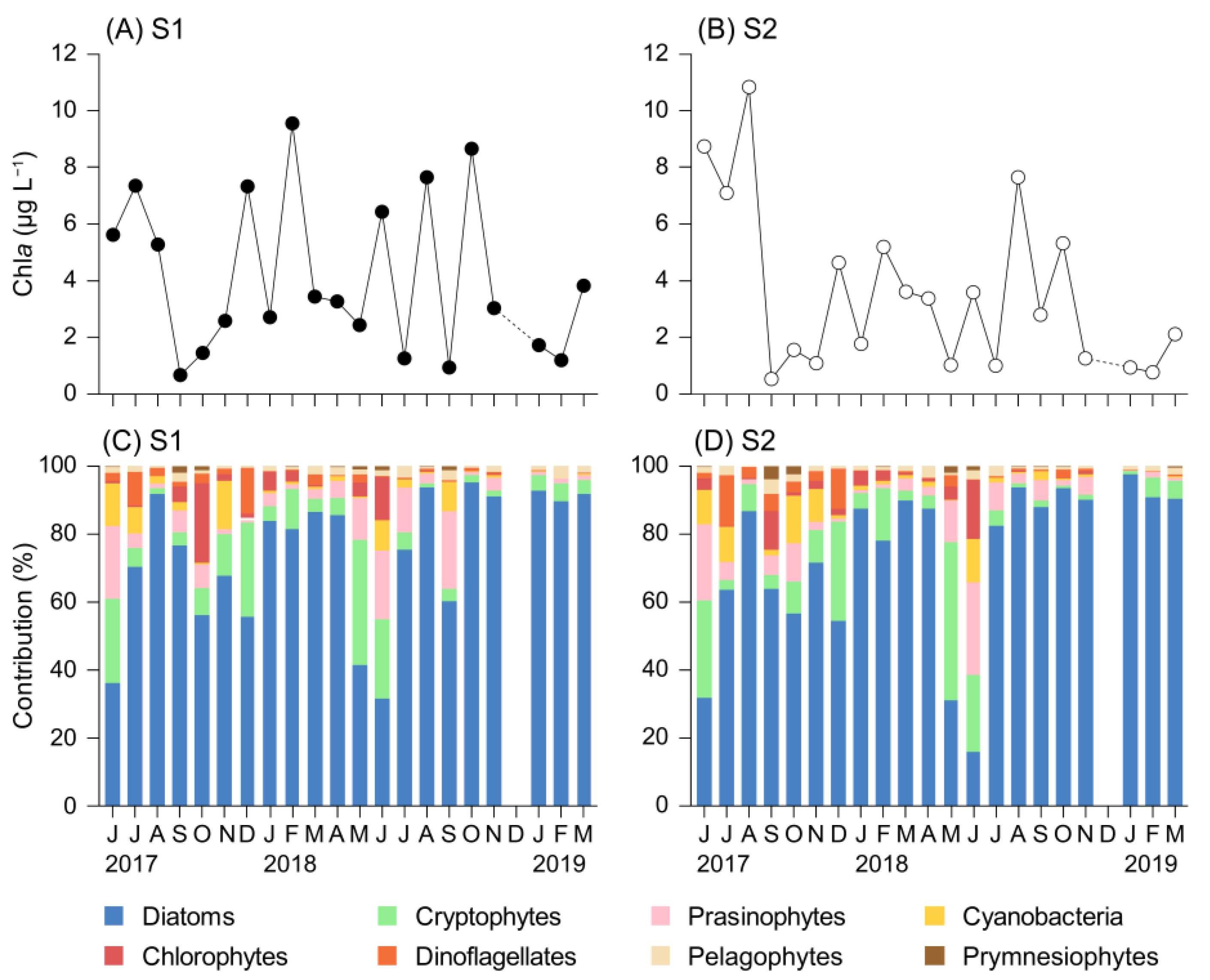
3.2. Chla Concentration and Phytoplankton Community Composition
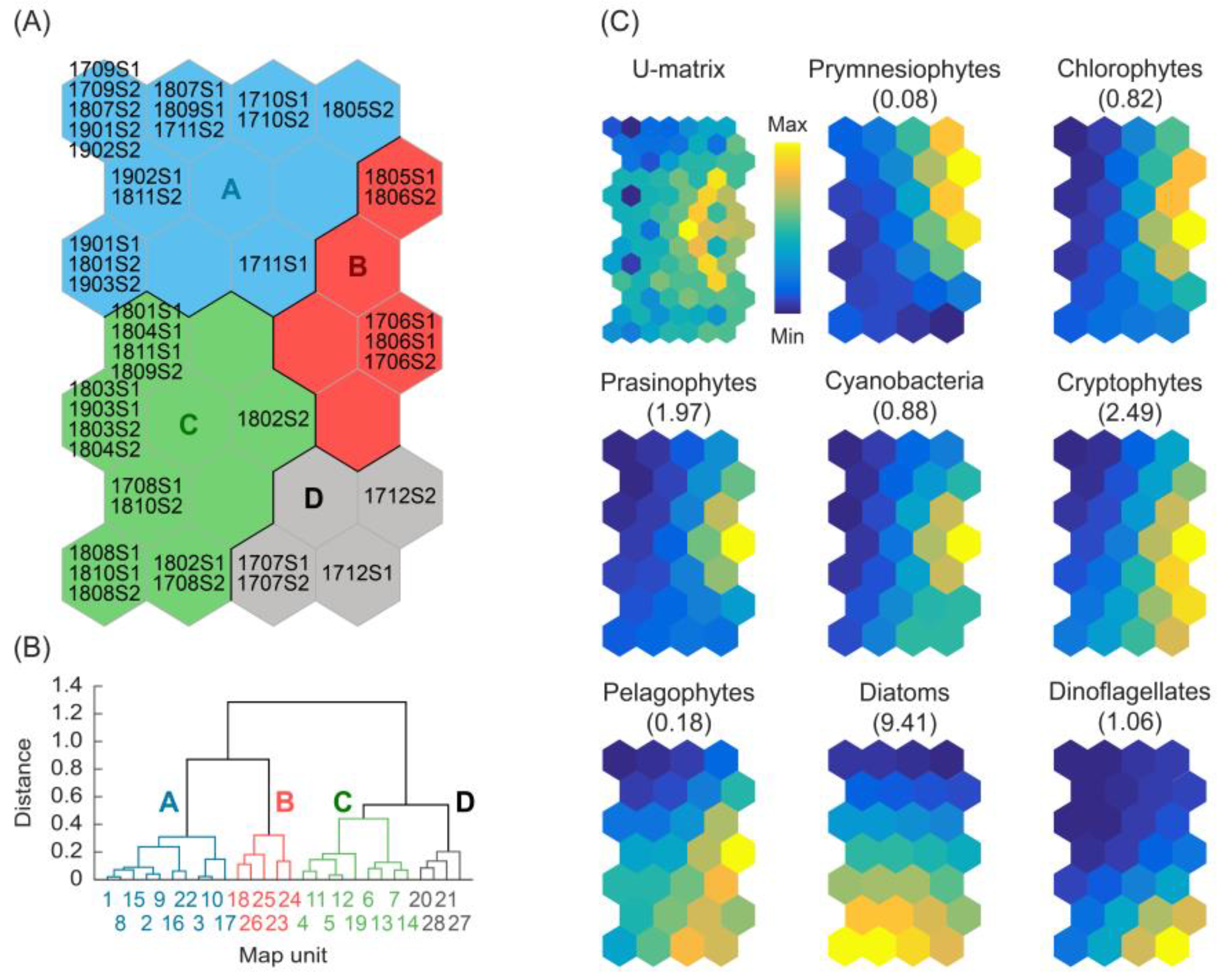
3.3. Patterning the Phytoplankton Communities
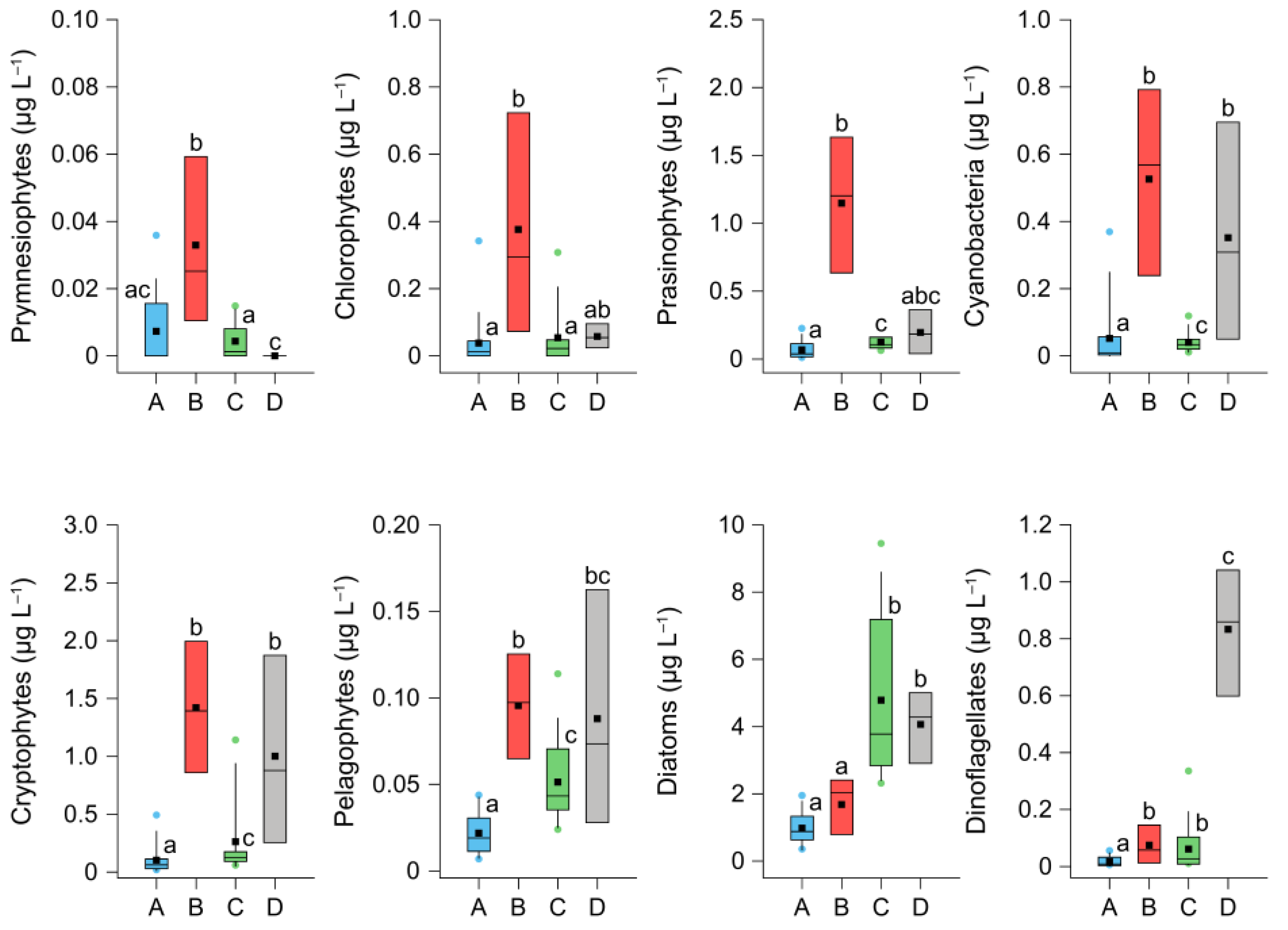
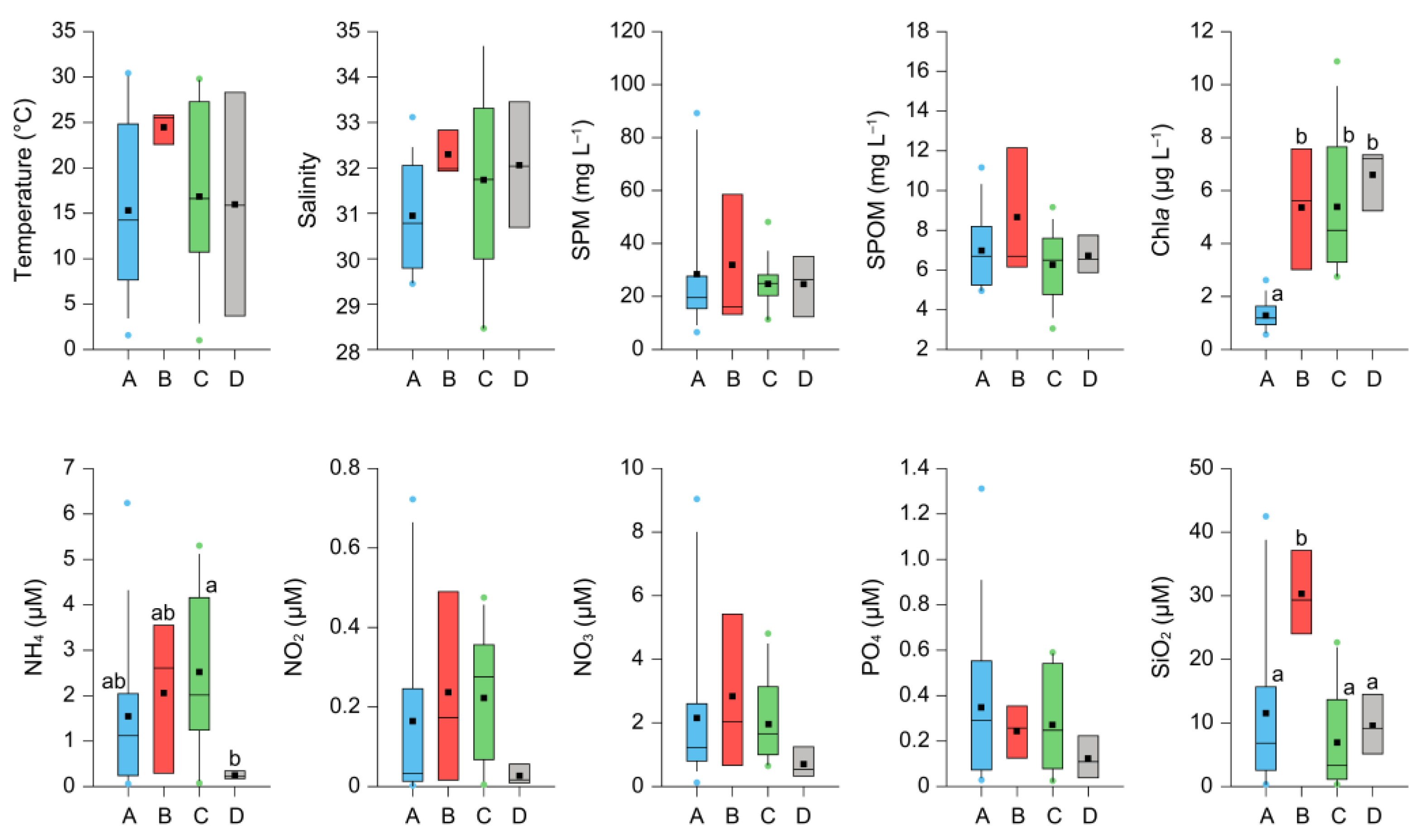
3.4. Characterization of the Environmental Conditions of Clusters
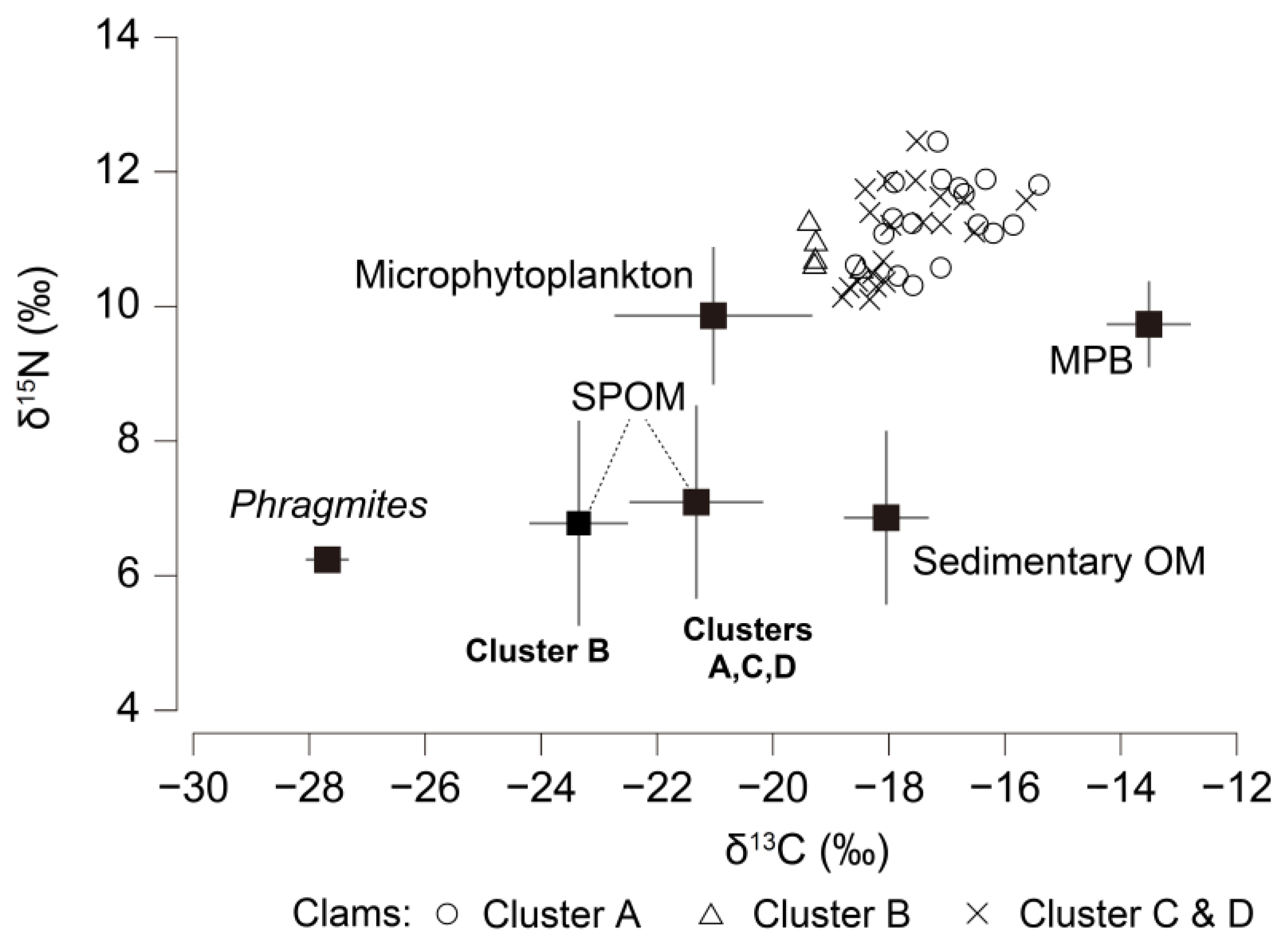
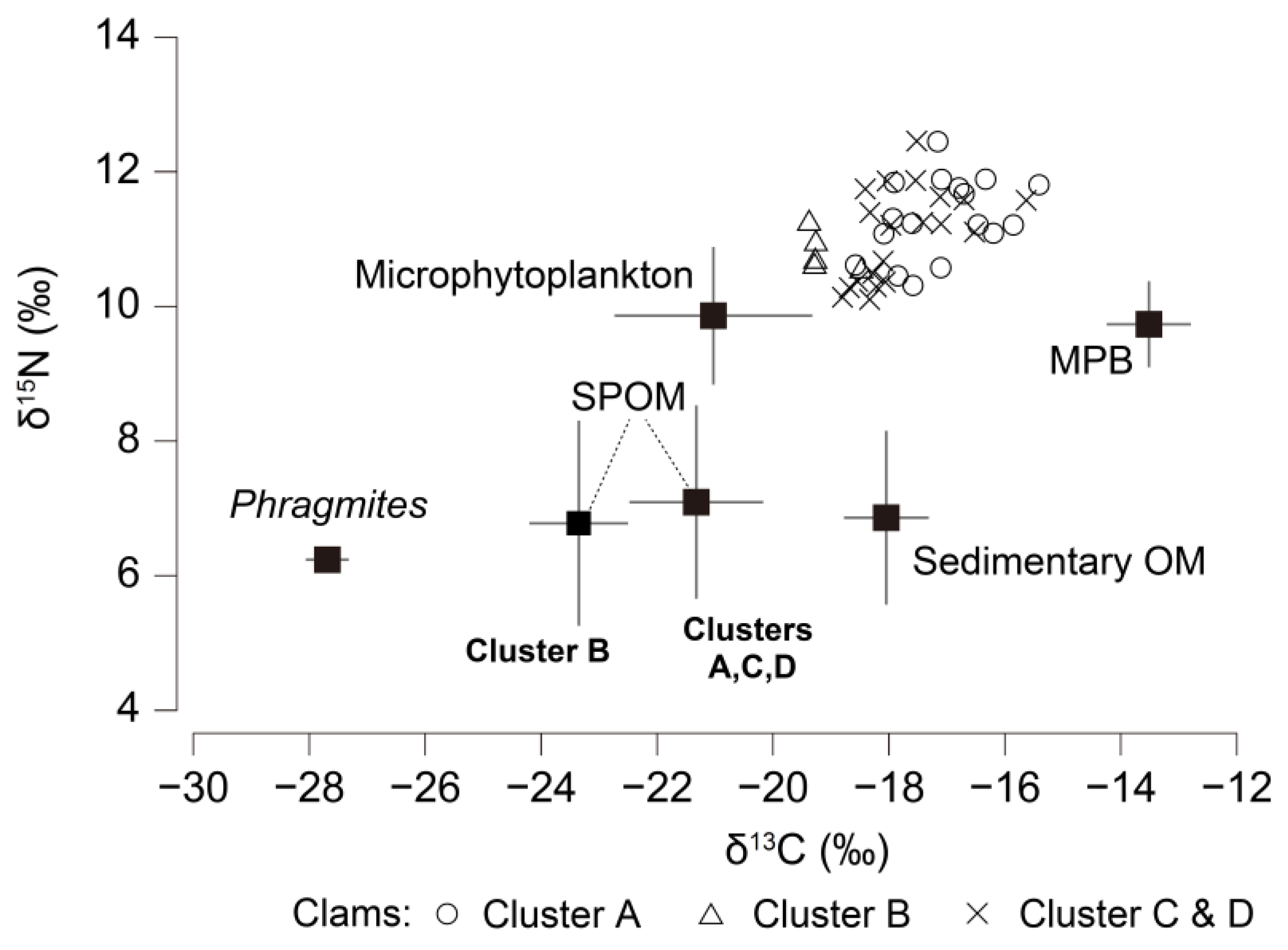
3.5. Isotope Signatures of Feasible Diet Components and Clams
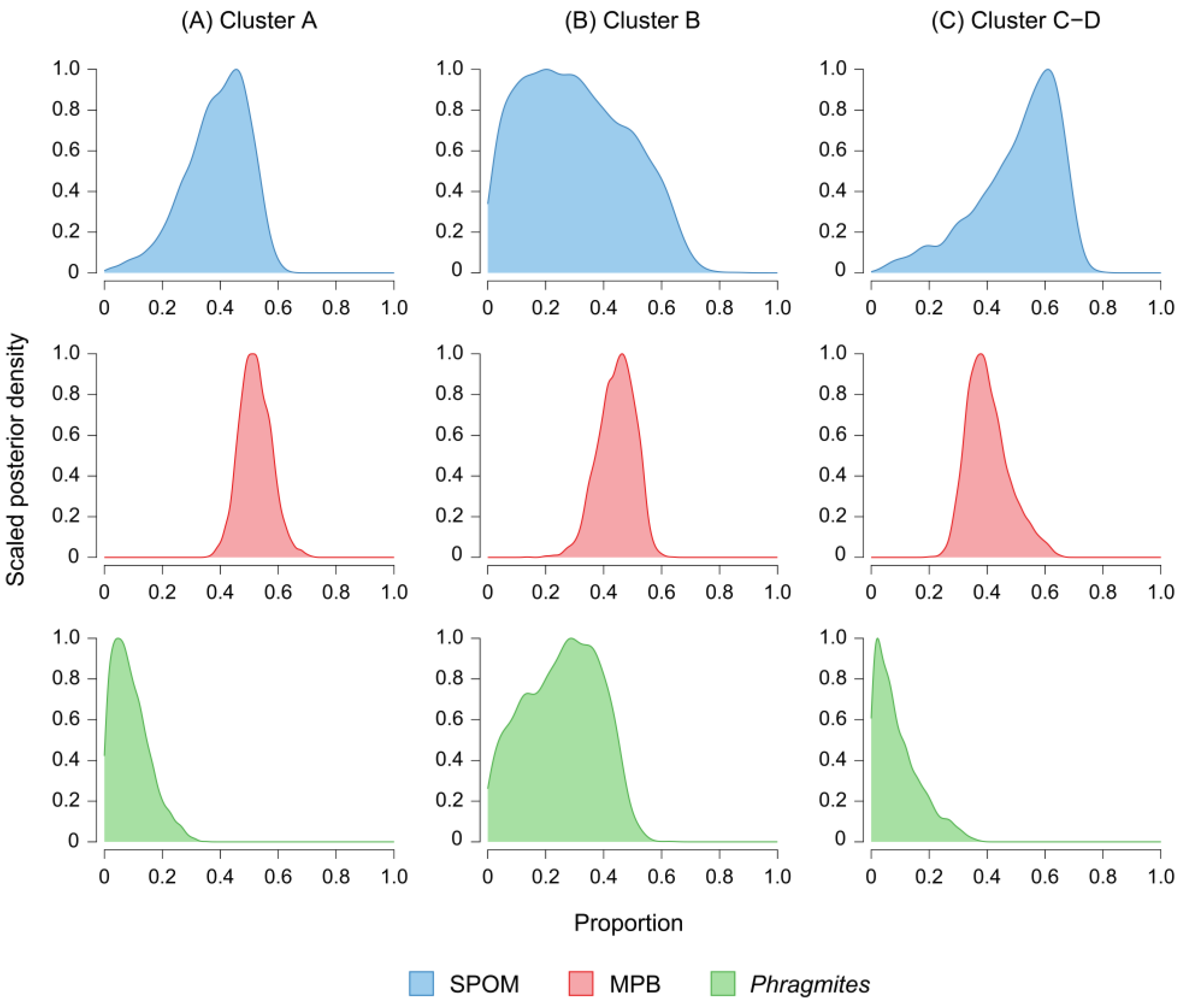
3.6. Contributions of Organic Matter Sources to Clam Nutrition
4. Discussion
4.1. Phytoplankton Community Patterns
4.2. Temporal Variation in the Contribution of Phytoplankton to Clam Nutrition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, M.; Ferentinos, G.; Banner, F.T. The hydrodynamics and sedimentology of a high (tidal and wave) energy embayment (Swansea Bay, northern Bristol Channel). Estuar. Coast. Mar. Sci. 1979, 8, 49–74. [Google Scholar] [CrossRef]
- Ke, X.; Evans, G.; Collins, M.B. Hydrodynamics and sediment dynamics of The Wash embayment, eastern England. Sedimentology 1996, 43, 157–174. [Google Scholar] [CrossRef]
- Duarte, P.; Alvarez-Salgado, X.A.; Fernández-Reiriz, M.J.; Piedracoba, S.; Labarta, U. A modeling study on the hydrodynamics of a coastal embayment occupied by mussel farms (Ria de Ares-Betanzos, NW Iberian Peninsula). Estuar. Coast. Shelf Sci. 2014, 147, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Millham, N.P.; Howes, B.L. Nutrient balance of a shallow coastal embayment: I. Patterns of groundwater discharge. Mar. Ecol. Prog. Ser. 1994, 112, 115–167. [Google Scholar] [CrossRef]
- Doval, M.D.; Álvarez-Salgado, X.A.; Pérez, F.F. Dissolved organic matter in a temperate embayment affected by coastal upwelling. Mar. Ecol. Prog. Ser. 1997, 157, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Barciela, R.M.; García, E.; Fernández, E. Modelling primary production in a coastal embayment affected by upwelling using dynamic ecosystem models and artificial neural networks. Ecol. Model. 1999, 120, 199–211. [Google Scholar] [CrossRef]
- Macintyre, H.L.; Geider, R.J.; Miller, D.C. Microphytobenthos: The Ecological role of the “secret garden” of unvegetated, shallow-water marine habitats. I. Distribution, abundance and primary production. Estuaries 1996, 19, 186–201. [Google Scholar] [CrossRef]
- Al-Maslamani, I.; Walton, M.E.M.; Kennedy, H.; Le Vay, L. Sources of primary production supporting food webs in an arid coastal embayment. Mar. Biol. 2012, 159, 1753–1762. [Google Scholar] [CrossRef]
- Akin, S.; Winemiller, K.O. Seasonal variation in food web composition and structure in a temperate tidal estuary. Estuar. Coast. 2006, 29, 552–567. [Google Scholar] [CrossRef]
- Christian, R.R.; Baird, D.; Luczkovich, J.; Johnson, J.C.; Scharler, U.M.; Ulanowicz, R.E. Role of network analysis in comparative ecosystem ecology of estuaries. In Aquatic Food Webs: An Ecosystem Approach; Belgrano, A., Scharler, U.M., Dunne, J., Ulanowicz, R., Eds.; Oxford Univ Press: Oxford, UK, 2005; pp. 25–40. [Google Scholar]
- Hagy, J.D.; Kemp, W.M. Estuarine food webs. In Estuarine Ecology, 2nd ed.; Day, J.W., Crump, B.C., Kemp, W.M., Yáñez-Arancibia, A., Eds.; Wiley-Blackwell: New York, NY, USA, 2013; pp. 417–441. [Google Scholar]
- Kang, C.K.; Lee, Y.W.; Choy, E.J.; Shin, J.K.; Seo, I.S.; Hong, J.S. Microphytobenthos seasonality determines growth and reproduction in intertidal bivalves. Mar. Ecol. Prog. Ser. 2006, 315, 113–127. [Google Scholar] [CrossRef]
- Kang, H.Y.; Seong, J.; Kim, C.; Lee, B.G.; Lee, I.T.; Kang, C.K. Seasonal energetic physiology in the ark shell Anadara kagoshimensis in response to rising temperature. Front. Mar. Sci. 2022, 9, 981504, in press. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, W.C.; Choy, E.J.; Choi, K.S.; Kang, C.K. Reproductive cycle and gross biochemical composition of the ark shell Scapharca subcrenata (Lischke, 1869) reared on subtidal mudflats in a temperate bay of Korea. Aquaculture 2011, 322–323, 149–157. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smtacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Cloern, J.E.; Dufford, R. Phytoplankton community ecology: Principles applied in San Francisco Bay. Mar. Ecol. Prog. Ser. 2005, 285, 11–28. [Google Scholar] [CrossRef]
- Garzke, J.; Connor, S.J.; Sommer, U.; O’Connor, M.I. Trophic interactions modify the temperature dependence of community biomass and ecosystem function. PLoS Biol. 2019, 17, e2006806. [Google Scholar] [CrossRef] [PubMed]
- Frelat, R.; Kortsch, S.; Kröncke, I.; Neumann, H.; Nordström, M.C.; Olivier, P.E.N.; Sell, A.F. Food web structure and community composition: A comparison across space and time in the North Sea. Ecography 2022, 2022, e05945. [Google Scholar] [CrossRef]
- Shumway, S.E.; Cucci, T.L.; Newell, R.C.; Yentsch, C.M. Particle selection, ingestion, and absorption in filter-feeding bivalves. J. Exp. Mar. Biol. Ecol. 1985, 91, 77–92. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Larsen, P.S. Comparative ecophysiology of active zoobenthic filter feeding, essence of current knowledge. J. Sea Res. 2000, 44, 169–193. [Google Scholar] [CrossRef]
- Bandelj, V.; Socal, G.; Park, Y.S.; Lek, S.; Coppola, J.; Camatti, E.; Capuzzo, E.; Milani, L.; Solidoro, C. Analysis of multitrophic plankton assemblages in the Lagoon of Venice. Mar. Ecol. Prog. Ser. 2008, 368, 23–40. [Google Scholar] [CrossRef]
- McMeans, B.C.; McCann, K.S.; Humphries, M.; Rooney, N.; Fisk, A.T. Food web structure in temporally-forced ecosystems. Trend. Ecol. Evol. 2015, 30, 662–672. [Google Scholar] [CrossRef]
- Kang, H.Y.; Kim, C.; Kim, D.; Lee, Y.J.; Park, H.J.; Kundu, G.K.; Kim, Y.K.; Bibi, R.; Jang, J.; Lee, K.H.; et al. Identifying patterns in the multitrophic community and food-web structure of a low-turbidity temperate estuarine bay. Sci. Rep. 2020, 10, 16637. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size-structured populations. Ann. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- D’Alelio, D.; Libralato, S.; Wyatt, T.; d’Alcalà, M.R. Ecological-network models link diversity, structure and function in the plankton food-web. Sci. Rep. 2016, 6, 21806. [Google Scholar] [CrossRef] [Green Version]
- Michener, R.H.; Kaufman, L. Stable Isotope Ratios as Tracers in Marine Food Webs: An Update in Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Michener, R., Lajtha, K., Eds.; Blackwell: New York, NY, USA, 2007; pp. 238–282. [Google Scholar]
- Fry, B.; Sherr, E. δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib. Mar. Sci. 1984, 27, 13–47. [Google Scholar]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Ann. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary consumer δ15N and δ13C and the trophic position of aquatic consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- Kang, C.K.; Kim, J.B.; Lee, K.S.; Kim, J.B.; Lee, P.Y.; Hong, J.S. Trophic importance of benthic microalgae to macrozoobenthos in coastal bay systems in Korea: Dual stable C and N isotope analyses. Mar. Ecol. Prog. Ser. 2003, 259, 79–92. [Google Scholar] [CrossRef]
- Kasai, A.; Toyohara, H.; Nakata, A.; Miura, T.; Azuma, N. Food sources for the bivalve Corbicula japonica in the foremost fishing lakes estimated from stable isotope analysis. Fish. Sci. 2006, 72, 105–114. [Google Scholar] [CrossRef]
- Decottignies, P.; Beninger, P.G.; Rincé, Y.; Robins, R.J.; Riera, P. Exploitation of natural food sources by two sympatric, invasive suspension-feeders: Crassostrea gigas and Crepidula fornicata. Mar. Ecol. Prog. Ser. 2007, 334, 179–192. [Google Scholar] [CrossRef]
- Prins, T.C.; Smaal, A.C.; Dame, R.F. A review of the feedbacks between bivalve grazing and ecosystem processes. Aquat. Ecol. 1997, 31, 349–359. [Google Scholar] [CrossRef]
- Murphy, A.E.; Emery, K.A.; Anderson, I.C.; Pace, M.L.; Brush, M.J.; Rheuban, J.E. Quantifying the effects of commercial clam aquaculture on C and N cycling: An integrated ecosystem approach. Estuar. Coast. 2016, 39, 1746–1761. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-J.; Han, E.; Wilberg, M.J.; Lee, W.C.; Choi, K.-S.; Kang, C.K. Physiological processes and gross energy budget of the submerged longline-cultured Pacific oyster Crassostrea gigas in a temperate bay of Korea. PLoS ONE 2018, 13, e0199752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.K.; Choy, E.J.; Hur, Y.B.; Myoung, J.I. Isotopic evidence of particle size dependent food partitioning in cocultured suspension feeders, the sea squirt Halocynthia roretzi and the Pacific oyster Crassostrea gigas. Aqua. Biol. 2009, 6, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Riera, P.; Stal, L.J.; Nieuwenhuiz, J.; Richard, P.; Blanchard, G.; Gentil, F. Determination of food sources for benthic invertebrates in a salt marsh (Aiguillon Bay, France) by carbon and nitrogen stable isotopes: Importance of locally produced sources. Mar. Ecol. Prog. Ser. 1999, 187, 301–307. [Google Scholar] [CrossRef]
- Lee, M.O.; Kim, J.K.; Kim, B.K. Marine environmental characteristics of Yeoja Bay, Korea—A review on a basis of previous studies. J. Korean Soc. Mar. Environ. Energy 2020, 23, 233–245. (In Korean) [Google Scholar] [CrossRef]
- Choi, M.; Kim, H.C.; Hwang, D.W.; Lee, I.S.; Kim, Y.S.; Kim, Y.J.; Choi, H.G. Organic enrichment and pollution in surface sediments from shellfish farming in Yeoja Bay and Gangjin Bay, Korea. Kor. J. Fish. Aquat. Sci. 2013, 46, 424–436. (In Korean) [Google Scholar]
- Hwang, D.W.; Kim, G.; Lee, Y.W.; Yang, H.S. Estimating submarine inputs of groundwater and nutrients to a coastal bay using radium isotopes. Mar. Chem. 2005, 96, 61–71. [Google Scholar] [CrossRef]
- Korean Statistical Information Service. KOSIS. Available online: https://kosis.kr (accessed on 1 April 2022).
- Murphy, J.; Riley, J.P. A Modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Helder, W.; de Vries, R.T.P. An automatic phenol-hypochlorite method for the determination of ammonia in sea and brackish water. Neth. J. Sea Res. 1979, 13, 154–160. [Google Scholar] [CrossRef]
- Hansen, H.P.; Grasshoff, K. Automated chemical analysis. In Methods of Seawater Analysis; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 347–379. [Google Scholar]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W. Application of pigment methods to oceanography. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffery, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 127–166. [Google Scholar]
- Park, H.J.; Kwak, J.H.; Kang, H.Y.; Kwon, K.-Y.; Lim, W.; Kang, C.K. Incorporation of Cochlodinium bloom-derived organic matter into a temperate subtidal macrobenthic food web as traced by stable isotopes. Mar. Pollut. Bull. 2020, 154, 111053. [Google Scholar] [CrossRef] [PubMed]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.W.; van den Enden, R.L. Phytoplankton community structure and stocks in the East Antarctic marginal ice zone (BROKE survey, January–March 1996) determined by CHEMTAX analysis of HPLC pigment signatures. Deep-Sea Res. II 2020, 47, 2363–2400. [Google Scholar] [CrossRef]
- Lee, Y.W.; Park, M.O.; Kim, Y.S.; Kim, S.S.; Kang, C.K. Application of photosynthetic pigment analysis using a HPLC and CHEMTAX program to studies of phytoplankton community composition. J. Korean Soc. Oceanogr. 2011, 16, 117–124. [Google Scholar]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organizing Maps, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Murtagh, F.; Hernández-Pajares, M. The Kohonen self-organizing map method: An assessment. J. Classif. 1995, 12, 165–190. [Google Scholar] [CrossRef]
- Park, Y.S.; Tison, J.; Lek, S.; Giraudel, J.L.; Coste, M.; Delmas, F. Application of a self-organizing map to select representative species in multivariate analysis: A case study determining diatom distribution patterns across France. Ecol. Inform. 2006, 1, 247–257. [Google Scholar] [CrossRef]
- Vesanto, J.; Alhoniemi, E. Clustering of the self-organizing map. IEEE Trans. Neural Netw. 2000, 11, 586–600. [Google Scholar] [CrossRef]
- Alhoniemi, E.; Himberg, J.; Parhankangas, J.; Vesanto, J. SOM Toolbox. 2000. Available online: http://www.cis.hut.fi/projects/somtoolbox (accessed on 18 April 2022).
- Park, H.J.; Kang, H.Y.; Park, T.H.; Kang, C.K. Comparative trophic structures of macrobenthic food web in two macrotidal wetlands with and without a dike on the temperate coast of Korea as revealed by stable isotopes. Mar. Environ. Res. 2017, 131, 134–145. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for Primer; Primer-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- De Cáceres, M.; Jansen, F. Studying the Statistical Relationship between Species and Groups of Sites. R Package: Indicspecies, Version 1.7.6. 2016. Available online: http://cran.r-project.org (accessed on 1 June 2022).
- R Development Core Team. R: A Language and Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Stock, B.C.; Semmens, B.X. MixSIAR GUI User Manual, Version 3.1. 2016. Available online: https://github.com/brianstock/MixSIAR (accessed on 5 October 2022).
- Yokoyama, H.; Tamaki, A.; Harada, K.; Shimoda, K.; Koyama, K.; Ishihi, Y. Variability of diet-tissue isotopic fractionation in estuarine macrobenthos. Mar. Ecol. Prog. Ser. 2005, 296, 115–128. [Google Scholar] [CrossRef]
- Wang, X.H. Tide-induced sediment resuspension and the bottom boundary layer in an idealized estuary with a muddy bed. J. Phys. Oceanogr. 2002, 32, 3113–3131. [Google Scholar] [CrossRef]
- Whipple, A.C.; Luettich, R.A.; Reynolds-Fleming, J.V.; Neve, R.H. Spatial differences in wind-driven sediment resuspension in a shallow, coastal estuary. Estuar. Coast. Shelf Sci. 2018, 213, 49–60. [Google Scholar] [CrossRef]
- Lee, D.I.; Choi, J.M.; Lee, Y.G.; Lee, M.O.; Lee, W.C.; Kim, J.K. Coastal environmental assessment and management by ecological simulation in Yeoja Bay, Korea. Estuar. Coast. Shelf Sci. 2008, 80, 495–508. [Google Scholar] [CrossRef]
- Lipschultz, F. Nitrogen-specific uptake rates of marine phytoplankton isolated from natural populations of particles by flow cytometry. Mar. Ecol. Prog. Ser. 1995, 123, 245–258. [Google Scholar] [CrossRef]
- Bibi, R.; Kang, H.Y.; Kim, D.; Jang, J.; Kundu, G.K.; Kim, Y.K.; Kang, C.K. Dominance of autochthonous phytoplankton-derived particulate organic matter in a low-turbidity temperate estuarine embayment, Gwangyang Bay, Korea. Front. Mar. Sci. 2020, 7, 580260. [Google Scholar]
- Bayne, B.L.; Newell, R.C. Physiological energetics of marine Mollusca. In The Mollusca; Saleuddin, A.S.M., Wilbur, K.M., Eds.; Academic Press: New York, NY, USA, 1983; Volume 4, pp. 407–515. [Google Scholar]
- Gosling, E. Marine Bivalve Molluscs, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar]
- Kang, H.Y.; Lee, Y.J.; Choi, K.S.; Park, H.J.; Yun, S.G.; Kang, C.K. Combined effects of temperature and seston concentration on the physiological energetics of the Manila clam Ruditapes Philippinarum. PLoS ONE 2016, 11, e0152427. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Choy, E.J.; Kang, C.K. Spatial and temporal variations of microphytobenthos on an intertidal bed in a marine protected area of the Yeoja Bay, Korea. Wetlands 2013, 33, 737–745. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | MS | Pseudo-F | p (perm) |
|---|---|---|---|---|
| Cluster | 3 | 8537.8 | 20.02 | <0.001 |
| Residual | 38 | 426.51 | ||
| Total | 41 | |||
| Groups | t | p (perm) | ||
| A vs. B | 4.271 | <0.001 | ||
| A vs. C | 5.427 | <0.001 | ||
| A vs. D | 4.072 | <0.001 | ||
| B vs. C | 5.152 | <0.001 | ||
| B vs. D | 2.523 | 0.006 | ||
| C vs. D | 2.951 | <0.001 | ||
| Cluster | Group | IndVal | p |
|---|---|---|---|
| Cluster B | Prymnesiophytes | 0.86 | <0.001 |
| Chlorophytes | 0.82 | 0.002 | |
| Prasinophytes | 0.82 | 0.002 | |
| Cluster C | Diatoms | 0.59 | 0.049 |
| Cluster D | Dinoflagellates | 0.90 | <0.001 |
| Variable | F | df | p | Post Hoc Test | |
|---|---|---|---|---|---|
| Comparison | p | ||||
| δ13C | 46.216 | 2, 89 | <0.001 | PHY vs. SPOM | 0.206 |
| PHY vs. Sedimentary OM | <0.001 | ||||
| SPOM vs. Sedimentary OM | <0.001 | ||||
| δ15N | 53.282 | 2, 89 | <0.001 | PHY vs. SPOM | <0.001 |
| PHY vs. Sedimentary OM | <0.001 | ||||
| SPOM vs. Sedimentary OM | 0.848 | ||||
| A | B | C & D | Mean ± SD | ||
| δ13C | |||||
| PHY | −21.9 ± 1.4 (12) | −21.3 ± 2.1 (5) | −21.0 ± 1.9 (15) | −21.0 ± 1.7 (32) | |
| SPOM | −21.6 ± 1.3 a (15) | −23.3 ± 0.9 b (5) | −21.1 ± 1.0 a (18) | −21.6 ± 1.3 (38) | |
| Sedimentary OM | −18.4 ± 0.4 (5) | −17.6 ± 0.9 (3) | −18.0 ± 0.8 (12) | −18.0 ± 0.7 (20) | |
| MPB | −13.5 ± 0.7 (12) * | ||||
| Phragmites | −27.7 ± 0.4 (12) * | ||||
| δ15N | |||||
| PHY | 9.7 ± 1.3 (12) | 10.5 ± 0.8 (5) | 9.8 ± 0.7 (15) | 9.9 ± 1.0 (12) | |
| SPOM | 6.8 ± 1.7 (15) | 6.8 ± 1.5 (5) | 7.4 ± 1.1 (18) | 7.1 ± 1.4 (38) | |
| Sedimentary OM | 7.1 ± 1.1 (5) | 6.1 ± 0.4 (3) | 6.9 ± 1.5 (12) | 6.9 ± 1.3 (20) | |
| MPB | 9.7 ± 0.6 (12) * | ||||
| Phragmites | 6.2 ± 0.2 (12) * | ||||
| Variable | F | df | p | ||
|---|---|---|---|---|---|
| δ13C | 13.016 | 2, 39 | < 0.001 | ||
| δ15N | 1.433 | 2, 39 | 0.251 | ||
| A | B | C & D | Mean ± SD | ||
| δ13C | |||||
| Ark shell | −17.1 ± 0.9 a (17) | −19.1 ±0.4 b (5) | −17.8 ± 0.8 c (20) | −17.6 ± 1.0 (42) | |
| δ15N | |||||
| Ark shell | 11.3 ± 0.6 (17) | 10.8 ± 0.3 (5) | 11.1 ± 0.7 (20) | 11.1 ± 0.6 (42) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.Y.; Kim, C.; Kim, D.; Kwon, K.-Y.; Lee, W.C.; Kang, C.-K. Dietary Changes in the Ark Clam (Anadara kagoshimensis) Is Associated with Phytoplankton Community Patterns in a Temperate Coastal Embayment. Water 2022, 14, 3497. https://doi.org/10.3390/w14213497
Kang HY, Kim C, Kim D, Kwon K-Y, Lee WC, Kang C-K. Dietary Changes in the Ark Clam (Anadara kagoshimensis) Is Associated with Phytoplankton Community Patterns in a Temperate Coastal Embayment. Water. 2022; 14(21):3497. https://doi.org/10.3390/w14213497
Chicago/Turabian StyleKang, Hee Yoon, Changseong Kim, Dongyoung Kim, Kee-Young Kwon, Won Chan Lee, and Chang-Keun Kang. 2022. "Dietary Changes in the Ark Clam (Anadara kagoshimensis) Is Associated with Phytoplankton Community Patterns in a Temperate Coastal Embayment" Water 14, no. 21: 3497. https://doi.org/10.3390/w14213497