
Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean

Abstract
1. Introduction
2. Materials and Methods
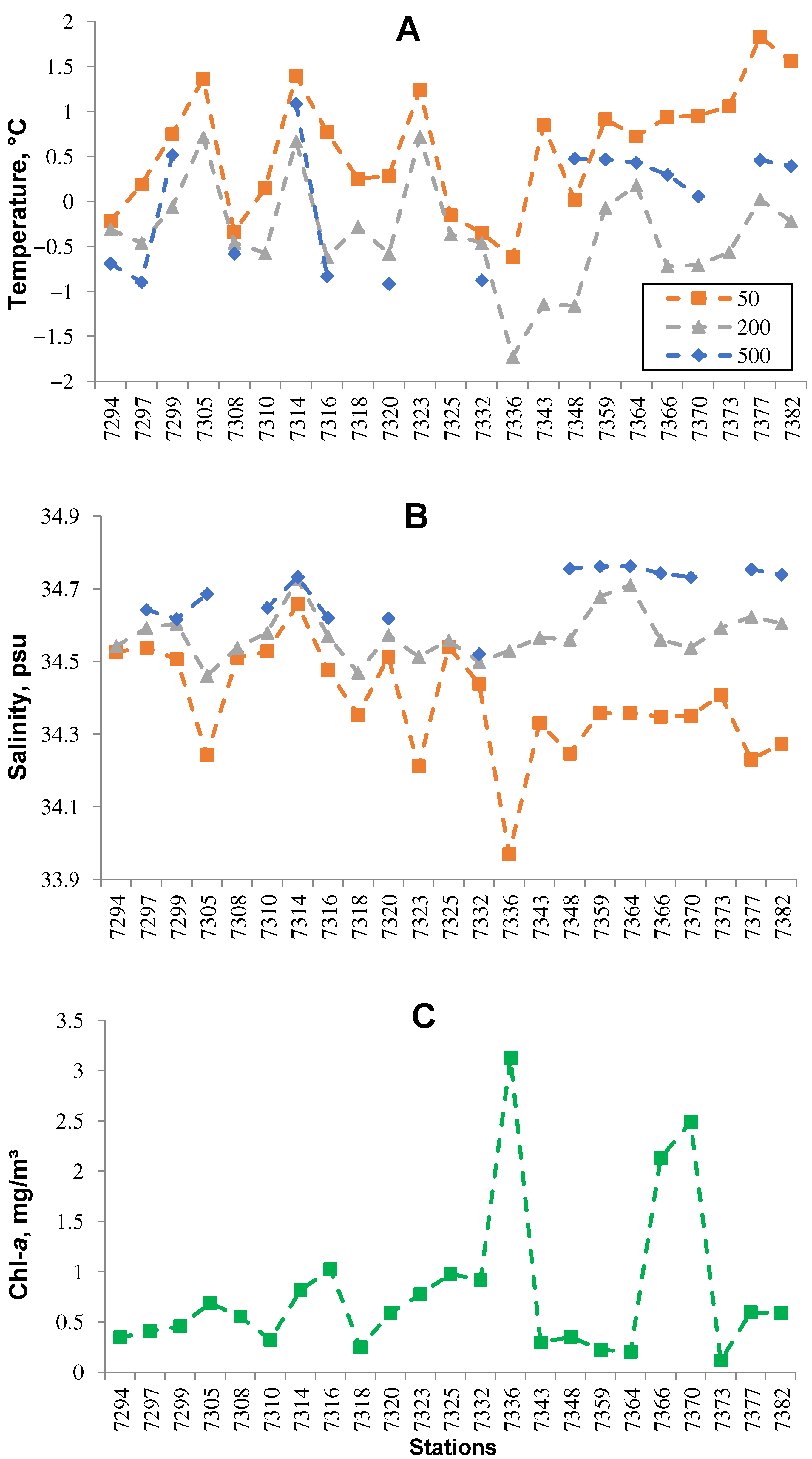
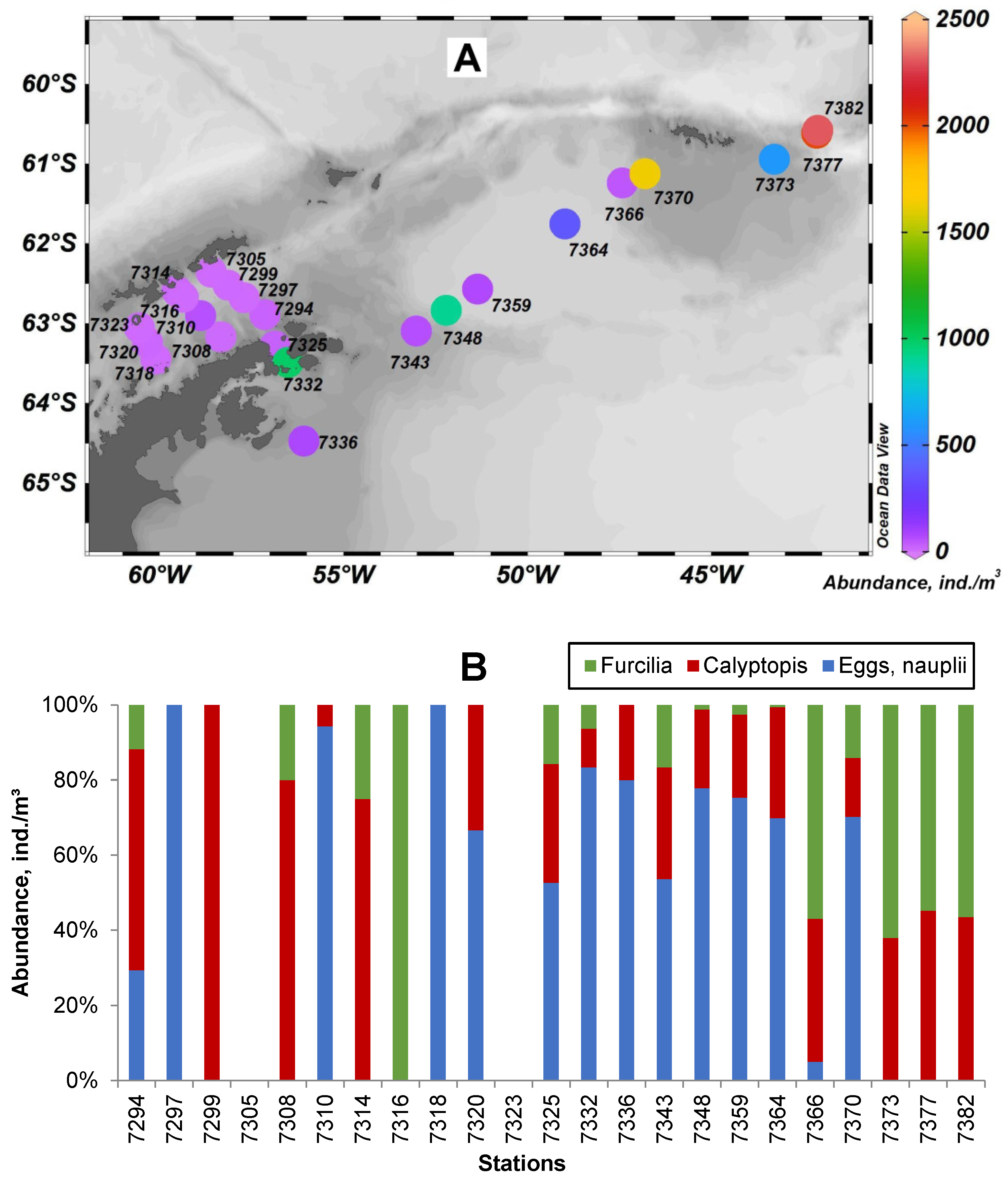
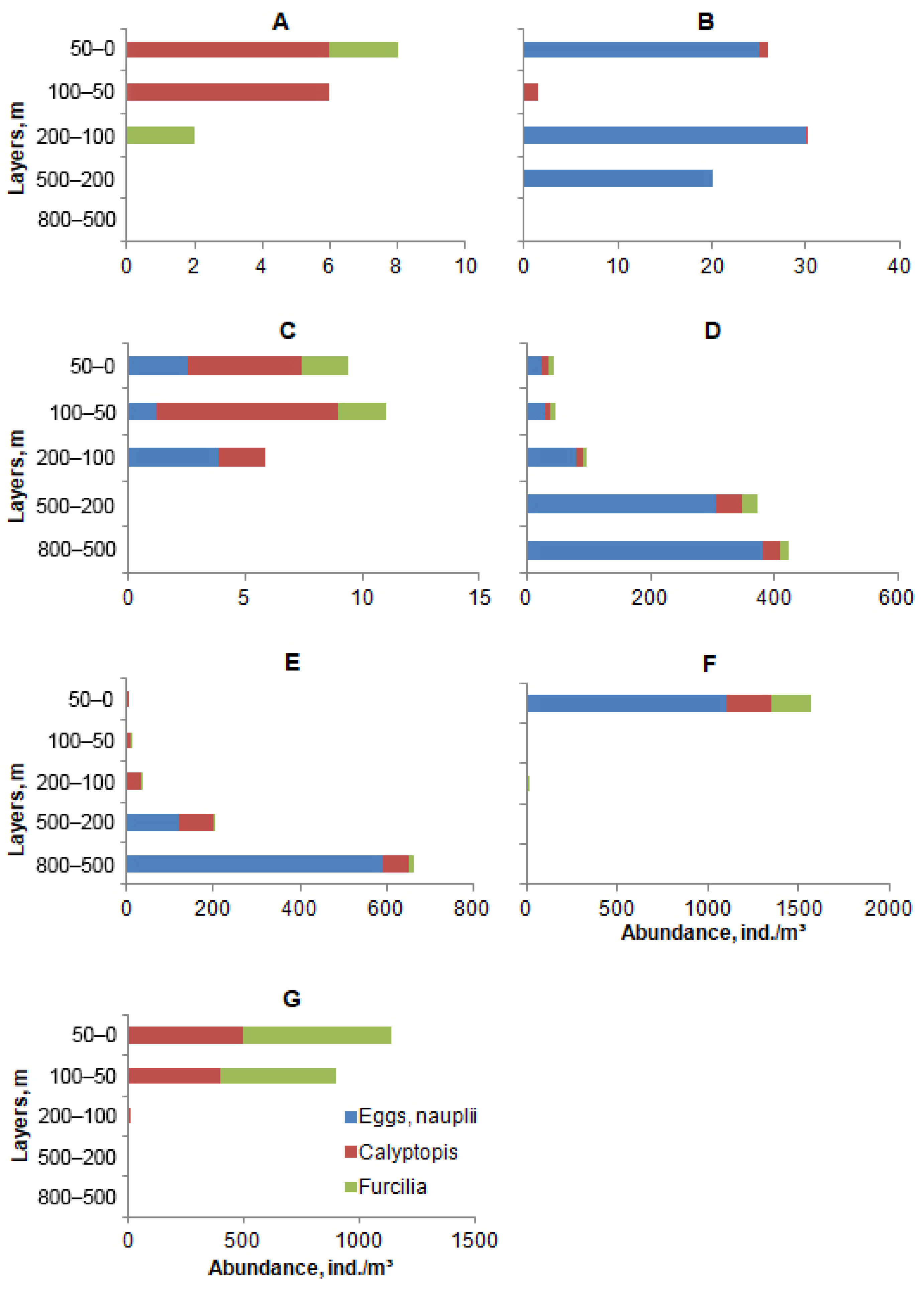
3. Results
4. Discussion and Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laws, R. Seals and whales of the Southern Ocean. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 279, 81–96. [Google Scholar] [CrossRef]
- Kawaguchi, S.; Nicol, S. Learning about Antarctic krill from the fishery. Antar. Sci. 2007, 19, 219–230. [Google Scholar] [CrossRef]
- Santa Cruz, F.; Ernst, B.; Arata, J.A.; Parada, C. Spatial and temporal dynamics of the Antarctic krill fishery in fishing hotspots in the Bransfield Strait and South Shetland Islands. Fish. Res. 2018, 208, 157–166. [Google Scholar] [CrossRef]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Hill, S.L.; Pakhomov, E.A.; Siegel, V.; Reiss, C.S.; Loeb, V.J.; Steinberg, D.K.; Schmidt, K.; Tarling, G.A.; Gerrish, L.; et al. Krill (Euphausia superba) distribution contracts southward during rapid regional warming. Nat. Clim. Change 2019, 9, 142–147. [Google Scholar] [CrossRef]
- Yang, G.; Atkinson, A.; Hill, S.L.; Guglielmo, L.; Granata, A.; Li, C. Changing circumpolar distributions and isoscapes of Antarctic krill: Indo-Pacific habitat refuges counter long-term degradation of the Atlantic sector. Limnol. Oceanogr. 2020, 66, 272–287. [Google Scholar] [CrossRef]
- Voronina, N.M. Comparative abundance and distribution of major filter-feeders in the Antarctic pelagic zone. J. Mar. Sys. 1998, 17, 375–390. [Google Scholar] [CrossRef]
- Hewes, C.D.; Reiss, C.S.; Holm-Hansen, O. A quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region. Deep-Sea Res. II 2009, 56, 1230–1241. [Google Scholar] [CrossRef]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schloss, I.R.; Hoffmeyer, M.S. Micro- and mesozooplankton successions in an Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, e0232614. [Google Scholar] [CrossRef] [PubMed]
- Pakhomov, E.; Pshenichnov, L.; Krot, A.; Paramonov, V.; Slypko, I.; Zabroda, P. Zooplankton distribution and community structure in the Pacific and Atlantic Sectors of the Southern Ocean during austral summer 2017–2018: A Pilot Study Conducted from Ukrainian Long-Liners. J. Mar. Sci. Eng. 2020, 8, 488. [Google Scholar] [CrossRef]
- Makarov, R.R. Larvae of Euphausia superba Dana in plankton from the Sea of Scotia. Tr. Vses. Nauchno-Issled. Ryb. Okeanogr. 1974, 99, 84–103. [Google Scholar]
- Shulenberger, E.; Wormuth, J.H.; Loeb, V.J. A large swarm of Euphausia superba: Overview of path structure and composition. J. Crustacean Biol. 1984, 4, 75–95. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, Z.; Huang, H.; Chen, X.; Feng, C.; Li, L. Geographical distribution and age composition of Euphausia superba larvae (Crustacea: Euphausiacea) in the South Shetland Islands region and southern Scotia Sea in relation to environmental conditions. Acta Oceanol. Sin. 2013, 32, 59–67. [Google Scholar] [CrossRef]
- Sologub, D.O. Distribution and age structure of euphausiids larvae at the Western Antarctic Peninsula (Subarea 48.1). Trudy VNIRO 2015, 154, 3–15. [Google Scholar]
- Yakovenko, V.A.; Spiridonov, V.A.; Gorbatenko, K.M.; Shadrin, N.V.; Samyshev, E.Z.; Minkina, N.I. Macro- and mesozooplankton in the Powell Basin (Antarctica): Species composition and distribution of abundance and biomass in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 131–141. [Google Scholar] [CrossRef]
- Meredith, M.P.; King, J.C. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Let. 2005, 32, L19604. [Google Scholar] [CrossRef]
- Stammerjohn, S.E.; Martinson, D.G.; Smith, R.C.; Yuan, X.; Rind, D. Trends in Antarctic annual sea ice retreat and advance and their relation to El Nino-Southern Oscillation and Southern Annular Mode variability. J. Geophys. Res Ocean. 2008, 113, C03S90. [Google Scholar] [CrossRef]
- Meredith, M.; Sommerkorn, S.; Cassotta, C.; Derksen, A.; Ekaykin, A.; Hollowed, G.; Kofinas, A.; Mackintosh, J.; Melbourne-Thomas, M.M.C.; Muelbert, G.; et al. Polar Regions. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; pp. 203–320. [Google Scholar] [CrossRef]
- Anisimov, O.A.; Vaughan, D.G.; Callaghan, T.V.; Furgal, C.; Marchant, H.; Prowse, T.D.; Vilhjalmsson, H.; Walsh, J.E. Polar regions (Arctic and Antarctic). In Climate Change 2007: Impacts, Adaptation and Vulnerability; Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, P.J., van der Linden, P., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 653–685. [Google Scholar]
- Xiao, C. Changes in antarctic climate system: Past, present and future. Adv. Clim. Change Res. 2008, 4, 1–7. [Google Scholar]
- Behrenfeld, M.J. Climate-mediated dance of the plankton. Nat. Clim. Change 2014, 4, 880–887. [Google Scholar] [CrossRef]
- Johnston, N.M.; Murphy, E.J.; Atkinson, A.; Constable, A.J.; Cotte, C.; Cox, M.; Daly, K.L.; Driscoll, R.; Flores, H.; Halfter, S.; et al. Status, change, and futures of zooplankton in the Southern Ocean. Front. Ecol. Evol. 2022, 9, 624692. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Kirsch, E. Effect of temperature on developmental times and survival of early larval stages of Euphausia superba Dana. J. Exp. Mar. Bio. Ecol. 1988, 121, 55–71. [Google Scholar] [CrossRef]
- Perry, F.A.; Kawaguchi, S.; Atkinson, A.; Sailley, S.F.; Tarling, G.A.; Mayor, D.J.; Lucas, C.H.; King, R.; Cooper, A. Temperature–induced hatch failure and nauplii malformation in Antarctic krill. Front. Mar. Sci. 2020, 7, 501. [Google Scholar] [CrossRef]
- Weikert, H.; John, H.-C. Experiences with a modified be multiple opening–closing plankton net. J. Plankton Res. 1981, 3, 167–176. [Google Scholar] [CrossRef]
- Tranter, D.J. Reviews on zooplankton sampling methods. In Zooplankton Sampling; Part I; Tranter, D.J., Ed.; UNESCO: Paris, France, 1968; pp. 11–144. [Google Scholar]
- Smith, P.E.; Richardson, S. Standard techniques for pelagic fish egg and larva surveys. FAO Fish. Tech. Pap. 1977, 175, 27–73. [Google Scholar]
- Pertsova, K.N. Larvae of euphausiids of the Antarctic. In Ecology and biogeography of plankton: Proceedings of the Institute of Oceanology RAS; Nauka: Moscow, Russia, 1976; Volume 105, pp. 147–170. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER–E Ltd.: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Heywood, K.J.; Garabato, A.C.N.; Stevens, D.P.; Muench, R.D. On the fate of the Antarctic Slope Front and the origin of the Weddell Front. J. Geophys. Res. 2004, 109, C06021. [Google Scholar] [CrossRef]
- Thompson, A.F.; Heywood, K.J.; Thorpe, S.E.; Renner, A.H.H.; Trasvica, A. Surface circulation at the tip of the Antarctic Peninsula from drifters. J. Phys. Oceanog. 2009, 39, 3–26. [Google Scholar] [CrossRef]
- Morozov, E.G.; Flint, M.V.; Spiridonov, V.A. Antarctic Peninsula region of the Southern Ocean; Advances in Polar Ecology; Springer: Cham, Switzerland, 2021; Volume 6, p. 433. [Google Scholar] [CrossRef]
- Zhou, X.Q.; Zhu, G.P.; Hu, S. Influence of tides on mass transport in the Bransfield Strait and the adjacent areas. Antarctic Polar Sci. 2020, 23, 100506. [Google Scholar] [CrossRef]
- Fedotova, A.A.; Kashin, S.V. Interannual variations of water mass properties in the central basin of the Bransfield Strait. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 131–141. [Google Scholar] [CrossRef]
- Krechik, V.A.; Frey, D.I.; Morozov, E.G. Peculiarities of water circulation in the central part of the Bransfield Strait in January 2020. Dokl. Earth Sci. 2021, 496, 92–95. [Google Scholar] [CrossRef]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and southern part of the Drake Passage using a method of statistical multidimensional analysis. Polish Pol. Res. 1987, 8, 333–366. [Google Scholar]
- Krek, A.V.; Krek, E.V.; Krechik, V.A. The Circulation and Mixing Zone in the Antarctic Sound in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 83–100. [Google Scholar] [CrossRef]
- Fedotova, A.A.; Stepanova, S.V. Water mass transformation in the Powell Basin. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 115–129. [Google Scholar] [CrossRef]
- Deacon, G.E.R. A general account of the hydrology of the South Atlantic Ocean. Discov. Rep. 1933, 7, 171–238. [Google Scholar]
- Kittel, W.; Jazdzewski, K. Studies on the larval stages of Euphausia superba Dana (Crustacea, Euphausiacea) in the southern Drake Passage and in the Bransfield Strait in February and March 1981 during the BIOMASS-FIBEX expedition. Pol. Polar Res. 1982, 3, 273–280. [Google Scholar]
- Spiridonov, V.A.; Zalota, A.K.; Yakovenko, V.A.; Gorbatenko, K.M. Composition of population and transport of juveniles of Antarctic krill in Powell Basin region (northwestern Weddell Sea) in January 2020. Trudy VNIRO 2020, 181, 33–51. [Google Scholar] [CrossRef]
- Voronina, N.M. Pelagic Ecosystems of the Southern Ocean; Nauka: Moscow, Russia, 1984; p. 206. [Google Scholar]
- Orsi, A.H.; Whitworth, T.; Nowlin, W.D., Jr. On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep Sea Res. I 1995, 42, 641–673. [Google Scholar] [CrossRef]
- Morozov, E.G.; Krechik, V.A.; Frey, D.I.; Polukhin, A.A.; Artemiev, V.A.; Kasyan, V.V.; Sapozhnikov, P.V.; Mukhametianov, R.Z. Frontal Zone Between Relatively Warm and Cold Waters in the Northern Weddell Sea. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 31–54. [Google Scholar] [CrossRef]
- Melnikov, I.; Spiridonov, V. Antarctic krill under perennial sea ice in the western Weddell Sea. Antarct. Sci. 1996, 8, 323–429. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Baker, K.S.; Vernet, M.; Smith, R.C. Growth limitation in young Euphausia superba under field conditions. Limnol. Oceanogr. 2000, 45, 31–43. [Google Scholar] [CrossRef]
- Hempel, I.; Hempel, G. Field observations on the developmental ascent of larval Euphausia superba (Crustacea). Polar Biol. 1986, 6, 123–126. [Google Scholar] [CrossRef]
- Spiridonov, V.A. Spatial and temporal variability in reproductive timing of Antarctic krill (Euphausia superba). Polar Biol. 1995, 15, 161–174. [Google Scholar] [CrossRef]
- Makarov, R.R.; Menshenina, L.L. On the study of distribution and phenology of euphausiid larvae in the Scotia Sea. Antarctica 1988, 28, 166–174. [Google Scholar]
- Hofmann, E.E.; Klinck, J.M.; Locarnini, R.A.; Fach, B. Krill transport in the Scotia Sea and environs. Antarct. Sci. 1998, 10, 406–415. [Google Scholar] [CrossRef]
- Siegel, V.; Watkins, J.L. Distribution, Biomass and Demography of Antarctic Krill, Euphausia superba. In Biology and Ecology of Antarctic Krill; Advances in Polar Ecology; Siegel, V., Ed.; Springer: Cham, Switzerland, 2016; pp. 21–100. [Google Scholar] [CrossRef]
- Meyer, B.; Atkinson, A.; Blume, B.; Bathmann, U.V. Feeding and energy budgets of larval Antarctic krill Euphausia superba in summer. Mar. Ecol. Prog. Ser. 2003, 57, 167–177. [Google Scholar] [CrossRef]
- Schmidt, K.; Atkinson, A.; Venables, H.J.; Pond, D.W. Early spawning of Antarctic krill in the Scotia Sea is fuelled by “superfluous” feeding on non–ice associated phytoplankton blooms. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2012, 59–60, 159–172. [Google Scholar] [CrossRef]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of Southern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- McBride, M.M.; Dalpadado, P.; Drinkwater, K.F.; Godo, O.R.; Hobday, A.J.; Hollowed, A.B.; Kristiansen, T.; Murphy, E.J.; Ressler, P.H.; Subbey, S.; et al. Krill, climate, and contrasting future scenarios for Arctic and Antarctic fisheries. ICES J. Mar. Sci. 2014, 71, 1934–1955. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Constable, A.J.; Melbourne-Thomas, J.; Corney, S.P.; Arrigo, K.R.; Barbraud, C.; Barnes, D.K.A.; Bindoff, N.L.; Boyd, P.W.; Brandt, A.; Costa, D.P.; et al. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Glob. Change Biol. 2014, 20, 3004–3025. [Google Scholar] [CrossRef]
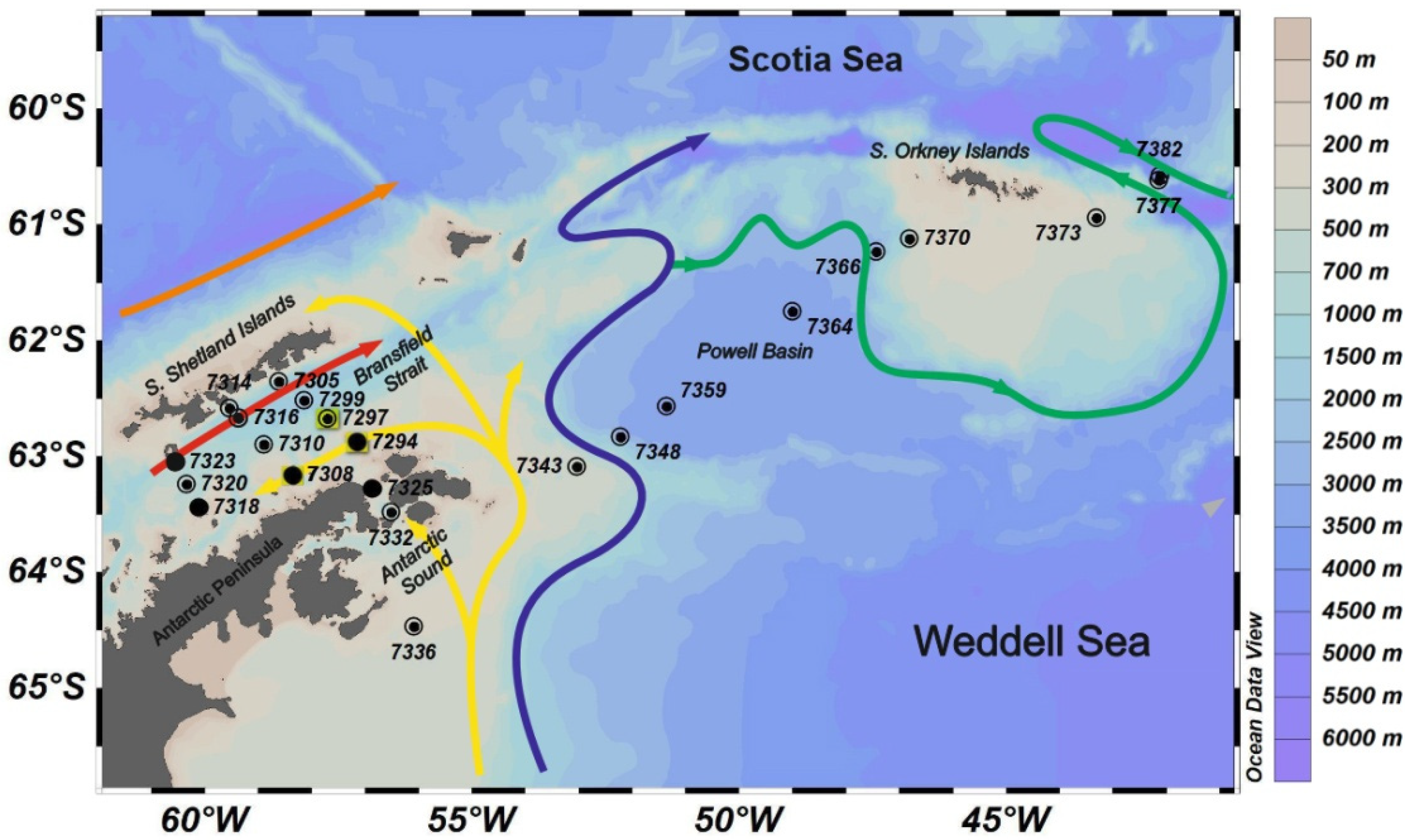
 —Multinet,
—Multinet,  —Bongo, and ●—WP-2.
—Multinet, —Bongo, and ●—WP-2.
—Bongo, and ●—WP-2.
—Multinet, —Bongo, and ●—WP-2.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | ρ | Mean ± SD | |||
|---|---|---|---|---|---|
| Group A | Group B | Group C | Group D | ||
| SST, °C | 0.307 | 0.36 ± 0.65 | 1.48 ± 0.39 | 0.34 ± 0.61 | 0.22 ± 0.04 |
| 200 m T, °C | 0.096 | –0.48 ± 0.61 | –0.25 ± 0.30 | –0.54 ± 0.56 | –0.37 ± 0.13 |
| 500 m T, °C | 0.089 | 0.03 ± 0.75 | 0.43 ± 0.05 | 0.02 ± 0.63 | –0.90 ± 0.07 |
| SSS, psu | 0.101 | 34.43 ± 0.18 | 34.30 ± 0.09 | 34.35 ± 0.08 | 34.44 ± 0.13 |
| 200 m S, psu | 0.093 | 34.59 ± 0.06 | 34.61 ± 0.02 | 34.58 ± 0.09 | 34.53 ± 0.09 |
| 500 m S, psu | 0.081 | 34.69 ± 0.07 | 34.75 ± 0.01 | 34.69 ± 0.12 | 34.64 ± 0.03 |
| Chl a, mg/m3 | 0.163 | 0.89 ± 0.91 | 0.43 ± 0.27 | 0.99 ± 1.05 | 0.33 ± 0.11 |
| BEST combination of factors (SST and Chl a) | 0.369 | ||||
| SST | SSS | 200 m T | 200 m S | 500 m T | 500 m S | Chl a | E and N | C | F | |
|---|---|---|---|---|---|---|---|---|---|---|
| E and N | –0.38 | –0.16 | –0.44 | –0.02 | –0.08 | 0.29 | 0.36 | 1 | ||
| C | 0.22 | –0.32 | –0.20 | 0.37 | 0.41 | 0.57 | –0.09 | 0.58 | 1 | |
| F | 0.33 | –0.18 | –0.16 | 0.26 | 0.23 | 0.44 | –0.06 | 0.32 | 0.86 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasyan, V.V. Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water 2022, 14, 3196. https://doi.org/10.3390/w14203196
Kasyan VV. Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water. 2022; 14(20):3196. https://doi.org/10.3390/w14203196
Chicago/Turabian StyleKasyan, Valentina V. 2022. "Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean" Water 14, no. 20: 3196. https://doi.org/10.3390/w14203196
APA StyleKasyan, V. V. (2022). Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water, 14(20), 3196. https://doi.org/10.3390/w14203196