
Applying Electrical Resistivity Tomography and Biological Methods to Assess the Hyporheic Zone Water Exchanges in Two Mediterranean Stream Reaches

 ,
,  ,
,  ,
, 
Abstract
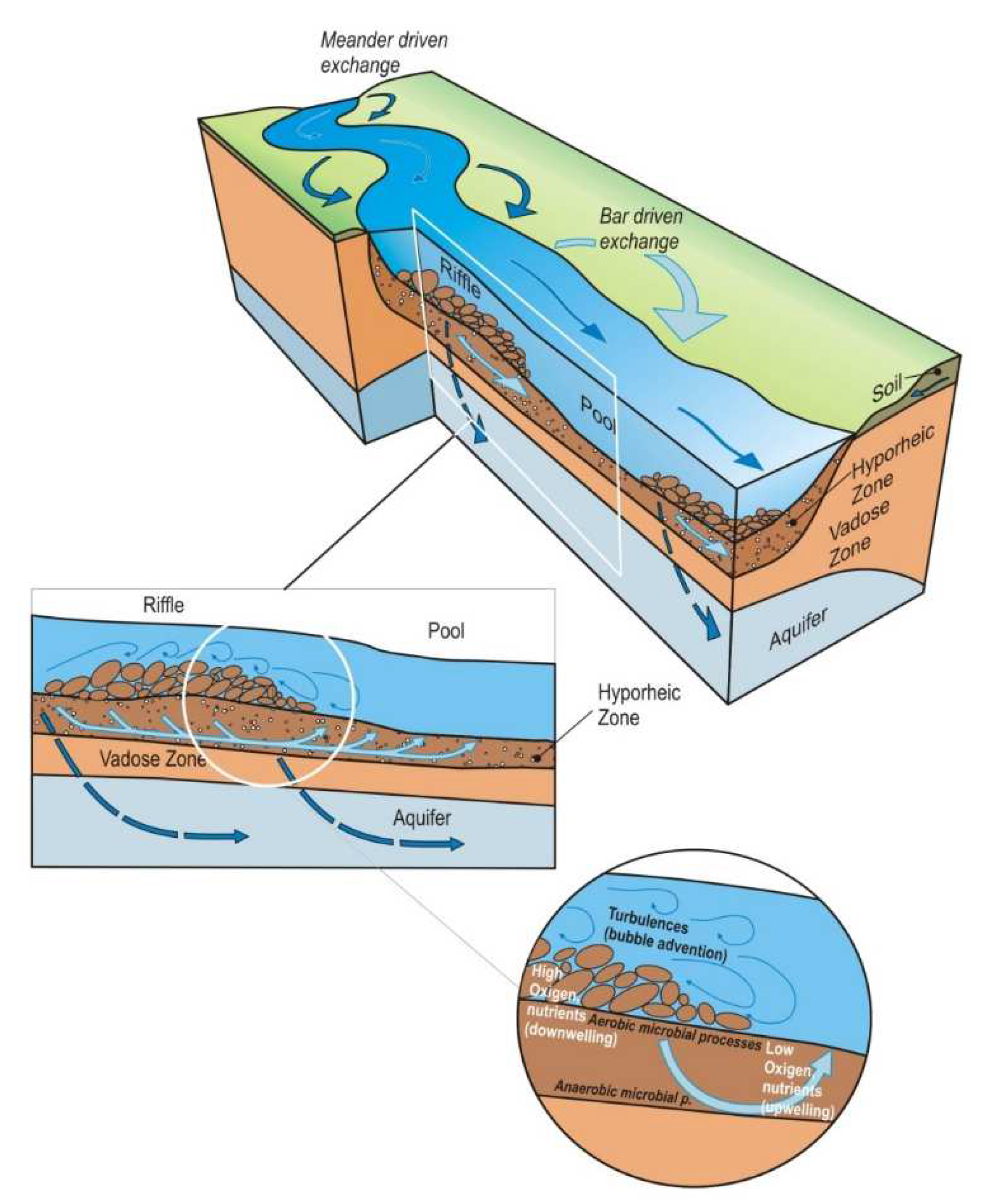
1. Introduction
2. Materials and Methods
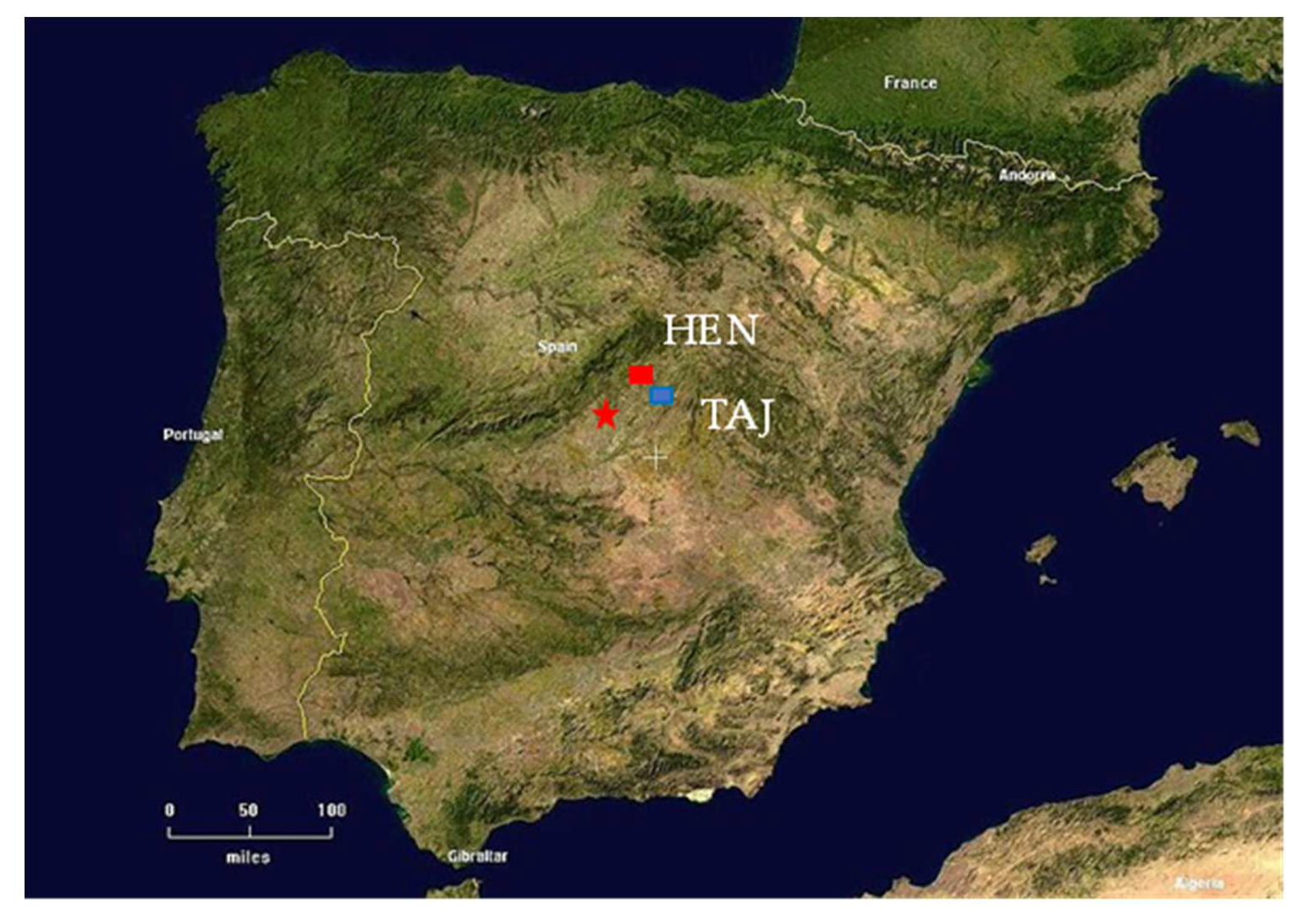
2.1. Study Sites and Sampling Strategy
2.2. Electrical Resistivity Measurements
2.3. Hyporheic Invertebrate Sampling and Community Characterisation
2.4. Data Analysis
3. Results
3.1. Environmental Conditions during the Experiments
3.2. Hyporheic Biota Assemblage’s Composition
3.3. Hyporheic Water Chemistry vs. Biota Relationships
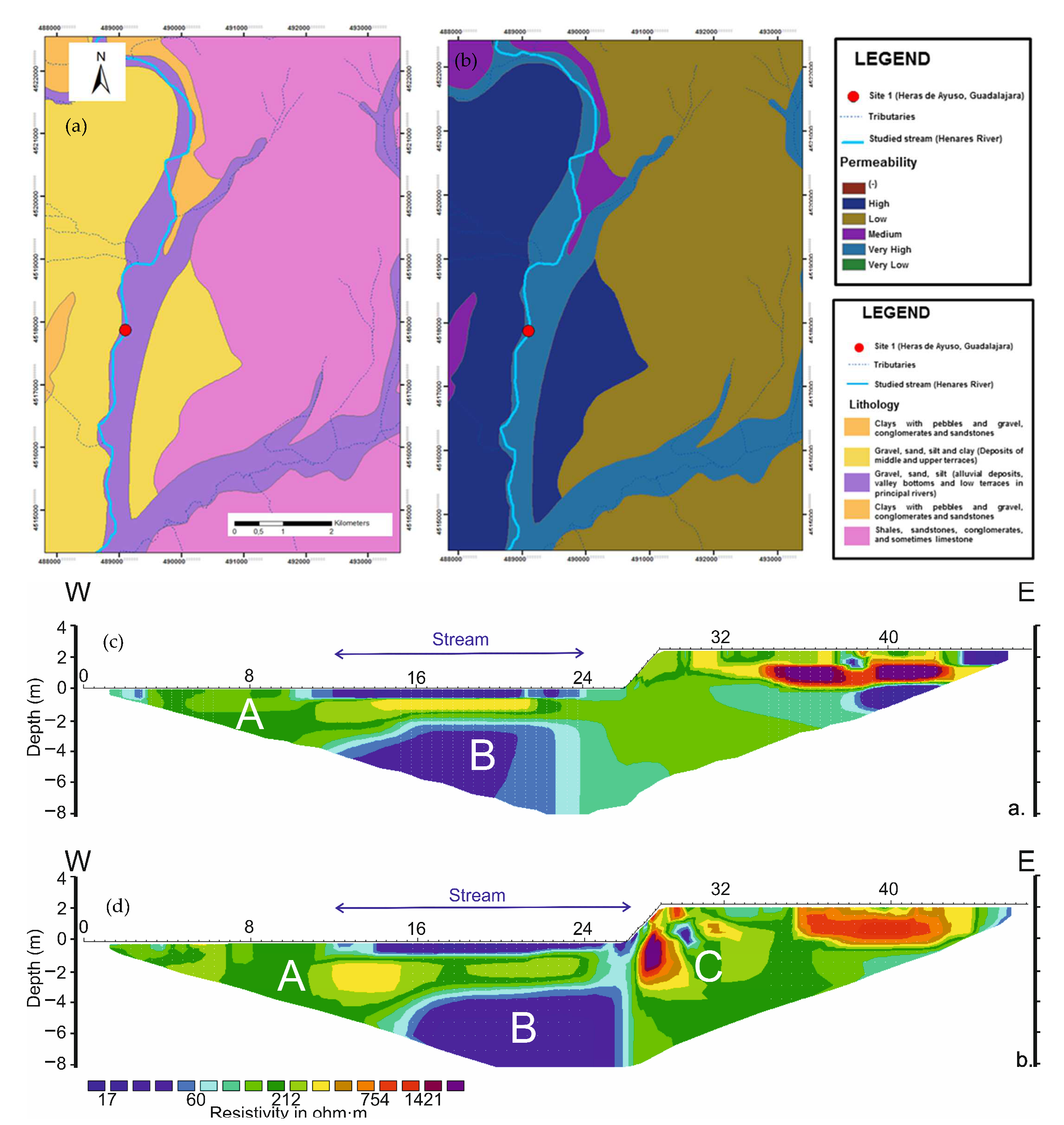
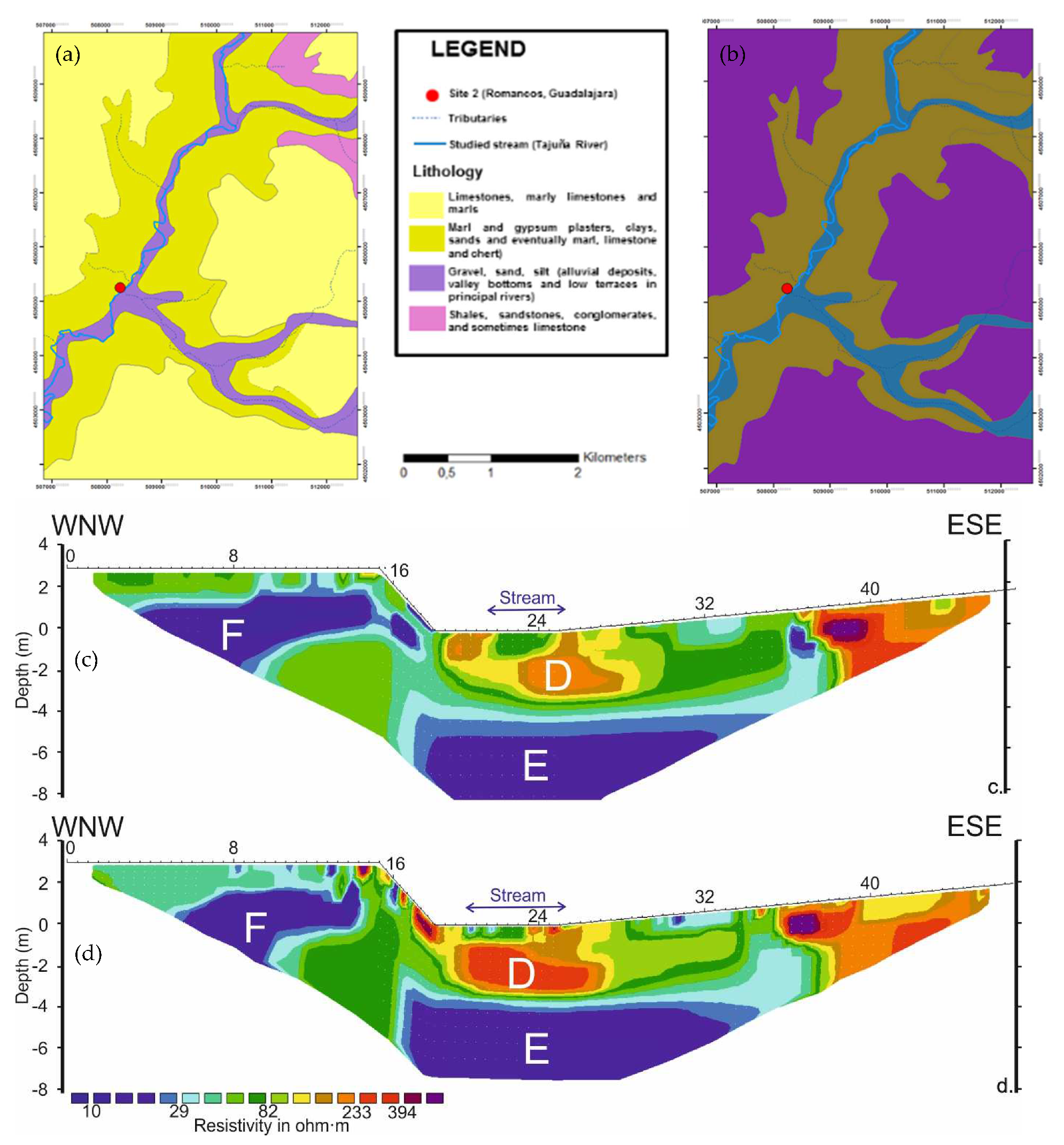
3.4. ERT Profiles
4. Discussion
Implication of Hyporheic Zone Biota as Indicators for Surface–Groundwater Exchanges: From Theory to Practice
5. Conclusions
Supplementary Materials
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orghidan, T. Un nou domeniu de viata acvatica subterana: ‘biotopul hiporeic’. Bul. Stiintific Sect. Biol. Stiinte Agric. 1955, 7, 657–676. [Google Scholar]
- Bencala, K.E. Hyporheic zone hydrological processes. Hydrol. Process. 2000, 14, 2797–2798. [Google Scholar] [CrossRef]
- Braun, A.; Auerswald, K.; Geist, J. Drivers and Spatio-Temporal Extent of Hyporheic Patch Variation: Implications for Sampling. PLoS ONE 2012, 7, e42046. [Google Scholar] [CrossRef] [PubMed]
- Leigh, C.; Stubbington, R.; Sheldon, F.; Boulton, A.J. Hyporheic invertebrates as bioindicators of ecological health in temporary rivers: A meta-analysis. Ecol. Indic. 2013, 32, 62–73. [Google Scholar] [CrossRef]
- Gibert, J.; Dole-Olivier, M.-J.; Marmonier, P.; Vervier, P. Surface water-groundwater ecotones. In he Ecology and Management of Aquatic-Terrestrial Ecotones; Naiman, R.J., Decamps, H., Eds.; UNESCO Parthenon Publishing: London, UK, 1990. [Google Scholar]
- Boulton, A.J.; Datry, T.; Kasahara, T.; Mutz, M.; Stanford, J.A. Ecology and management of the hyporheic zone: Stream-groundwater interactions. J. N. Am. Benthol. Soc. 2010, 29, 26–40. [Google Scholar] [CrossRef]
- Triska, F.J.; Kennedy, V.C.; Avanzino, R.J.; Zellweger, G.W.; Bencala, K.E. Retention and transport of nutrients in a third-order stream in northwestern California: Hyporheic processes. Ecology 1989, 70, 1893–1905. [Google Scholar] [CrossRef]
- Kautz, R.; Kawula, R.; Hoctor, T.; Comiskey, J.; Jansen, D.; Jennings, D.; Kasbohm, J.; Mazzotti, F.; McBride, R.; Richardson, L.; et al. How much is enough? Landscape-scale conservation for the Florida panther. Biol. Conserv. 2006, 130, 118–133. [Google Scholar] [CrossRef]
- Krause, S.; Hannah, D.M.; Fleckenstein, J.H.; Heppell, C.M.; Pickup, R.; Pinay, G.; Robertson, A.; Wood, P.J.; Kaeser, D. Inter-disciplinary perspectives on processes in the hyporheic zone. Ecohydrology 2011, 4, 481–499. [Google Scholar] [CrossRef]
- McGarr, J.T.; Wallace, C.D.; Ntarlagiannis, D.; Sturmer, D.M.; Soltanian, M.R. Geophysical mapping of hyporheic processes controlled by sedimentary architecture within compound bar deposits. Hydrol. Process. 2021, 35, e14358. [Google Scholar] [CrossRef]
- Palmer, M.A.; Ambrose, R.F.; Poff, N.L. Ecological Theory and Community Restoration Ecology. Restor. Ecol. 1997, 5, 291–300. [Google Scholar] [CrossRef]
- Boulton, A.J.; Findlay, S.; Marmonier, P.; Stanley, E.H.; Valett, H.M. The functional significance of the hyporheic zone in streams and rivers. Annu. Rev. Ecol.Syst. 1998, 29, 59–81. [Google Scholar] [CrossRef]
- Malcolm, I.A.; Soulsby, C.; Youngson, A.F.; Petry, J. Heterogeneity in ground water–surface water interactions in the hyporheic zone of a salmonid spawning ground. Hydrol. Process. 2004, 17, 601–617. [Google Scholar] [CrossRef]
- Stonedahl, S.H.; Harvey, J.W.; Harvey, J.W.; Salehin, M.; Packman, A.I. A multiscale model for integrat-ing hyporheic exchange from ripples to meanders. Water Resour. Res. 2010, 46. [Google Scholar] [CrossRef]
- Stubbington, R. The Hyporheic Zone as A Refugium for Benthic Invertebrates in 589 Groundwater-Dominated Streams. Ph.D. Thesis, Loughborough University, Loughborough, UK, 2011. [Google Scholar]
- Boulton, A.J.; Fenwick, G.D.; Hancock, P.J.; Harvey, M.S. Biodiversity, functional roles and ecosystem services of groundwater invertebrates. Invert. Syst. 2008, 22, 103–116. [Google Scholar] [CrossRef]
- Iepure, S.; Kalache, M.; Rasines-Ladero, R. Land-use influence on hyporheic biota from Mediterranean streams in central Spain. Studia UBB Ambientum 2020, 16, 13–42. [Google Scholar] [CrossRef]
- Zarnetske, J.P.; Haggerty, R.; Wondzell, S.M.; Baker, M.A. Labile dissolved organic carbon supply limits hyporheic denitrification. J. Geophys. Res. 2011, 116, G04036. [Google Scholar] [CrossRef]
- Brosten, T.R.; Bradford, J.H.; McNamara, J.P.; Gooseff, M.N.; Zarnetske, J.P.; Bowden, W.B.; Johnston, M.E. Multi-offset GPR methods for hyporheic zone investigations. Near Surf. Geophys. 2009, 7, 247–257. [Google Scholar] [CrossRef]
- Ward, A.S.; Schmadel, N.M.; and Wondzell, S.M. Time-Variable Transit Time Distributions in the Hyporheic Zone of a Headwater Mountain Stream. Water Resour. Res. 2018, 54, 2017–2036. [Google Scholar] [CrossRef]
- Busato, L.; Boaga, J.; Perri, M.T.; Majone, B.; Bellin, A.; Cassiani, G. Hydrogeophysical characterization and monitoring of the hyporheic and riparian zones: The Vermigliana Creek case study. Sci. Total Environ. 2019, 648, 1105–1120. [Google Scholar] [CrossRef]
- Dole-Olivier, M.J.; Marmonier, P.; Beffy, J.L. Response of invertebrates to lotic disturbance: Is the hyporheic zone a patchy refugium? Freshw. Biol. 1997, 37, 257–276. [Google Scholar] [CrossRef]
- Hancock, P.J. Human impacts on the stream–groundwater exchange zone. Environ. Manag. 2002, 29, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Dole-Olivier, M.J.; Castellarini, F.; Coineau, N.; Galassi, D.M.P.; Martin, P.; Mori, N.; Valdecasas, A.; Gibert, J. Towards an optimal sampling strategy to assess groundwater biodiversity: Comparison across six European regions. Freshw. Biol. 2009, 54, 777–796. [Google Scholar] [CrossRef]
- Iepure, S.; Martinez-Hernandez, V.; Herrera, S.; Rasines-Ladero, R.; de Bustamante, I. Response of microcrustacean communities from the surface-groundwater interface to water contamination in urban river system of the Jarama basin (central Spain). Environ. Sci. Pollut. Res. 2013, 20, 5813–5826. [Google Scholar] [CrossRef] [PubMed]
- Iepure, S.; Meffe, R.; Carreño, F.; Rasines-Ladero, R.; de Bustamante, I. Geochemical, geological and hydrological influence on ostracod assemblages distribution in the hyporheic zone of two Mediterranean rivers in central Spain. Int. Rev. Hydrobiol. 2014, 99, 435–449. [Google Scholar] [CrossRef]
- Bork, J.; Berkhoff, S.E.; Bork, S.; Hahn, H.J. Using subsurface metazoan fauna to indicate groundwater–surface water interactions in the Nakdong River floodplain, South Korea. Hydrol. J. 2009, 17, 61–75. [Google Scholar] [CrossRef]
- Stoch, F.; Artheau, M.; Brancelj, A.; Galassi, D.M.P.; Malard, F. Biodiversity indicators in European ground waters: Towards a predictive model of stygobiotic species richness. Freshwat. Biol. 2009, 54, 745–755. [Google Scholar] [CrossRef]
- Stein, H.; Kellermann, C.; Schmidt, S.I.; Brielmann, H.; Steube, C.; Berkhoff, S.E.; Fuchs, A.; Hahn, H.J.; Thulin, B.; Griebler, C. The potential use of fauna and bacteria as ecological indicators for the assessment of groundwater quality. J. Environ. Monit. 2010, 12, 242–254. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Lombardo, P.; Fiasca, B.; Di Cioccio, A.; Di Lorenzo, T.; Petitta, M.; Di Carlo, P. Earthquakes trigger the loss of groundwater biodiversity. Sci. Rep. 2014, 4, 6273. [Google Scholar] [CrossRef]
- Petitta, M.; Caschetto, M.; Galassi, D.M.P.; Aravena, R. Dualflow in karst aquifers toward a steady discharge spring (Presciano, central Italy): Influences on a subsurface groundwater dependent ecosystem and on changes related to post-earthquake hydrodynamics. Environ. Earth Sci. 2015, 73, 2609–2625. [Google Scholar] [CrossRef]
- Mori, N.; Kanduč, T.; Opalički Slabe, M.; Brancelj, A. Groundwater drift as a tracer for identifying sources of spring discharge. Groundwater 2015, 53, 123–132. [Google Scholar] [CrossRef]
- Stoch, F.; Fiasca, B.; Di Lorenzo, T.; Porfirio, S.; Petitta, M.; Galassi, D.M.P. Exploring copepod distribution patterns at three nested spatial scales in a spring system: Habitat partitioning and potential for hydrological bioindication. J. Limnol. 2016, 75, 1–13. [Google Scholar] [CrossRef]
- Boulton, A.J.; Humphreys, W.F.; Eberhard, S.M. Imperiled subsurface waters in Australia: Biodiversity, threatening processes and conservation. Aquat. Ecosys. Health 2003, 6, 41–54. [Google Scholar] [CrossRef]
- Moldovan, O.T.; Levei, E.; Banciu, M.; Banciu, H.L.; Marin, C.; Pavelescu, C.; Brad, T.; Cimpean, M.; Meleg, I.; Iepure, S.; et al. Spatial distribution patterns of the hyporheic invertebrate communities in a polluted river in Romania. Hydrobiologia 2011, 669, 63–82. [Google Scholar] [CrossRef]
- Rasines-Ladero, R.; Iepure, S. Parent lithology and organic matter influence hyporheic biota from two mediterranean rivers in central Spain. Limnetica 2016, 35, 19–36. [Google Scholar]
- AEMET. Available online: https://www.aemet.es/ (accessed on 1 June 2013).
- IGME. Available online: https://www.ign.es/web/resources/sismologia/tproximos/sismotectonica/pag_sismotectonicas/bet_centro.html (accessed on 1 February 2015).
- Arribasa, J.; Critelli, S.; Le Pera, E.C.; Tortosaa, A. Composition of modem stream sand derived from a mixture of sedimentary and metamorphic source rocks (Henares River, Central Spain). Sediment. Geol. 2000, 133, 27–48. [Google Scholar] [CrossRef]
- Benitez-Mora, A.; Camargo, J.A. Ecological Responses of Aquatic Macrophytes and Benthic Macroinvertebrates to Dams in the Henares River Basin (Central Spain). Hydrobiologia 2014, 728, 167–178. [Google Scholar] [CrossRef]
- Telford, W.M.; Geldart, L.P.; Sheriff, R.E.; Keys, D.A. Applied Geophysics, 2nd ed.; Cambridge University Press: Cambridge, UK, 1990; p. 770. [Google Scholar]
- Reynolds, J.M. An Introduction to Applied Environmental Geophysics; Wiley: New York, NY, USA, 1997; p. 780. [Google Scholar]
- Sasaki, Y. Resolution of resistivity tomography inferred from numerical simulation. Geophys. Prospect. 1992, 40, 453–464. [Google Scholar] [CrossRef]
- Loke, M.H. Electrical Imaging Surveys for Environmental and Engineering Studies. A Pract. Guide 2-D 3-D Surv. 2000, 61. [Google Scholar]
- Loke, M.H.; Wilkinson, P.B.; Chambers, P.B.; Strutt, M. Optimized arrays for 2-D cross-borehole electrical tomography surveys: Geophys. Prospect. 2014, 62, 172–189. [Google Scholar]
- Loke, M.H.; Acworth, I.; Dahlin, T. A comparison of smooth and blocky inversion methods in 2D electrical imaging surveys. Explor. Geophys. 2003, 34, 182–187. [Google Scholar] [CrossRef]
- Bernard, J. The Depth of Investigation of Electrical Methods; IRIS-instruments: Orléans, France, 2003. [Google Scholar]
- Malard, F.; Dole-Olivier, M.J.; Mathieu, J.; Stoch, F. Sampling manual for the assessment of regional groundwater biodiversity. In Sampling Manual Published within the Framework of the EU Project PASCALIS (Protocols for the ASsessment and Conservation of Aquatic Life In the Subsurface); EU: Maastricht, The Netherlands, 2002. [Google Scholar]
- Meisch, C. Freshwater Ostracoda from Western and Central Europe Süβwasserfauna von Mitteleuropa; Spektrum Akademischer Verlag: Heidelberg, Germany, 2000. [Google Scholar]
- Dussart, B.; Defaye, D. World Directory of Crustacea Copepoda of Inland Waters II—Cyclopiformes; Backhuys Publishers: Leiden, The Netherlands, 2006. [Google Scholar]
- Gibert, J.; Danielopol, D.L.; Stanford, J.A. Groundwater Ecology; Academic Press Inc.: San Diego, CA, USA, 1994. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2008, 26, 32–46. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Gower, J.C. Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Kennedy, P. A Guide to Econometrics, 4th ed.; MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Ugland, K.I.; Gray, J.S.; Ellingsen, K.E. The Species-Accumulation Curve and Estimation of Richness. J. Anim. Ecol. 2003, 72, 888–897. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rahbek, C.; Gotelli, N. The mid-domain effect and species richness patterns: What have we learned so far? Am. Nat. 2004, 163, E1–E23. [Google Scholar] [CrossRef]
- Chao, A.; Lee, S.M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Yek, S.H.; Williams, S.E.; Burwell, C.J.; Robson, S.K.A.; Crozier, R.H. Ground dwelling ants and surrogates for establishing conservation priorities in the australian wet tropics. J. Insects Sci. 2009, 9, 12. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Austral. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Exploratory null hypothesis testing for community data: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- McDonough, T.O.; Hosen, J.; Plamer, M. Temporary streams: The hydrology, geopgraphy and ecology of non-perennially flowing waters. In River Ecosystems: Dynamics, Management and Conservation, Hannah, S., Elliot, Lucas, E., Martin, Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; ISBN 978-1-61209-145-7. [Google Scholar]
- Sanchez-Hernandes, J.; Nunn, A.D. Environmental changes in a Mediterranean river: Implications for the fish assemblage. Ecohydrology 2016, 9, 1439–1451. [Google Scholar] [CrossRef]
- Bonada, N.; Rieradevall, M.; Dallas, H.; Davis, J.; Day, J.; Figueroa, R.; Resh, V.H.; Prat, N. Multi-scale assessment of macroinvertebrate richness and composition in Mediterranean-climate rivers. Freshw. Biol. 2008, 53, 772–788. [Google Scholar] [CrossRef]
- Dole-Olivier, M.-J.; Maazouzi, C.; Cellot, B.; Fiers, F.; Galassi, D.M.P.; Claret, C.; Marmonier, P. Assessing invertebrate assemblages in the subsurface zone of stream sediments (0–15 cm deep) using a hyporheic sampler. Water Resour. Res. 2014, 50, 453–465. [Google Scholar] [CrossRef]
- Nikolaidis, N.P.; Demetropoulou, L.; Froebrich, J.; Jacobs, C.; Gallart, F.; Prat, N.; Lo Porto, A.; Campana, C.; Papadoulakis, V.; Skoulikidis, N.; et al. Towards sustainable management of Mediterranean river basins: Policy recommendations on management aspects of temporary streams. Water Policy 2013, 15, 830–849. [Google Scholar] [CrossRef]
- Storey, R.; Howard, K.; Williams, D. Factors controlling rifflescale hyporheic exchange flows and their seasonal changes in a gaining stream: A three-dimensional groundwater flow model, Water Resour. Res. 2003, 39, 1034. [Google Scholar] [CrossRef]
- Rasines-Ladero, R. The Ecology of the Hyporheic Zone Associated with the Rivers Henares and Tajuña (Jarama River Basin, Spain). Ph.D. Thesis, University Rey Juan Carlos, Madrid, Spain, 2017. [Google Scholar]
- Iepure, S.; Rasines, R.; Meffe, R.; Carreño, F.; Mostaza, D.; Sundberg, A.; di Lorenzo, T.; Barroso, J. Exploring the distribution of groundwater Crustacea (Copepoda and Ostracoda) to disentangle aquifer type features—A case study in the upper Tajo basin (Central Spain). Ecohydrology 2017, 10, e1876. [Google Scholar] [CrossRef]
- Buendia, C.; Gibbins, C.N.; Vericat, D.; Batalla, R.J. Reach and catchment-scale influences on invertebrate assemblages in a river with naturally high fine sediment loads. Limnologica 2013, 43, 362–370. [Google Scholar] [CrossRef]
- Danielopol, D.L. Groundwater fauna associated with riverine aquifers. J. N. Am. Benthol. Soc. 1989, 8, 18–35. [Google Scholar] [CrossRef]
- Schwoerbel, J. Die Bedeutung des Hyporheals fur “gerollfu” hrender Hochgebirgsba¨che mit bewegter gewa¨ssers. Verh. Der Int. Ver. FuR Theor. Und Angew. Limnol. 1964, 15, 215–226. [Google Scholar]
- Ward, J.V.; Bretschko, G.; Brunke, M.; Danielopol, D.L.; Gibert, J.; Gonser, T.; Hildrew, A.G. The boundaries of river systems: The metazoan perspective. Freshw. Biol. 1998, 40, 531–569. [Google Scholar] [CrossRef]
- Robertson, A.; Rundle, S.D.; Schmid-Araya, J.M. An introduction to a special issue on lotic meiofauna. Freshw. Biol. 2000, 44, 1–3. [Google Scholar] [CrossRef]
- Marmonier, P.; Archambaud, G.; Belaidi, N.; Bougon, N.; Breil, P.; Chauvet, E.; Claret, C.; Cornut, J.; Datry, T.; Dole-Olivier, M.J.; et al. The role of organisms in hyporheic processes: Gaps in current knowledge, needs for future research and applications. Ann. Limnol.-Int. J. Lim. 2012, 48, 253–266. [Google Scholar] [CrossRef]
- Stanford, J.A.; Ward, J.V. The Hyporheic Habitat of River Ecosystems. Nature 1988, 335, 64–66. [Google Scholar] [CrossRef]
- Hakenkamp, C.C.; Palmer, M.A. The ecology of hyporheic meiofauna. In Streams and Ground Waters; Jones, J.B., Mulholland, P.J., Eds.; Academic Press: London, UK, 2000. [Google Scholar]
- Stanley, E.H.; Boulton, A.J. Hydrology and the distribution of hyporheos: Perspectives from a mesic river and a desert stream. J. N. Am. Benthol. Soc. 1993, 12, 79–83. [Google Scholar] [CrossRef]
- Trayler, K.M.; Davis, J.A. Forestry impacts and the vertical distribution of stream invertebrates in southwestern Australia. Freshw. Biol. 1998, 40, 331–342. [Google Scholar] [CrossRef]
- Dumas, P.; Bou, C.; Gibert, J. Groundwater macrocrustaceans as natural indicators of the Ariége alluvial aquifer. Int. Rev. Hydrobiol. 2001, 86, 619–633. [Google Scholar] [CrossRef]
- Cannan, C.E.; Armitage, P.D. The influence of catchment geology on the longitudinal distribution of macroinvertebrate assemblages in a groundwater dominated river. Hydrol. Process. 1999, 13, 355–369. [Google Scholar] [CrossRef]
- Malard, F.; Tockner, K.; Dole-Olivier, M.-J.; Ward, J.V. A landscape perspective of surface–subsurface hydrological exchanges in river 30 corridors. Freshw. Biol. 2002, 47, 621–640. [Google Scholar] [CrossRef]
- Malard, J.; Adamowski, J.; Días, M.R.; Yax, H.T.; Arévalo-Rodríguez, L.A.; Melgar-Quiñonez, H. Agroecological food webs: Modelling a missing piece to climate change adaptation. Geophys. Res. Abstr. 2019, 21, EGU2019-11656. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Units/Indices |
|---|---|
| Geographical | Altitude (m a.s.l.) |
| Lithology | |
| Hydrology | Water depth (m) |
| Channel width (m) | |
| Surface water discharge (m/s) | |
| Surface water discharge (m/s monthly average) | |
| Chemico-physical | Conductivity (μS/cm, EC) |
| Non-purgeable organic carbon (in mg/L−1, NPOC) | |
| Total carbon (TC) | |
| Inorganic carbon (IC) | |
| Total organic carbon (mg/L−1, TOC) | |
| Dissolved organic carbon (mg/L−1, DOC) | |
| Total suspended solids (mg/L−1, TSS) | |
| Total alkalinity (mg/L−1, as HCO3−) | |
| Cations Mg2+, K+, Ca2+ | |
| Major anions Cl−, SO42−, NO3− | |
| Biological | Total abundance of species |
| Species richness (SR) | |
| Stygobites number (SB) | |
| Non-stygobites number (nSB) | |
| Statistical levels | Sub-basins (SUB): Henares (HEN), Tajuna (TAJ) |
| Habitats (HAB): surface (SW), hyporheic zone (HYPORHEIC ZONE) Season (SEA): cold (C), mild (M) |
| Sites/ Environmental Variables | Site 1 Hera de Ayuso (HEN) | Site 2 Romanones (TAJ) | |||
|---|---|---|---|---|---|
| SW/HW | June 2013 | April 2014 | June 2013 | April 2014 | |
| Depth (cm) | SW | ||||
| HW | |||||
| Water discharge * (m/s) | SW | 2.64 | 2.5 | 0.46 | 0.48 |
| Water discharge (m/s monthly average) | SW | 2.83 | 3.6 | 0.52 | 0.49 |
| Temperature (°C) | SW | 15.6± | 13.23± | 11.86± | 6.43± |
| HW | 16.75± | 13.75± | 12.45± | 7.1± | |
| Dissolved O2 (mg/L) | SW | 9± | 10.6± | 12.46± | |
| HW | 8.91± | 8.45± | 10.14± | ||
| pH | SW | 8.04± | 8.4± | 8.36± | 8.1± |
| HW | 8.17± | 8.24± | 8.15± | 7.7± | |
| Conductivity (μS/cm) | SW | 1107± | 1024± | 549.3± | 736.6± |
| HW | 1102.5± | 1017± | 555± | 779.5± | |
| ER (Ohm) | HW | 19.1–73.7 | 47.9–77.5 | ||
| Biotic parameters | |||||
| Total abundance * | HZ | 595 | 404 | 195 | 1226 |
| Species richness ** | HZ | 9 | 6 | 3 | 8 |
| Stygobites ** | HZ | 3 | 2 | 0 | 1 |
| Non-stygobites | HZ | 6 | 4 | 3 | 7 |
| Species | Contrib% | Cum.% |
|---|---|---|
| Paracyclops imminutus | 16.35 | 16.35 |
| Bryocampthus sp. 2 | 12.47 | 28.81 |
| Microcyclops rubellus | 11.62 | 40.43 |
| Macrocyclops albidus | 11.50 | 51.94 |
| Diacyclops languidoides lang. ssp | 11.22 | 63.16 |
| Microcyclops sp. | 7.41 | 70.57 |
| Parastenocaris sp.1 | 6.63 | 77.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iepure, S.; Gomez-Ortiz, D.; Lillo, J.; Rasines-Ladero, R.; Lorenzo, T.D. Applying Electrical Resistivity Tomography and Biological Methods to Assess the Hyporheic Zone Water Exchanges in Two Mediterranean Stream Reaches. Water 2022, 14, 3396. https://doi.org/10.3390/w14213396
Iepure S, Gomez-Ortiz D, Lillo J, Rasines-Ladero R, Lorenzo TD. Applying Electrical Resistivity Tomography and Biological Methods to Assess the Hyporheic Zone Water Exchanges in Two Mediterranean Stream Reaches. Water. 2022; 14(21):3396. https://doi.org/10.3390/w14213396
Chicago/Turabian StyleIepure, Sanda, David Gomez-Ortiz, Javier Lillo, Rubén Rasines-Ladero, and Tiziana Di Lorenzo. 2022. "Applying Electrical Resistivity Tomography and Biological Methods to Assess the Hyporheic Zone Water Exchanges in Two Mediterranean Stream Reaches" Water 14, no. 21: 3396. https://doi.org/10.3390/w14213396
APA StyleIepure, S., Gomez-Ortiz, D., Lillo, J., Rasines-Ladero, R., & Lorenzo, T. D. (2022). Applying Electrical Resistivity Tomography and Biological Methods to Assess the Hyporheic Zone Water Exchanges in Two Mediterranean Stream Reaches. Water, 14(21), 3396. https://doi.org/10.3390/w14213396