
Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
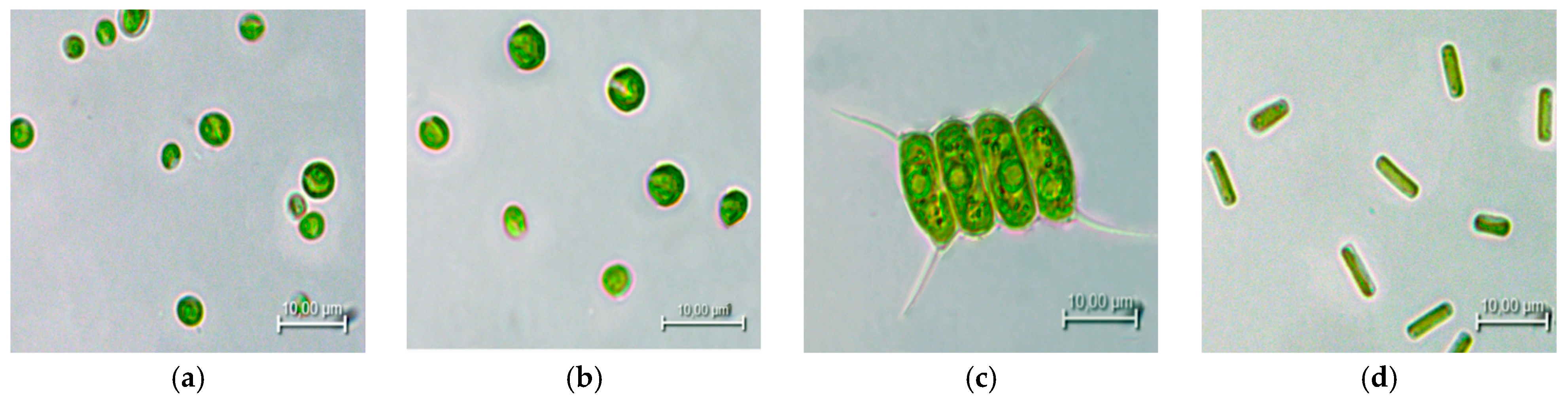
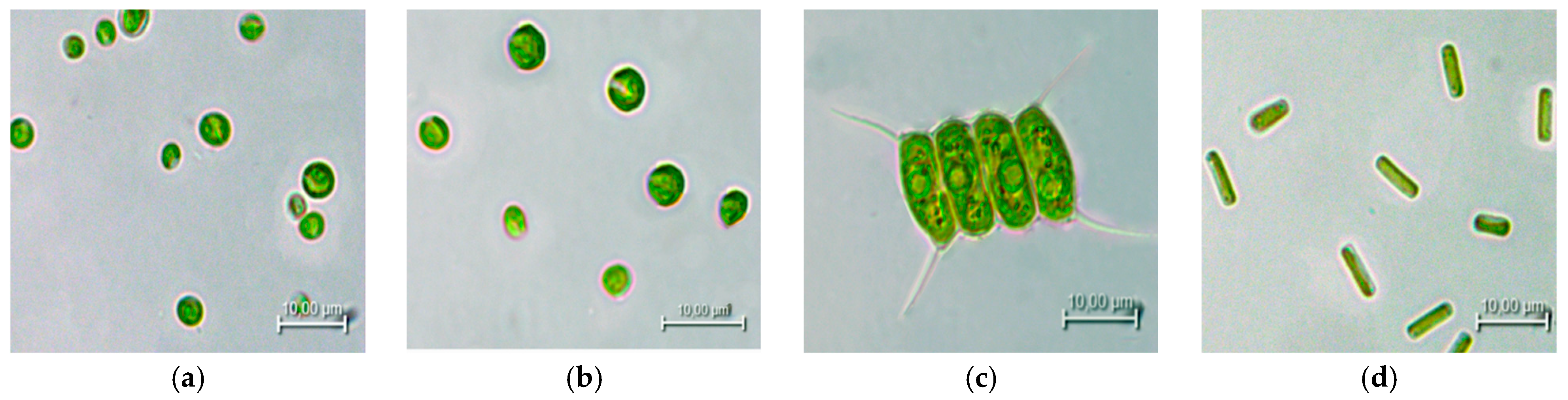
2.1. Microalgae Species
2.2. ADE Collection
2.3. Pre-Cultures Maintenance
2.4. Species Cultivation
2.4.1. Media Composition
2.4.2. Culture Conditions
2.4.3. pH Control
2.5. Analytical Measurements
2.5.1. Growth Determination
2.5.2. Nutrient Analysis
2.5.3. Pigments Determination
2.5.4. Lipids Determination
2.5.5. Proteins Determination
2.5.6. Carbohydrates Determination
2.6. Statistical Analysis
3. Results and Discussion
3.1. ADE Composition
3.2. Screening of Microalgae Species
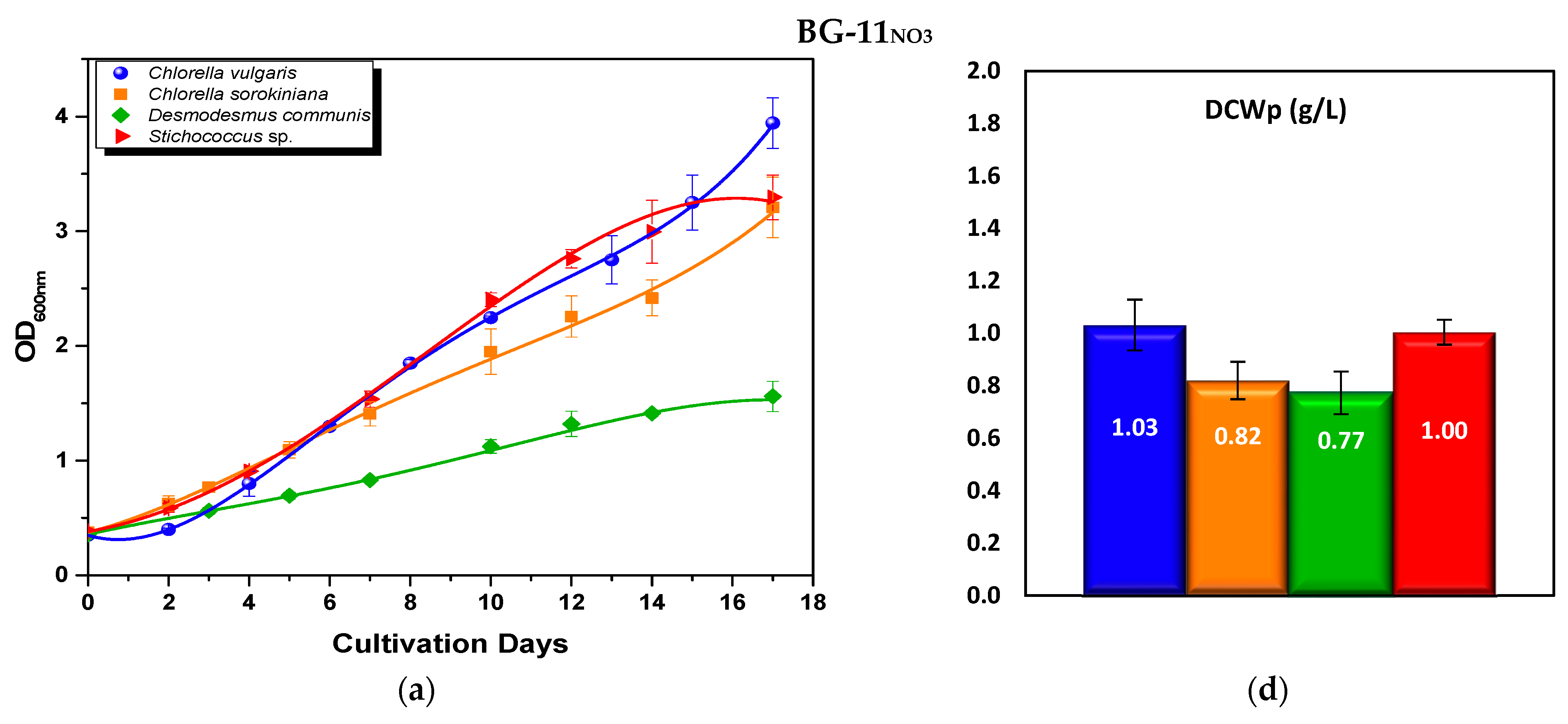
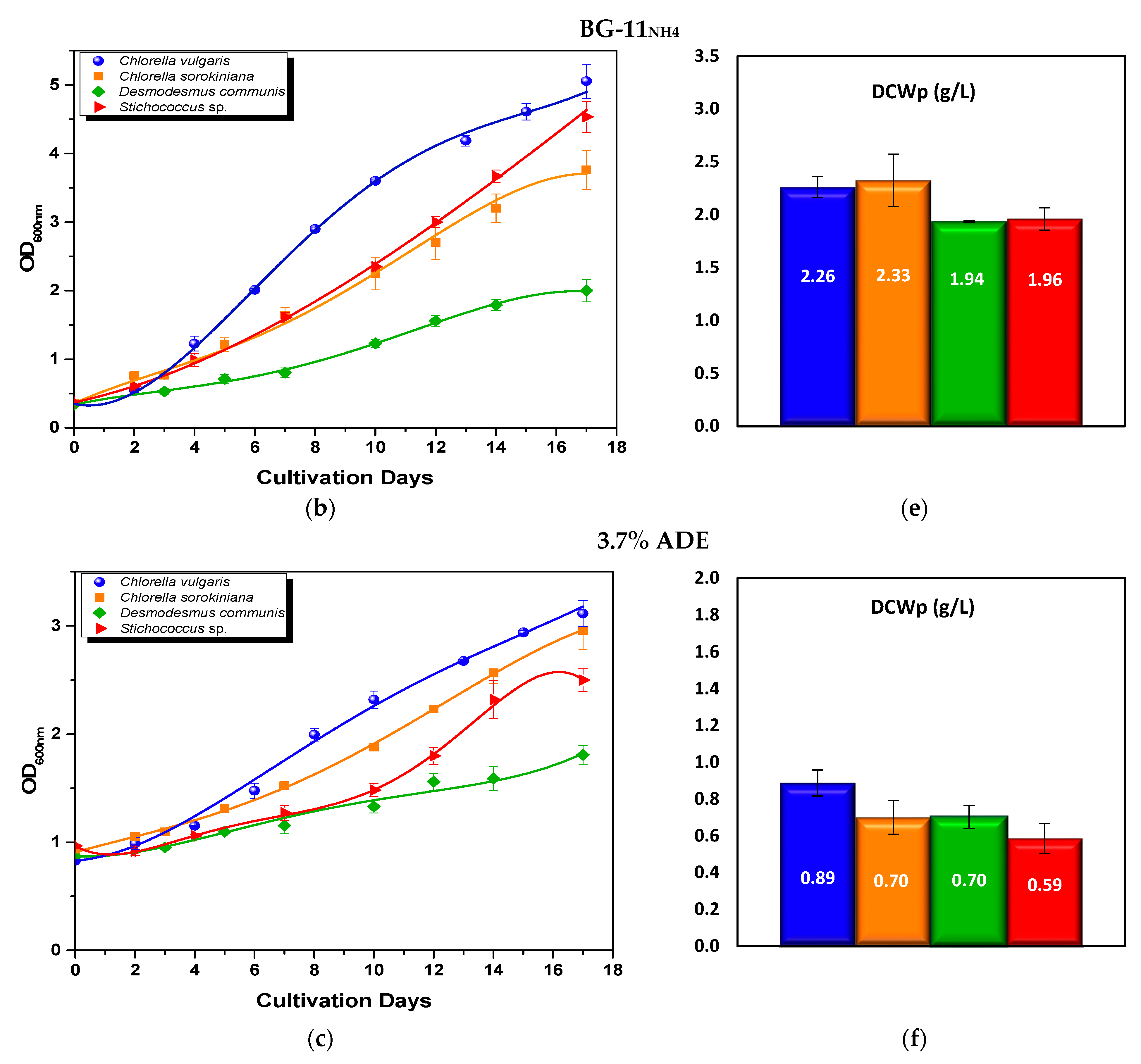
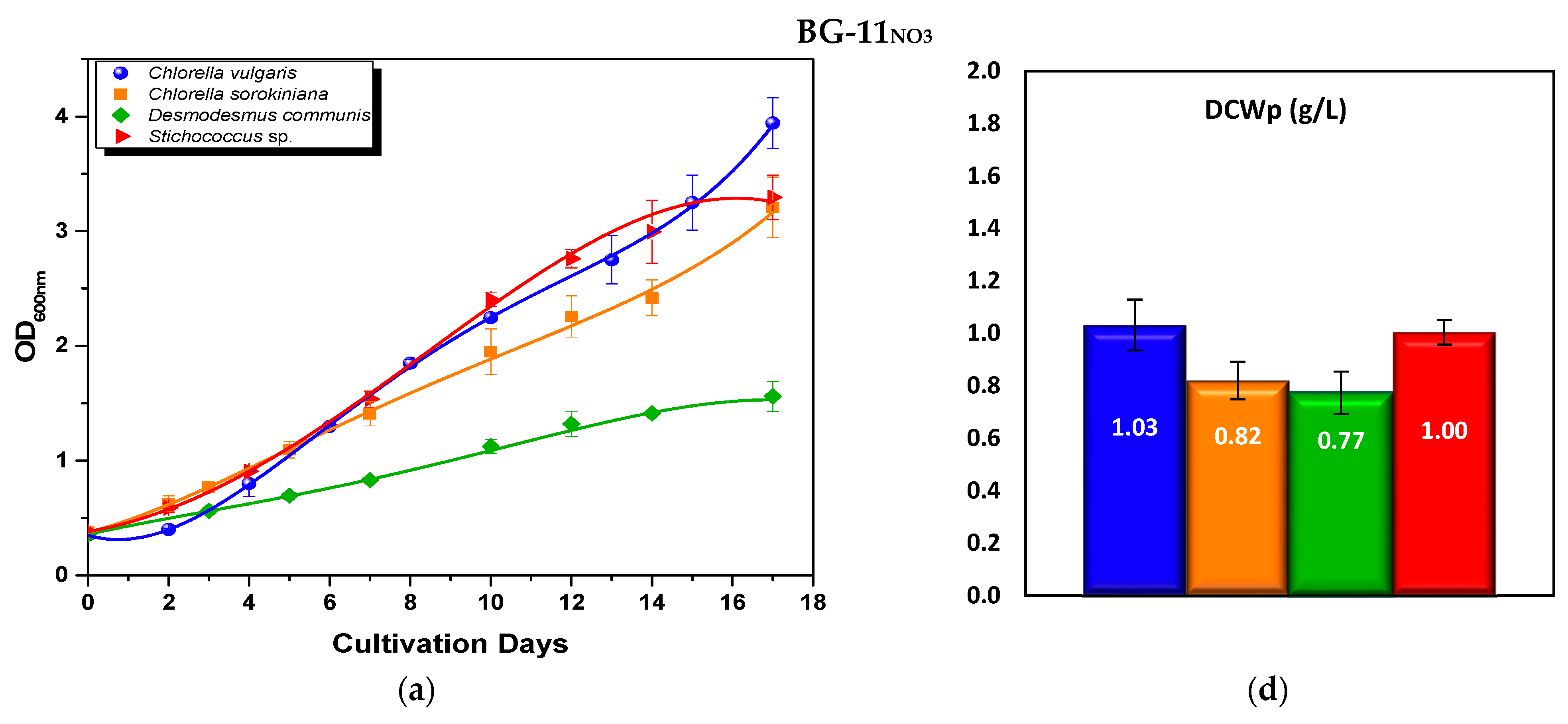
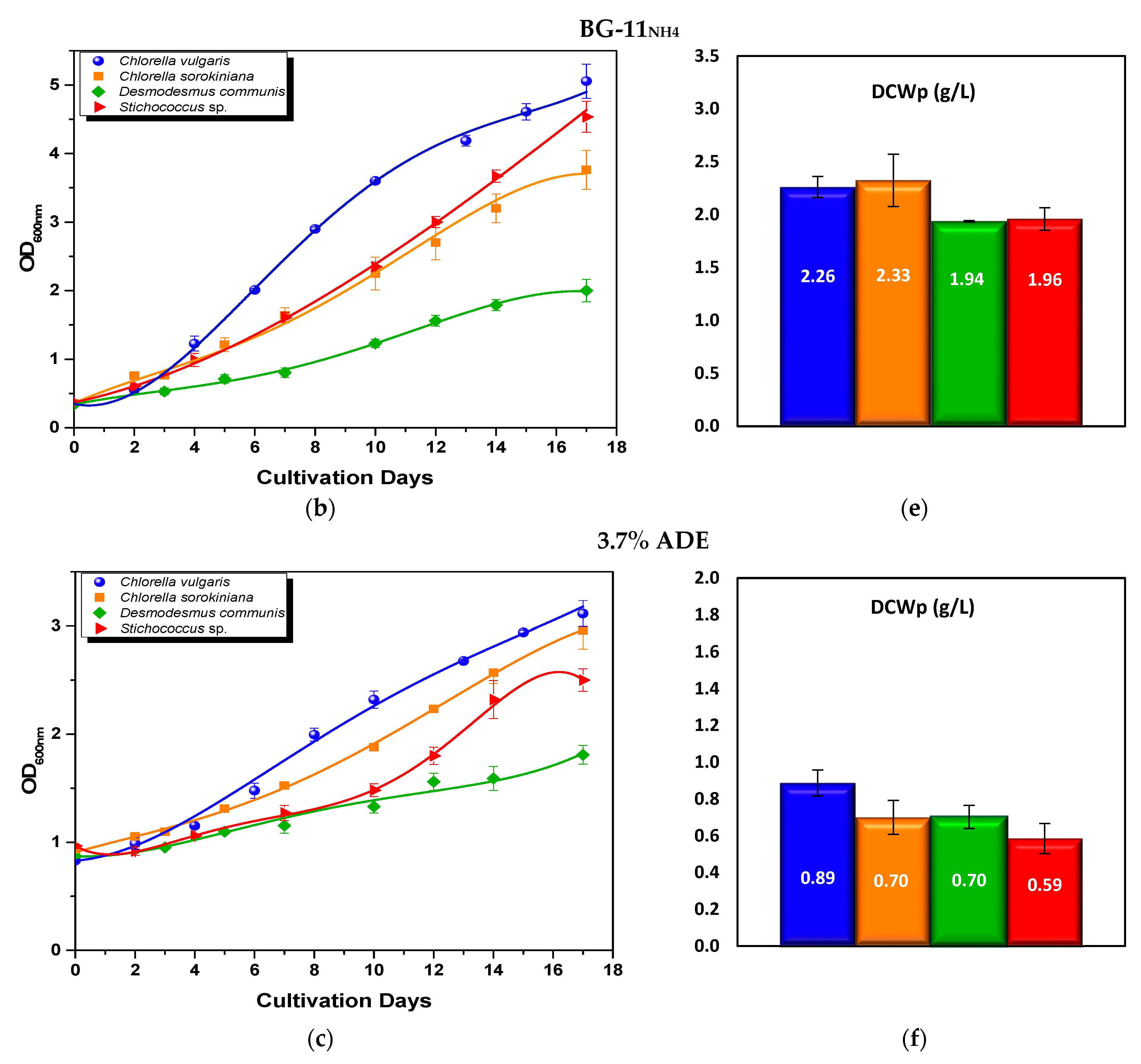
3.2.1. Optical Density and Biomass Production
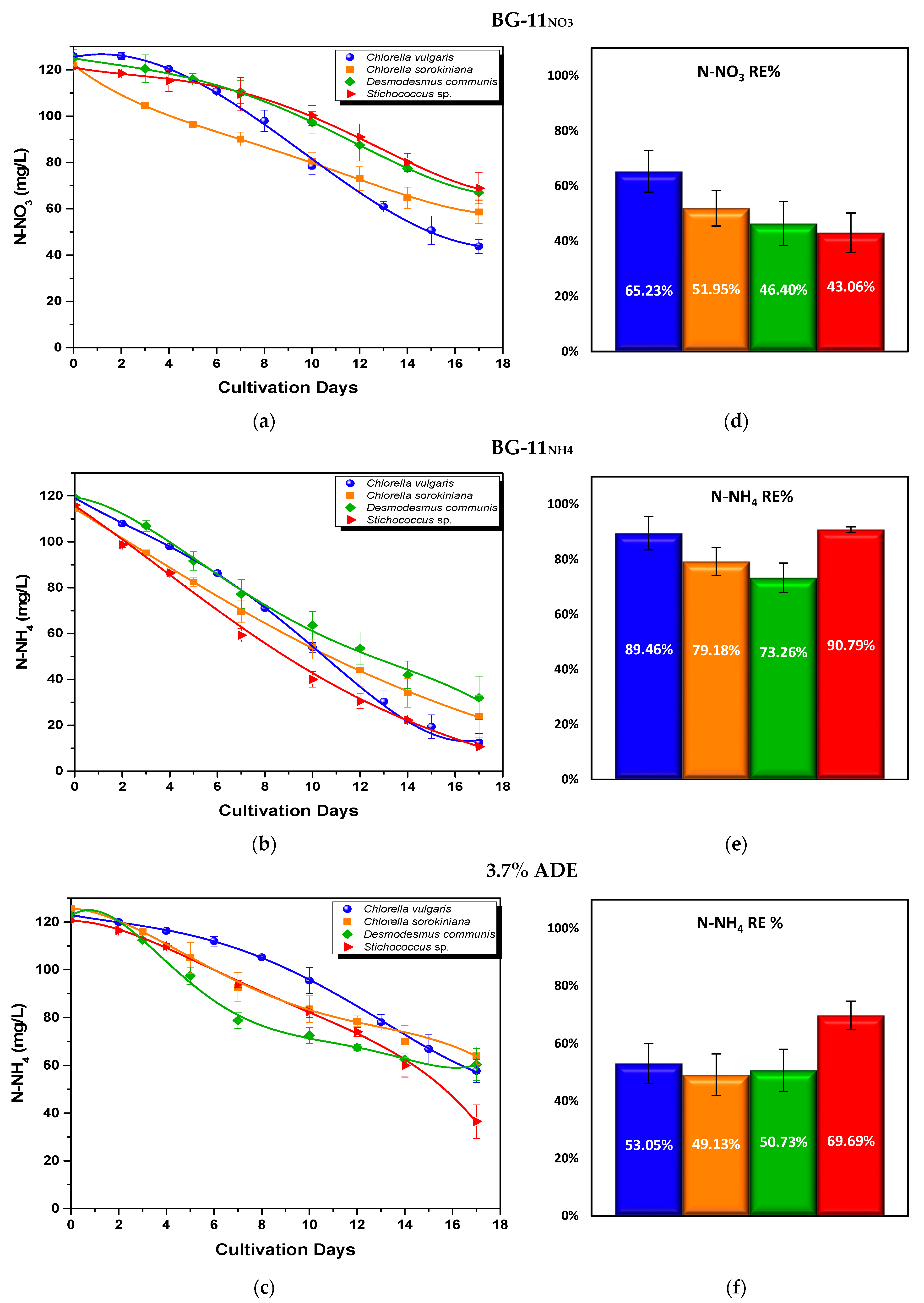
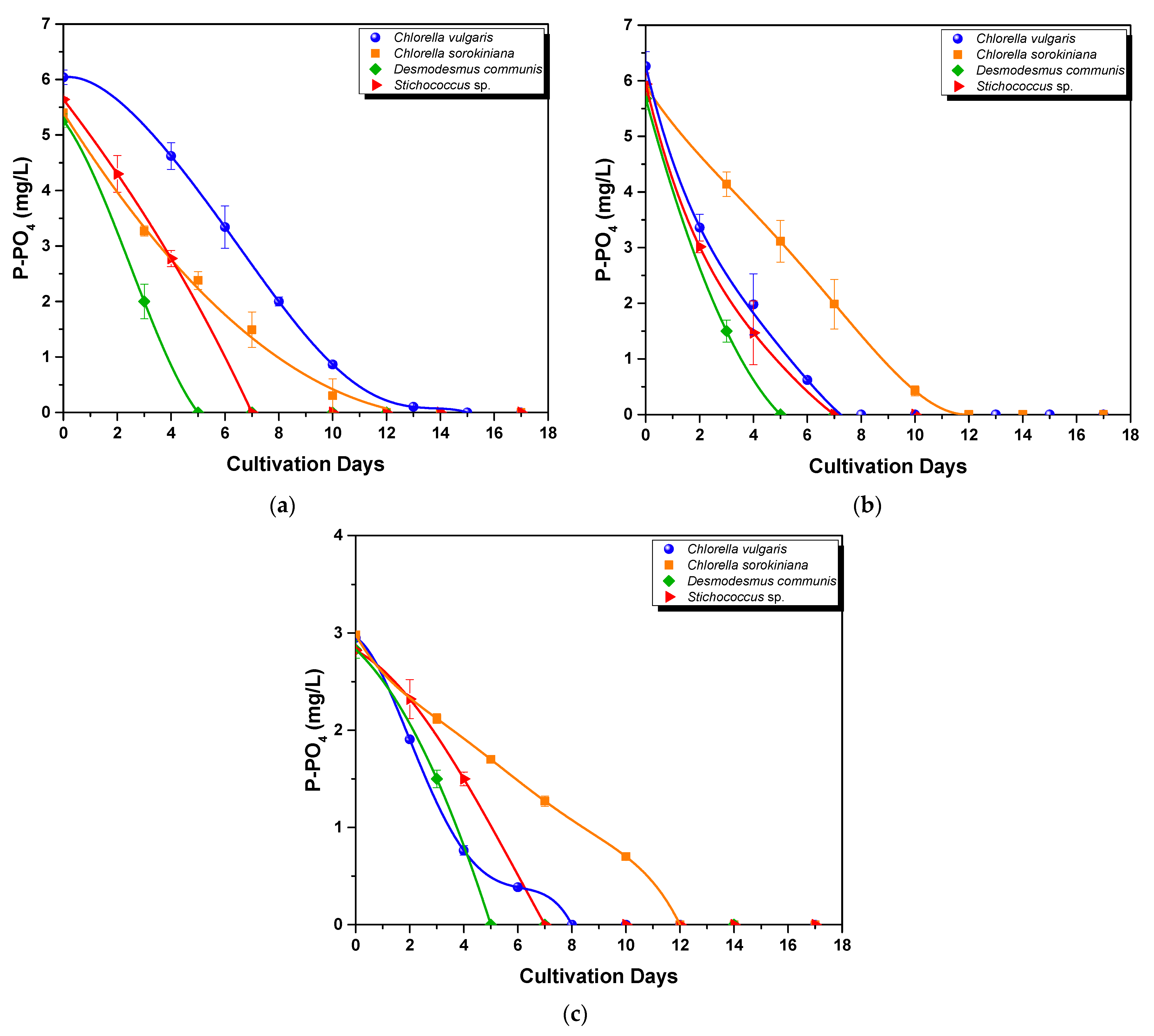
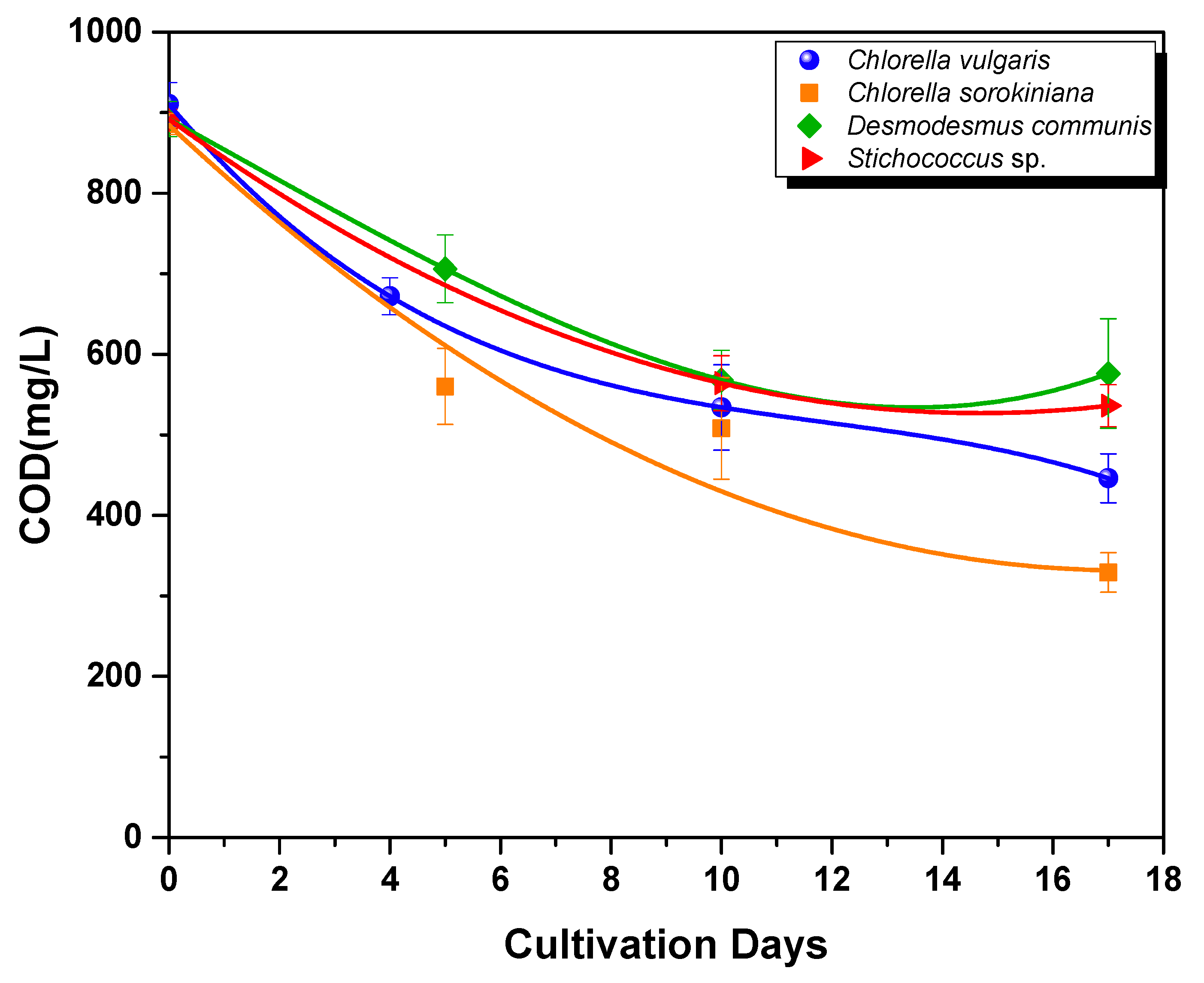
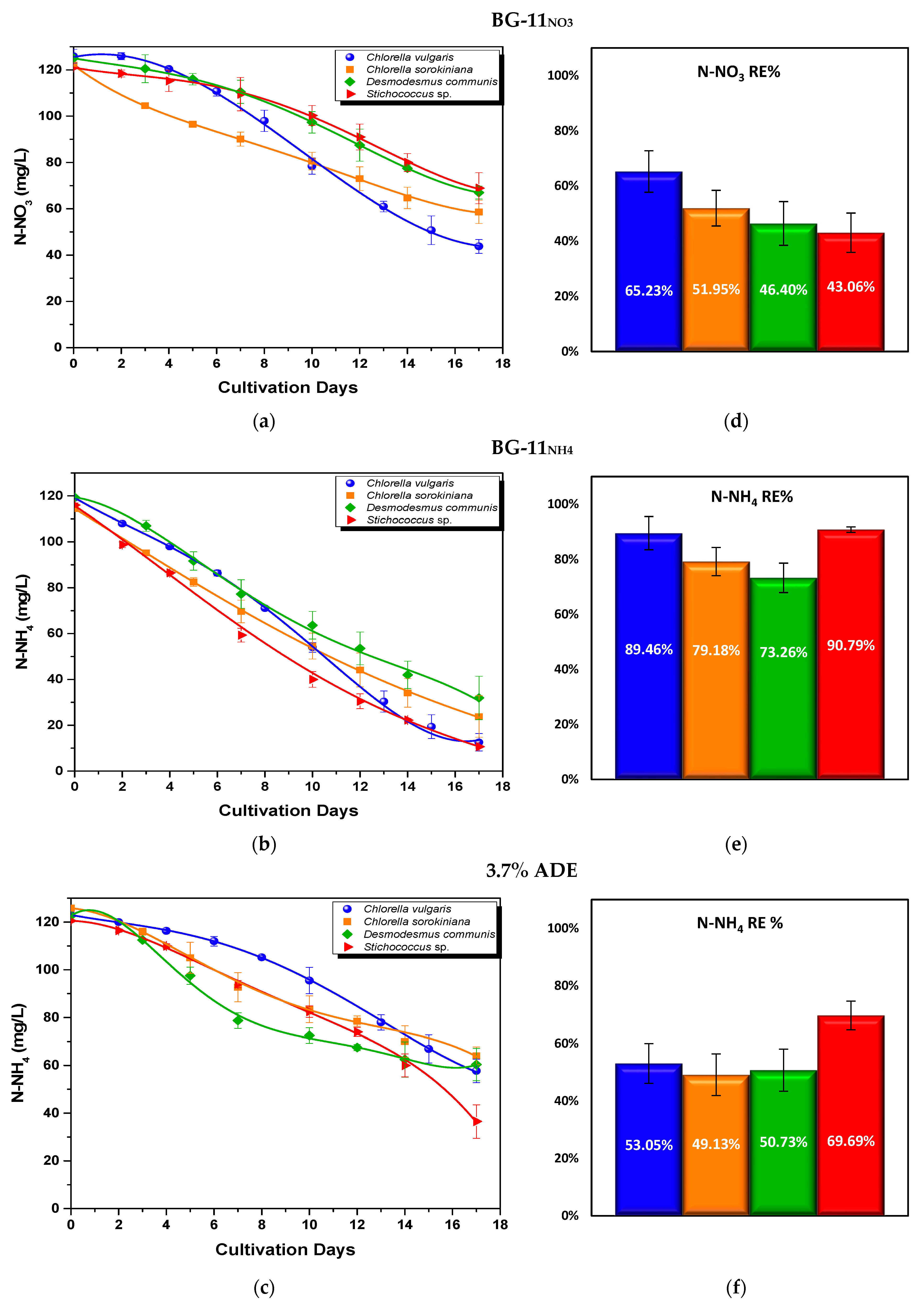
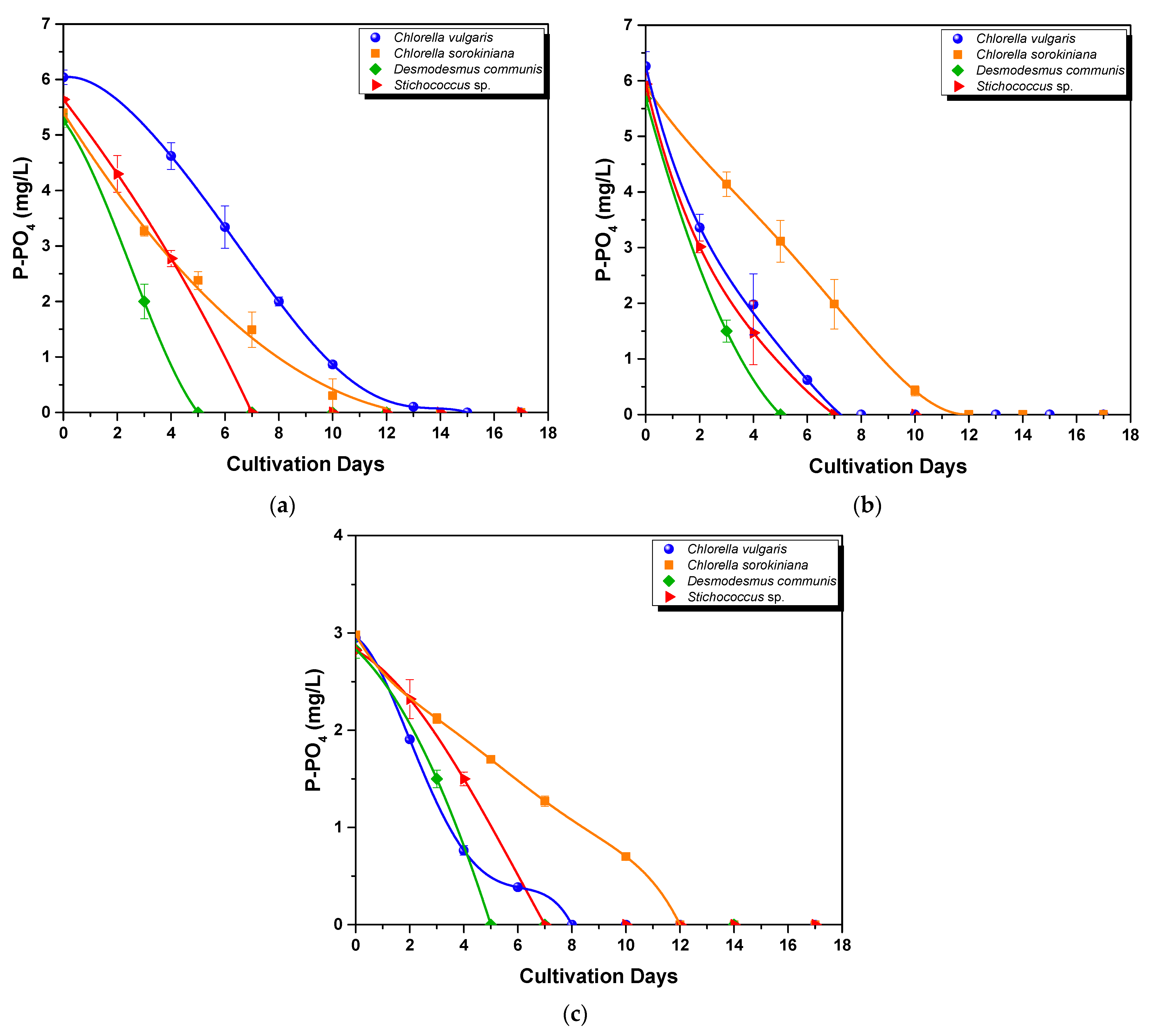
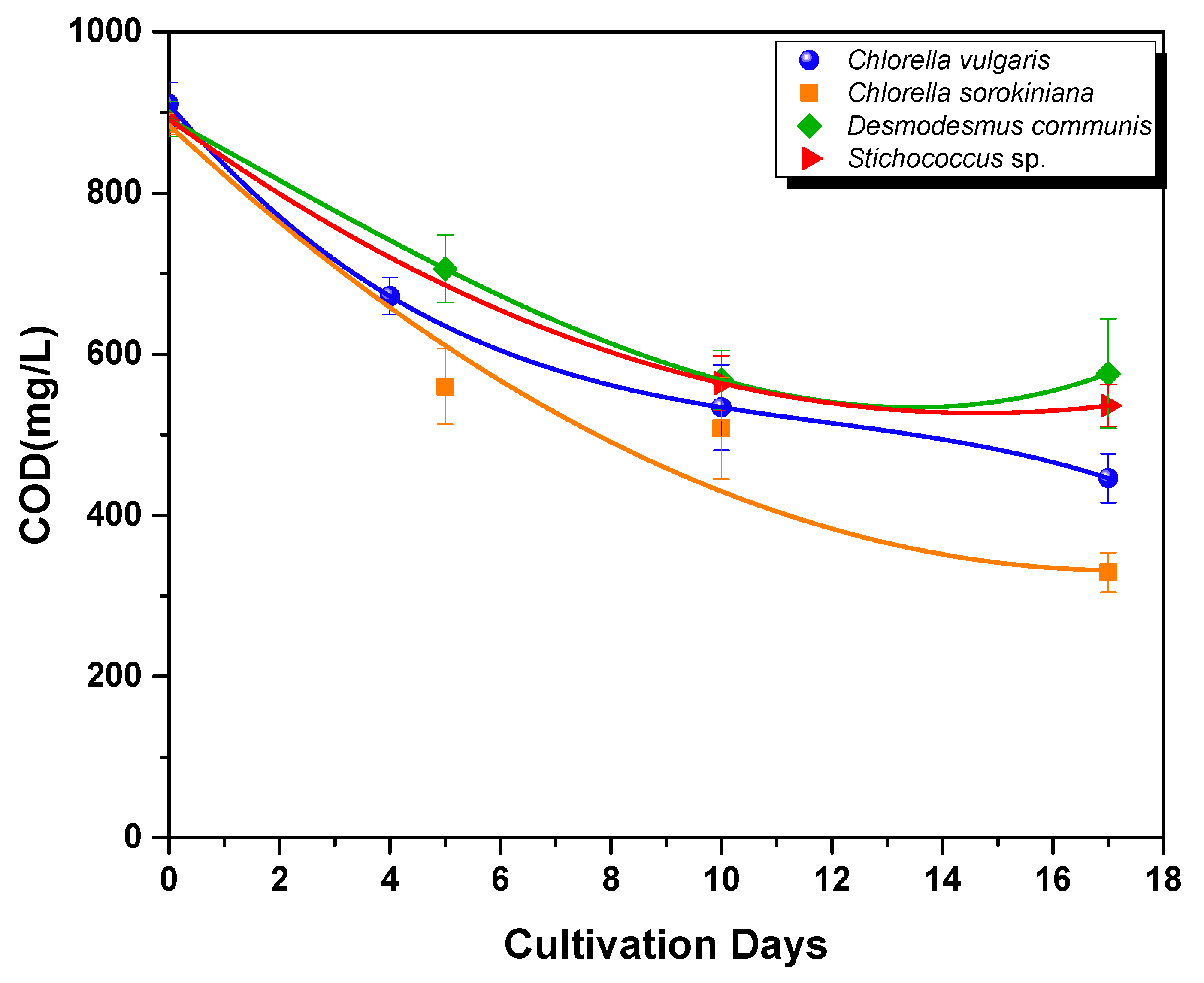
3.2.2. Nutrient Assimilation
- Nitrogen uptake
- Phosphorus uptake
- COD removal
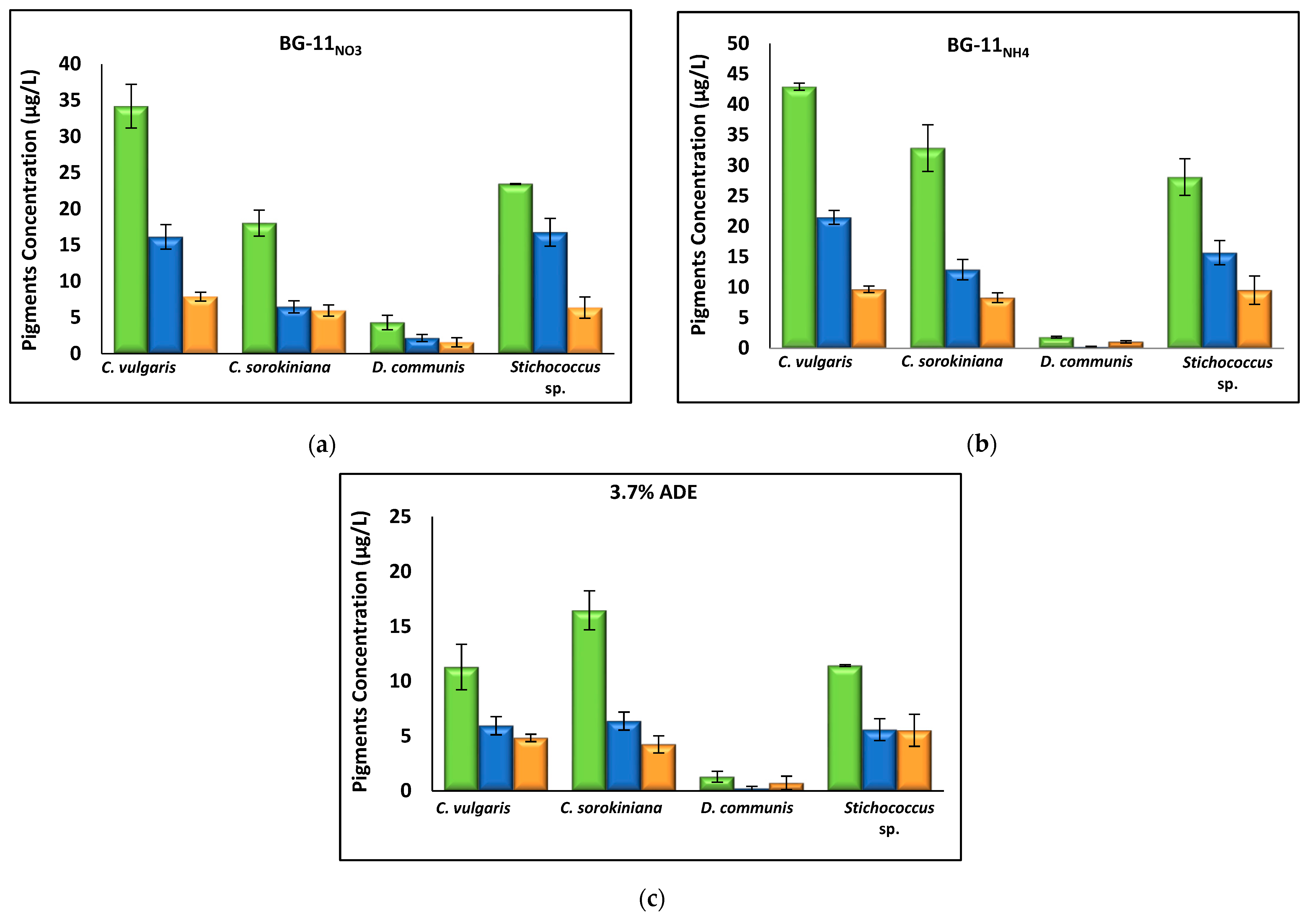
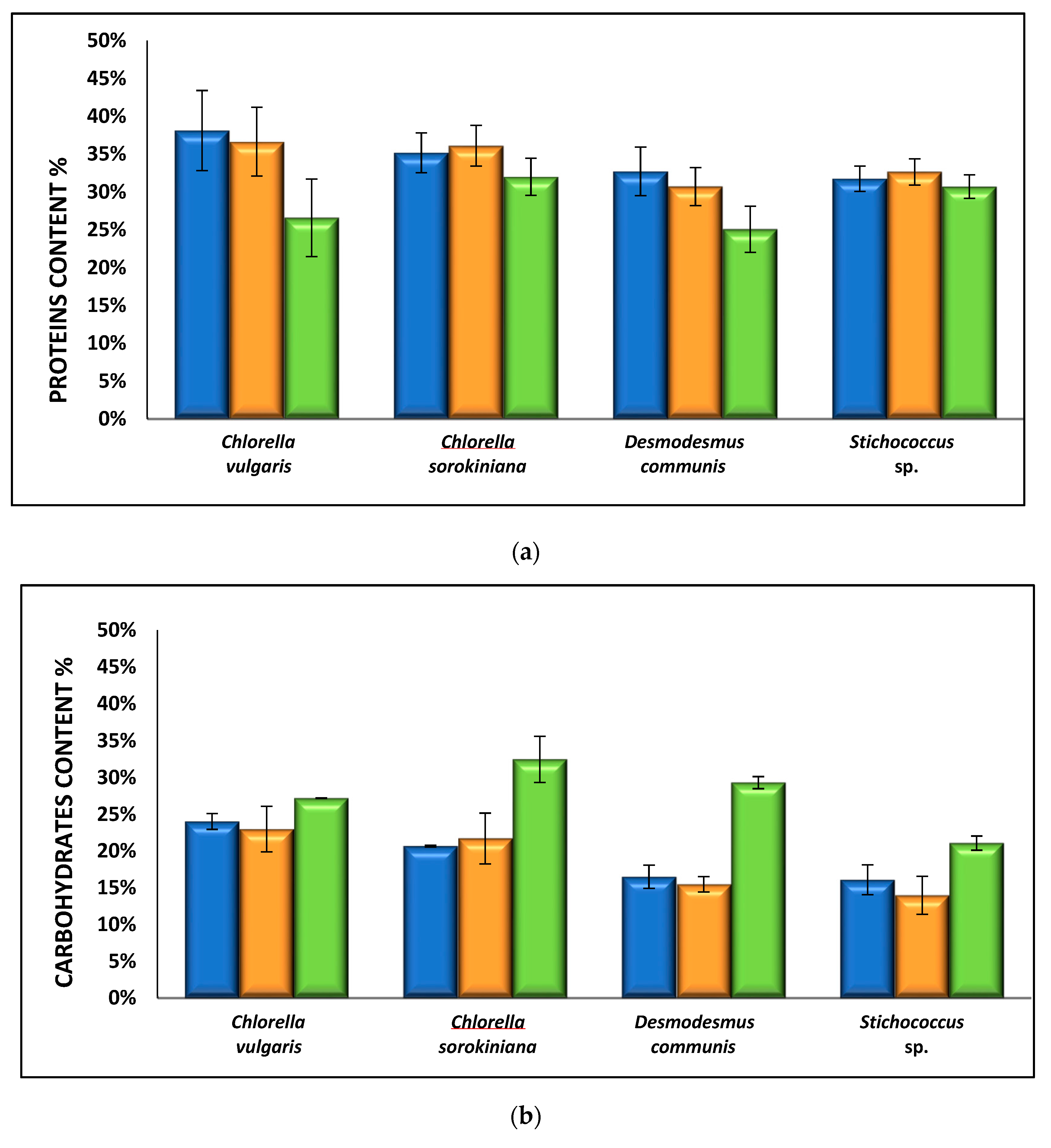
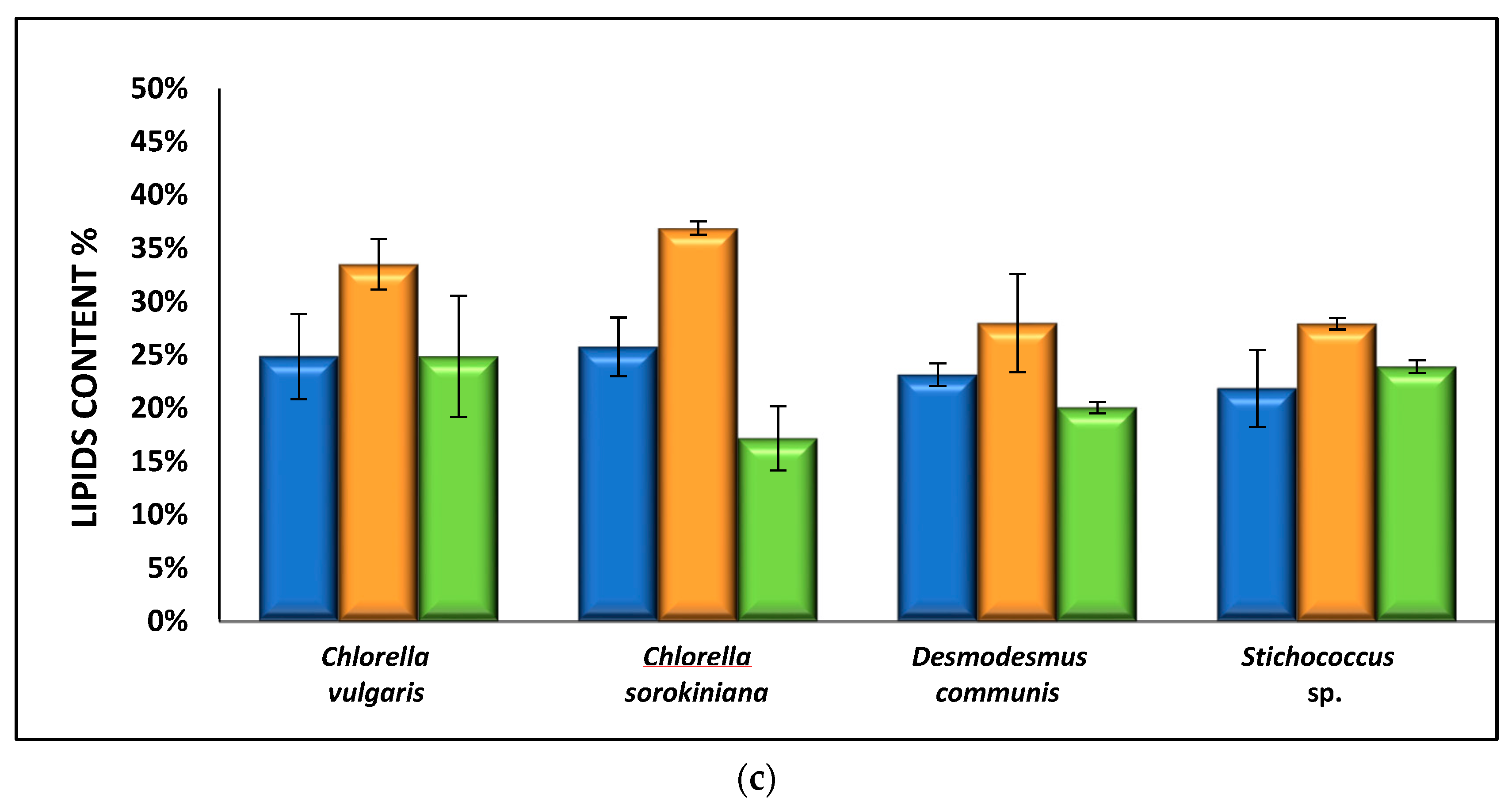
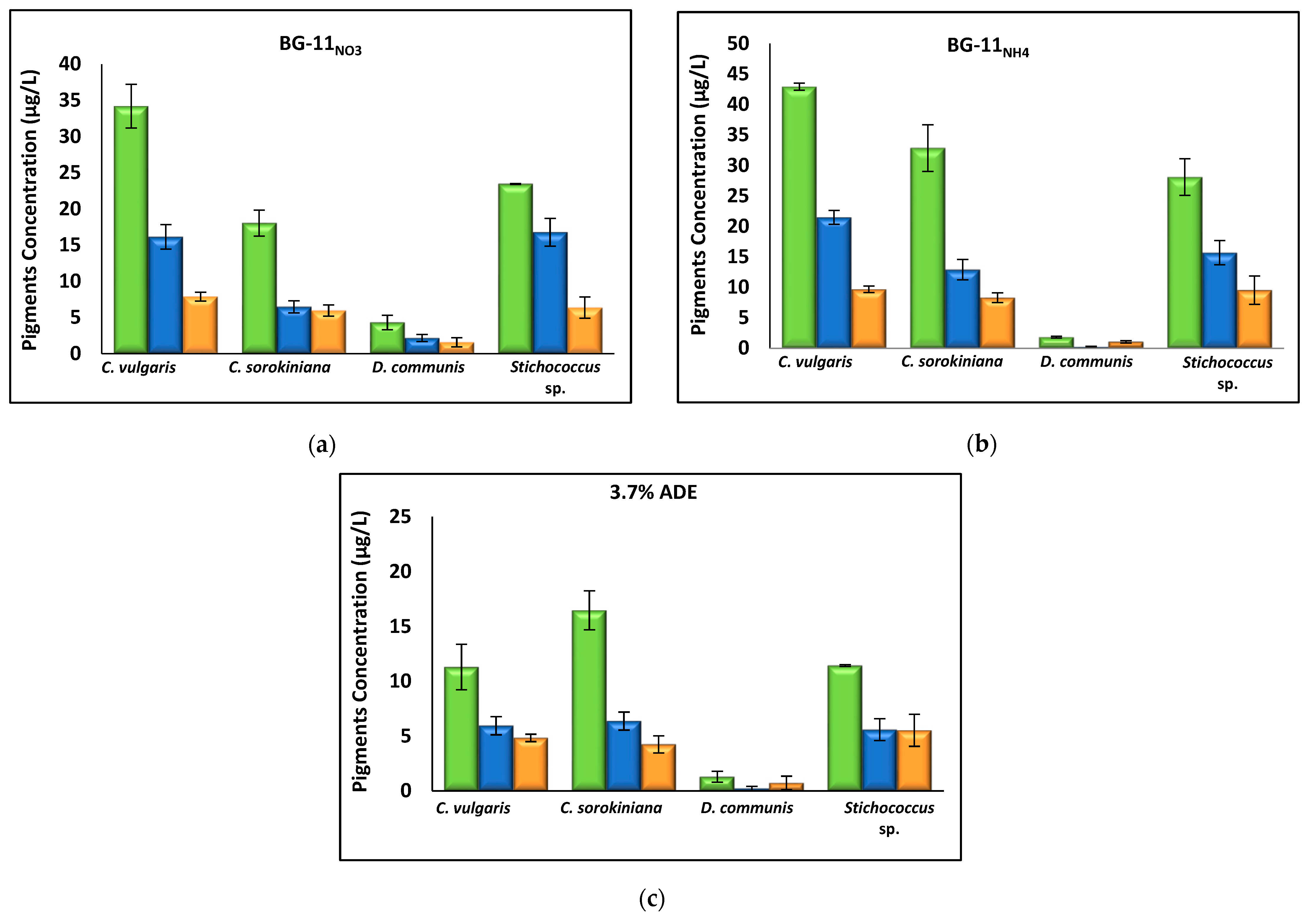
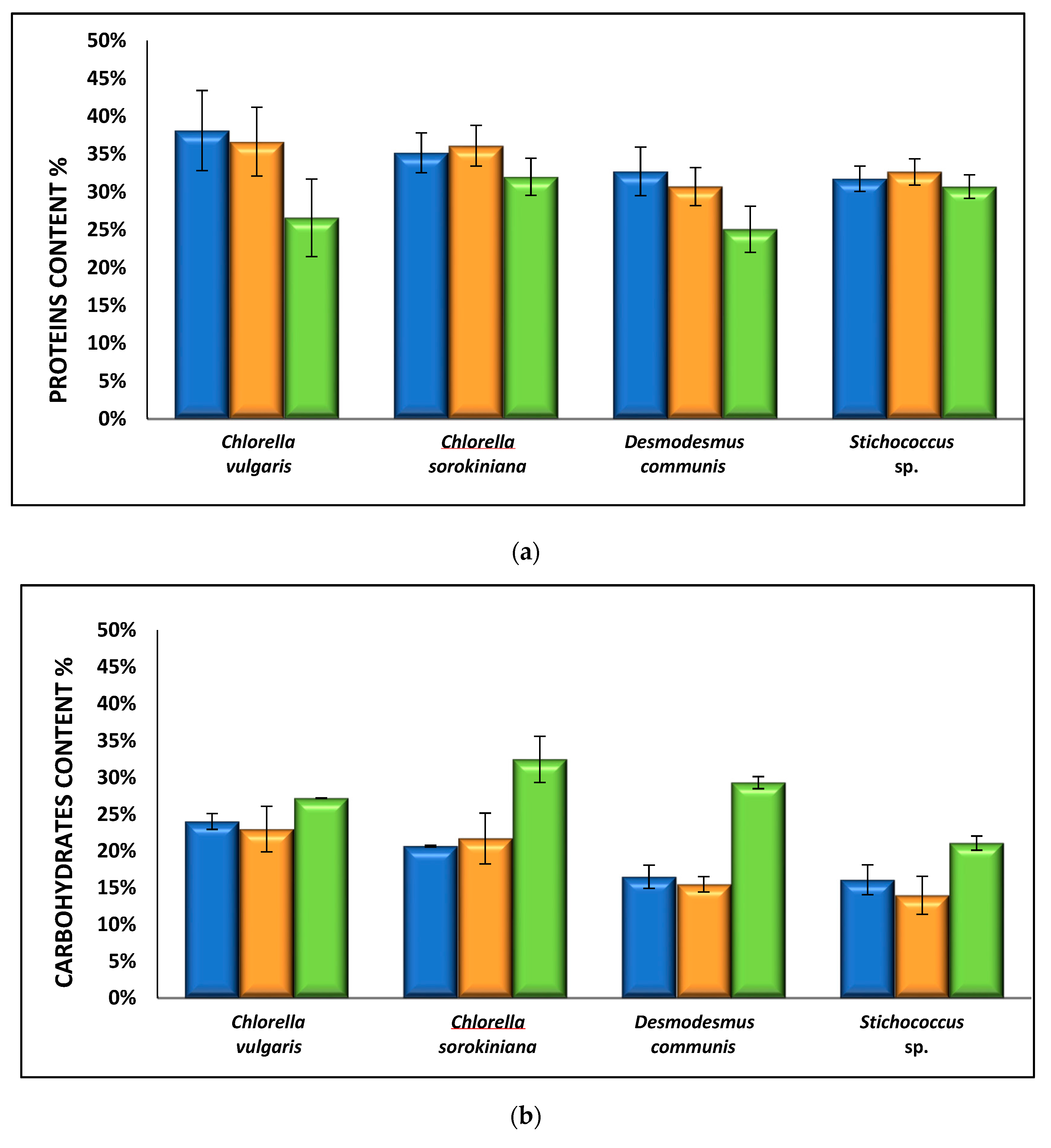
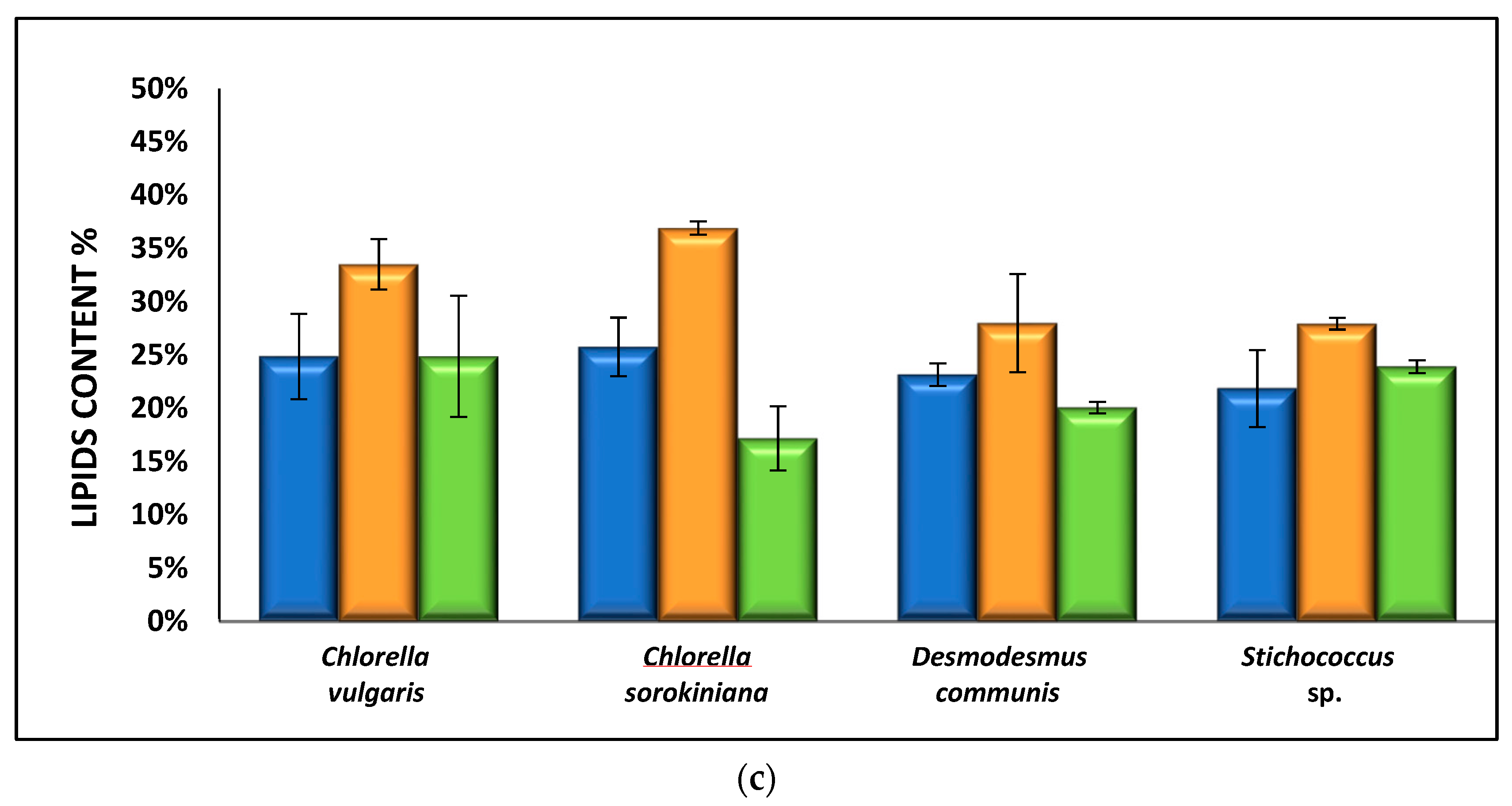
3.2.3. Effect of Nutrient Media on Biomass Composition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae Cultivation Technologies as an Opportunity for Bioenergetic System Development—Advantages and Limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Bohutskyi, P.; Liu, K.; Nasr, L.K.; Byers, N.; Rosenberg, J.N.; Oyler, G.A.; Betenbaugh, M.J.; Bouwer, E.J. Bioprospecting of Microalgae for Integrated Biomass Production and Phytoremediation of Unsterilized Wastewater and Anaerobic Digestion Centrate. Appl. Microbiol. Biotechnol. 2015, 99, 6139–6154. [Google Scholar] [CrossRef] [PubMed]
- Colling Klein, B.; Bonomi, A.; Maciel Filho, R. Integration of Microalgae Production with Industrial Biofuel Facilities: A Critical Review. Renew. Sustain. Energy Rev. 2018, 82, 1376–1392. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, R.A.; Pawar, S.B. Microalgal Bioremediation of Food-Processing Industrial Wastewater under Mixotrophic Conditions: Kinetics and Scale-up Approach. Front. Chem. Sci. Eng. 2016, 10, 499–508. [Google Scholar] [CrossRef]
- Mayers, J.J.; Ekman Nilsson, A.; Svensson, E.; Albers, E. Integrating Microalgal Production with Industrial Outputs—Reducing Process Inputs and Quantifying the Benefits. Ind. Biotechnol. 2016, 12, 219–234. [Google Scholar] [CrossRef]
- Bricker, S.B.; Longstaff, B.; Dennison, W.; Jones, A.; Boicourt, K.; Wicks, C.; Woerner, J. Effects of Nutrient Enrichment in the Nation’s Estuaries: A Decade of Change. Harmful Algae 2008, 8, 21–32. [Google Scholar] [CrossRef]
- Li, K.; Liu, Q.; Fang, F.; Luo, R.; Lu, Q.; Zhou, W.; Huo, S.; Cheng, P.; Liu, J.; Addy, M.; et al. Microalgae-Based Wastewater Treatment for Nutrients Recovery: A Review. Bioresour. Technol. 2019, 291, 121934. [Google Scholar] [CrossRef]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating micro-algae into wastewater treatment: A review. Sci. Total Environ. 2021, 752, 142168. [Google Scholar] [CrossRef]
- EBA. Statistical Report; European Biogas Association: Brussels, Belgium, 2020. [Google Scholar]
- Mayers, J.J.; Ekman Nilsson, A.; Albers, E.; Flynn, K.J. Nutrients from Anaerobic Digestion Effluents for Cultivation of the Microalga Nannochloropsis sp.—Impact on Growth, Biochemical Composition and the Potential for Cost and Environmental Impact Savings. Algal Res. 2017, 26, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, W.; Drosg, B. Assessment of the State of the Art of Technologies for the Processing of Digestate Residue from Anaerobic Digesters. Water Sci. Technol. 2013, 67, 1984–1993. [Google Scholar] [CrossRef] [Green Version]
- Logan, M.; Visvanathan, C. Management Strategies for Anaerobic Digestate of Organic Fraction of Municipal Solid Waste: Current Status and Future Prospects. Waste Manag. Res. J. Sustain. Circ. Econ. 2019, 37 (Suppl. 1), 27–39. [Google Scholar] [CrossRef] [Green Version]
- Xia, A.; Murphy, J.D. Microalgal Cultivation in Treating Liquid Digestate from Biogas Systems. Trends Biotechnol. 2016, 34, 264–275. [Google Scholar] [CrossRef]
- Kim, G.-Y.; Yun, Y.-M.; Shin, H.-S.; Han, J.-I. Cultivation of Four Microalgae Species in the Effluent of Anaerobic Digester for Biodiesel Production. Bioresour. Technol. 2017, 224, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, J.; Herbes, C.; Nelles, M. Biogas Digestate Marketing: Qualitative Insights into the Supply Side. Resour. Conserv. Recycl. 2015, 104, 152–161. [Google Scholar] [CrossRef]
- Plana, P.V.; Noche, B. A Review of the Current Digestate Distribution Models: Storage and Transport. WIT Trans. Ecol. Environ. 2016, 202, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Dębowski, M.; Zieliński, M.; Kisielewska, M.; Kazimierowicz, J.; Dudek, M.; Świca, I.; Rudnicka, A. The cultivation of lipid-rich microalgae biomass as anaerobic digestate valorization technology—A pilot-scale study. Processes 2020, 8, 517. [Google Scholar] [CrossRef]
- Dębowski, M.; Kisielewska, M.; Kazimierowicz, J.; Rudnicka, A.; Dudek, M.; Romanowska-Duda, Z.; Zieliński, M. The effects of Microalgae Biomass Co-Substrate on Biogas Production from the Common Agricultural Biogas Plants Feedstock. Energies 2020, 13, 2186. [Google Scholar] [CrossRef]
- Bauer, L.; Ranglová, K.; Masojídek, J.; Drosg, B.; Meixner, K. Digestate as Sustainable Nutrient Source for Microalgae—Challenges and Prospects. Appl. Sci. 2021, 11, 1056. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Ignacio Gayo-Peláez, J.; Ndovela, V.; Wood, E.; Vijay Kapoore, R.; Anne Llewellyn, C. Towards a Circular Economy: A Novel Microalgal Two-Step Growth Approach to Treat Excess Nutrients from Digestate and to Produce Biomass for Animal Feed. Bioresour. Technol. 2021, 320, 124349. [Google Scholar] [CrossRef] [PubMed]
- Morales-Amaral, M.d.M.; Gómez-Serrano, C.; Acién, F.G.; Fernández-Sevilla, J.M.; Molina-Grima, E. Production of Microalgae Using Centrate from Anaerobic Digestion as the Nutrient Source. Algal Res. 2015, 9, 297–305. [Google Scholar] [CrossRef]
- Singh, M.; Reynolds, D.L.; Das, K.C. Microalgal System for Treatment of Effluent from Poultry Litter Anaerobic Digestion. Bioresour. Technol. 2011, 102, 10841–10848. [Google Scholar] [CrossRef]
- Zuliani, L.; Frison, N.; Jelic, A.; Fatone, F.; Bolzonella, D.; Ballottari, M. Microalgae Cultivation on Anaerobic Digestate of Municipal Wastewater, Sewage Sludge and Agro-Waste. Int. J. Mol. Sci. 2016, 17, 1692. [Google Scholar] [CrossRef]
- Lortou, U.; Gkelis, S. Polyphasic Taxonomy of Green Algae Strains Isolated from Mediterranean Freshwaters. J. Biol. Res.-Thessalon. 2019, 26, 11. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Panou, M. Capturing Biodiversity: Linking a Cyanobacteria Culture Collection to the “Scratchpads” Virtual Research Environment Enhances Biodiversity Knowledge. Biodivers. Data J. 2016, 4, e7965-1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eustance, E.; Gardner, R.D.; Moll, K.M.; Menicucci, J.; Gerlach, R.; Peyton, B.M. Growth, Nitrogen Utilization and Biodiesel Potential for Two Chlorophytes Grown on Ammonium, Nitrate or Urea. J. Appl. Phycol. 2013, 25, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Henriques, M.; Silva, A.; Rocha, J. Extraction and Quantification of Pigments from a Marine Microalga: A Simple and Reproducible Method. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. Formatex 2007, 2, 586–593. [Google Scholar]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Schwenzfeier, A.; Wierenga, P.A.; Gruppen, H. Isolation and Characterization of Soluble Protein from the Green Microalgae Tetraselmis sp. Bioresour. Technol. 2011, 102, 9121–9127. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and Wastewater Treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madakka, M.; Jayaraju, N.; Rajesh, N.; Subhosh Chandra, M.R.G. Development in the Treatment of Municipal and Industrial Wastewater by Microorganism. In Recent Developments in Applied Microbiology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 263–273. [Google Scholar] [CrossRef]
- Shahid, A.; Malik, S.; Zhu, H.; Xu, J.; Nawaz, M.Z.; Nawaz, S.; Alam, M.A.; Mehmood, M.A. Cultivating microalgae in wastewater for biomass production, pollutant removal, and atmospheric carbon mitigation; a review. Sci. Total Environ. 2020, 704, 135303. [Google Scholar] [CrossRef] [PubMed]
- Psachoulia, P.; Chatzidoukas, C. Illumination Policies for Stichococcus sp. Cultures in an Optimally Operating Lab-Scale PBR toward the Directed Photosynthetic Production of Desired Products. Sustainability 2021, 13, 2489. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P.J. Acclimation and Toxicity of High Ammonium Concentrations to Unicellular Algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Kwan, T.A.; Zimmerman, J.B.; Peccia, J. Ammonia Inhibition in Oleaginous Microalgae. Algal Res. 2016, 19, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J. Growth Optimisation of Microalga Mutant at High CO2 Concentration to Purify Undiluted Anaerobic Digestion Effluent of Swine Manure. Bioresour. Technol. 2015, 177, 240–246. [Google Scholar] [CrossRef]
- Franchino, M.; Comino, E.; Bona, F.; Riggio, V.A. Growth of Three Microalgae Strains and Nutrient Removal from an Agro-Zootechnical Digestate. Chemosphere 2013, 92, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Marjakangas, J.M.; Chen, C.-Y.; Lakaniemi, A.-M.; Puhakka, J.A.; Whang, L.-M.; Chang, J.-S. Selecting an Indigenous Microalgal Strain for Lipid Production in Anaerobically Treated Piggery Wastewater. Bioresour. Technol. 2015, 191, 369–376. [Google Scholar] [CrossRef]
- Salbitani, G.; Carfagna, S. Ammonium Utilization in Microalgae: A Sustainable Method for Wastewater Treatment. Sustainability 2021, 13, 956. [Google Scholar] [CrossRef]
- Lachmann, S.C.; Mettler-Altmann, T.; Wacker, A.; Spijkerman, E. Nitrate or Ammonium: Influences of Nitrogen Source on the Physiology of a Green Alga. Ecol. Evol. 2019, 9, 1070–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tale, M.; Ghosh, S.; Kapadnis, B.; Kale, S. Isolation and Characterization of Microalgae for Biodiesel Production from Nisargruna Biogas Plant Effluent. Bioresour. Technol. 2014, 169, 328–335. [Google Scholar] [CrossRef]
- Ji, F.; Zhou, Y.; Pang, A.; Ning, L.; Rodgers, K.; Liu, Y.; Dong, R. Fed-Batch Cultivation of Desmodesmus sp. in Anaerobic Digestion Wastewater for Improved Nutrient Removal and Biodiesel Production. Bioresour. Technol. 2015, 184, 116–122. [Google Scholar] [CrossRef]
- Makaroglou, G.; Marakas, H.; Fodelianakis, S.; Axaopoulou, V.A.; Koumi, I.; Kalogerakis, N.; Gikas, P. Optimization of Biomass Production from Stichococcous sp. Biofilms Coupled to Wastewater Treatment. Biochem. Eng. J. 2021, 169, 107964. [Google Scholar] [CrossRef]
- Safonova, E.; Kvitko, K.V.; Iankevitch, M.I.; Surgko, L.F.; Afti, I.A.; Reisser, W. Biotreatment of Industrial Wastewater by Selected Algal-Bacterial Consortia. Eng. Life Sci. 2004, 4, 347–353. [Google Scholar] [CrossRef]
- Cai, T.; Park, S.Y.; Li, Y. Nutrient Recovery from Wastewater Streams by Microalgae: Status and Prospects. Renew. Sustain. Energy Rev. 2013, 19, 360–369. [Google Scholar] [CrossRef]
- Markou, G.; Angelidaki, I.; Georgakakis, D. Microalgal Carbohydrates: An Overview of the Factors Influencing Carbohydrates Production, and of Main Bioconversion Technologies for Production of Biofuels. Appl. Microbiol. Biotechnol. 2012, 96, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. Removal of Nitrogen and Phosphorus by Chlorella Sorokiniana Cultured Heterotrophically in Ammonia and Nitrate. Int. Biodeterior. 2013, 85, 511–516. [Google Scholar] [CrossRef]
- Xin, L.; Hong-ying, H.; Ke, G.; Jia, Y. Growth and Nutrient Removal Properties of a Freshwater Microalga Scenedesmus sp. LX1 under Different Kinds of Nitrogen Sources. Ecol. Eng. 2010, 36, 379–381. [Google Scholar] [CrossRef]
- Kumar, A.; Bera, S. Revisiting Nitrogen Utilization in Algae: A Review on the Process of Regulation and Assimilation. Bioresour. Technol. Rep. 2020, 12, 100584. [Google Scholar] [CrossRef]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of Nitrogen Sources on Cell Growth and Biochemical Composition of Marine Chlorophyte Tetraselmis sp. for Lipid Production. Algae 2016, 31, 257–266. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic Cultures of Microalgae: Metabolism and Potential Products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Noel, E.A.; Barnes, A.; Watson, A.; Rosenberg, J.N.; Erickson, G.; Oyler, G.A. Characterization of Three Chlorella Sorokiniana Strains in Anaerobic Digested Effluent from Cattle Manure. Bioresour. Technol. 2013, 150, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielinski, M.; Dębowski, M.; Szwaja, S.; Kisielewska, M. Anaerobic Digestion Effluents (ADEs) Treatment Coupling with Chlorella sp. Microalgae Production. Water Environ. Res. 2018, 90, 155–163. [Google Scholar] [CrossRef]
- Ji, F.; Liu, Y.; Hao, R.; Li, G.; Zhou, Y.; Dong, R. Biomass Production and Nutrients Removal by a New Microalgae Strain Desmodesmus sp. in Anaerobic Digestion Wastewater. Bioresour. Technol. 2014, 161, 200–207. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, X.; Zou, G.; Zhou, T.; Liu, Y.; Ruan, R. Cultivation of Chlorella Vulgaris in Manure-Free Piggery Wastewater with High-Strength Ammonium for Nutrients Removal and Biomass Production: Effect of Ammonium Concentration, Carbon/Nitrogen Ratio and PH. Bioresour. Technol. 2019, 273, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Chuka-ogwude, D.; Ogbonna, J.; Borowitzka, M.A.; Moheimani, N.R. Screening, Acclimation and Ammonia Tolerance of Microalgae Grown in Food Waste Digestate. J. Appl. Phycol. 2020, 32, 3775–3785. [Google Scholar] [CrossRef]
- Raeisossadati, M.; Vadiveloo, A.; Bahri, P.A.; Parlevliet, D.; Moheimani, N.R. Treating Anaerobically Digested Piggery Effluent (ADPE) Using Microalgae in Thin Layer Reactor and Raceway Pond. J. Appl. Phycol. 2019, 31, 2311–2319. [Google Scholar] [CrossRef]
- González, C.; Marciniak, J.; Villaverde, S.; León, C.; García, P.A.; Muñoz, R. Efficient Nutrient Removal from Swine Manure in a Tubular Biofilm Photo-Bioreactor Using Algae-Bacteria Consortia. Water Sci. Technol. 2008, 58, 95–102. [Google Scholar] [CrossRef]
- Vadiveloo, A.; Nwoba, E.G.; Moheimani, N.R. Viability of Combining Microalgae and Macroalgae Cultures for Treating Anaerobically Digested Piggery Effluent. J. Environ. Sci. 2019, 82, 132–144. [Google Scholar] [CrossRef]
- Benítez, M.B.; Champagne, P.; Ramos, A.; Torres, A.F.; Ochoa-Herrera, V. Wastewater Treatment for Nutrient Removal with Ecuadorian Native Microalgae. Environ. Technol. 2019, 40, 2977–2985. [Google Scholar] [CrossRef] [PubMed]
- Min, M.; Wang, L.; Li, Y.; Mohr, M.J.; Hu, B.; Zhou, W.; Chen, P.; Ruan, R. Cultivating Chlorella sp. in a Pilot-Scale Photobioreactor Using Centrate Wastewater for Microalgae Biomass Production and Wastewater Nutrient Removal. Appl. Biochem. Biotechnol. 2011, 165, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Tan, F. Nitrogen and Phosphorus Removal Coupled with Carbohydrate Production by Five Microalgae Cultures Cultivated in Biogas Slurry. Bioresour. Technol. 2016, 221, 385–393. [Google Scholar] [CrossRef]
- Liu, X.; Fujiwara, M.; Kodera, T.; Watanabe, K.; Akizuki, S.; Kishi, M.; Koyama, M.; Toda, T.; Ban, S. Conditions for Continuous Cultivation of Chlorella Sorokiniana and Nutrient Removal from Anaerobic Digestion Effluent of Aquatic Macrophytes. Int. Biodeterior. Biodegrad. 2020, 149, 104923. [Google Scholar] [CrossRef]
- Choi, H.J.; Lee, S.M. Effect of the N/P Ratio on Biomass Productivity and Nutrient Removal from Municipal Wastewater. Bioprocess Biosyst. Eng. 2015, 38, 761–766. [Google Scholar] [CrossRef]
- Markou, G. Fed-Batch Cultivation of Arthrospira and Chlorella in Ammonia-Rich Wastewater: Optimization of Nutrient Removal and Biomass Production. Bioresour. Technol. 2015, 193, 35–41. [Google Scholar] [CrossRef]
- Xie, B.; Gong, W.; Tian, Y.; Qu, F.; Luo, Y.; Du, X.; Tang, X.; Xu, D.; Lin, D.; Li, G.; et al. Biodiesel Production with the Simultaneous Removal of Nitrogen, Phosphorus and COD in Microalgal-Bacterial Communities for the Treatment of Anaerobic Digestion Effluent in Photobioreactors. Chem. Eng. J. 2018, 350, 1092–1102. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, S.; Hu, C.; Zhang, H.; Xu, J.; Ping, L. Performance of Three Microalgal Strains in Biogas Slurry Purification and Biogas Upgrade in Response to Various Mixed Light-Emitting Diode Light Wavelengths. Bioresour. Technol. 2015, 187, 338–345. [Google Scholar] [CrossRef]
- Gupta, S.; Pawar, S.B. An Integrated Approach for Microalgae Cultivation Using Raw and Anaerobic Digested Wastewaters from Food Processing Industry. Bioresour. Technol. 2018, 269, 571–576. [Google Scholar] [CrossRef]
- Sánchez-Bayo, A.; Morales, V.; Rodríguez, R.; Vicente, G.; Bautista, L.F. Cultivation of Microalgae and Cyanobacteria: Effect of Operating Conditions on Growth and Biomass Composition. Molecules 2020, 25, 2834. [Google Scholar] [CrossRef]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A Review on the Assessment of Stress Conditions for Simultaneous Production of Microalgal Lipids and Carotenoids. Front. Microbiol. 2016, 7, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roopnarain, A.; Sym, S.; Gray, V.M. Effect of Nitrogenous Resource on Growth, Biochemical Composition and Ultrastructure of Isochrysis Galbana (Isochrysidales, Haptophyta): Effect of N Source on I. Galbana. Phycol. Res. 2015, 63, 43–50. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | BG-11NO3 | BG-11NH4 | 3.7% ADE | Raw ADE |
|---|---|---|---|---|
| N-NH4 (mg/L) | ND | 123.92 | 124 | 3351.35 |
| Ν-ΝO3 (mg/L) | 123.92 | ND | 3.3 | 89.19 |
| TN (mg/L) | 123.92 | 123.92 | 170 | 4594.6 |
| P (mg/L) | 5.50 | 5.50 | 3 | 81.1 |
| COD (mg/L) | ND | ND | 895.63 | 24,200 |
| BOD (mg/L) | ND | ND | 238 | 6430 |
| pH | 7.8 | 8 | 8.2 | 8.6 |
| K (mg/L) | 13.70 | 13.70 | 116.96 | 3161 |
| Ca (mg/L) | 9.81 | 9.81 | 13.66 | 369.1 |
| Na (mg/L) | 212.28 | 9.41 | 69.73 | 1884.57 |
| Fe (mg/L) | 1.28 | 1.28 | 2.00 | 54.24 |
| Cl (mg/L) | 18.02 | 331.58 | 60.44 | 1633.6 |
| Mg (mg/L) | 6.98 | 6.98 | 8.3 | 225.14 |
| Mn (mg/L) | 0.50 | 0.50 | 0.23 | 6.32 |
| Zn (mg/L) | 0.11 | 0.11 | 0.42 | 11.26 |
| Cu (mg/L) | 0.02 | 0.02 | 0.075 | 2.03 |
| S (mg/L) | 9.82 | 9.82 | NM | NM |
| Co (mg/L) | 0.01 | 0.01 | NM | NM |
| Mo (mg/L) | 0.16 | 0.16 | NM | NM |
| B (mg/L) | 0.50 | 0.50 | NM | NM |
| Citric acid (mg/L) | 6.00 | 6.00 | ND | ND |
| PIPES (g/L) | ND | 12 | ND | ND |
| BG-11NO3 | BG-11NH4 | 3.7% ADE | |
|---|---|---|---|
| Initial OD600nm | |||
| C. vulgaris | 0.34 ± 0.00 | 0.34 ± 0.00 | 0.91 ± 0.01 |
| C. sorokiniana | 0.35 ± 0.01 | 0.35 ± 0.00 | 0.91 ± 0.01 |
| D. communis | 0.35 ± 0.01 | 0.35 ± 0.01 | 0.88 ± 0.00 |
| Stichococcus sp. | 0.37 ± 0.01 | 0.37 ± 0.01 | 0.97 ± 0.01 |
| Initial DCW (g/L) | |||
| C. vulgaris | 0.12 ± 0.00 | 0.12 ± 0.00 | 0.12 ± 0.00 |
| C. sorokiniana | 0.11 ± 0.00 | 0.11 ± 0.00 | 0.11 ± 0.00 |
| D. communis | 0.27 ± 0.01 | 0.27 ± 0.01 | 0.27 ± 0.01 |
| Stichococcus sp. | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.14 ± 0.01 |
| Initial cell number (cells/mL) | |||
| C. vulgaris | 4.17 × 106 ± 3.16 × 105 | 4.21 × 106 ± 3.05 × 105 | 4.09 × 106 ± 2.22 × 105 |
| C. sorokiniana | 4.36 × 106 ± 1.21 × 105 | 4.13 × 106 ± 2.16 × 105 | 4.32 × 106 ± 1.19 × 105 |
| D. communis | 3.367 × 105 ± 5.92 × 104 | 3.561 × 105 ± 1.98 × 104 | 3.489 × 105 ± 3.01 × 104 |
| Stichococcus sp. | 3.47 × 106 ± 2.17 × 105 | 3.29 × 106 ± 1.77 × 105 | 3.52 × 106 ± 1.09 × 105 |
| C. vulgaris | C. sorokiniana | D. communis | Stichococcus sp. | |||||
|---|---|---|---|---|---|---|---|---|
| Nitrogen (mg/L) | Day 0 | Day 17 | Day 0 | Day 17 | Day 0 | Day 17 | Day 0 | Day 17 |
| ΤΝ | 169 ± 1.42 | 104.25 ± 14.01 | 176 ± 0.97 | 114.5 ± 15.81 | 177 ± 0.56 | 115 ± 13.45 | 186 ± 1.19 | 102 ± 10.09 |
| N-NH4 (mg/L) | 122.94 ± 0.25 | 57.72 ± 12.03 | 125.8 ± 0.69 | 64 ± 13.77 | 122.6 ± 0.34 | 60.4 ± 12.53 | 120.5 ± 0.14 | 36.52 ± 8.72 |
| N-NO3 (mg/L) | 18.06 ± 0.11 | 18 ± 0.10 | 15.5 ± 0.04 | 15.8 ± 0.14 | 24.1 ± 0.02 | 24.1 ± 0.06 | 34.9 ± 0.03 | 36 ± 0.72 |
| N-organic | 28 ± 1.50 | 28.53 ± 2.65 | 34.7 ± 0.34 | 34.7 ± 3.08 | 30.3 ± 0.34 | 30.5 ± 1.22 | 30.6 ± 1.44 | 29.48 ± 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Psachoulia, P.; Schortsianiti, S.-N.; Lortou, U.; Gkelis, S.; Chatzidoukas, C.; Samaras, P. Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent. Water 2022, 14, 221. https://doi.org/10.3390/w14020221
Psachoulia P, Schortsianiti S-N, Lortou U, Gkelis S, Chatzidoukas C, Samaras P. Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent. Water. 2022; 14(2):221. https://doi.org/10.3390/w14020221
Chicago/Turabian StylePsachoulia, Paraskevi, Sofia-Natalia Schortsianiti, Urania Lortou, Spyros Gkelis, Christos Chatzidoukas, and Petros Samaras. 2022. "Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent" Water 14, no. 2: 221. https://doi.org/10.3390/w14020221
APA StylePsachoulia, P., Schortsianiti, S.-N., Lortou, U., Gkelis, S., Chatzidoukas, C., & Samaras, P. (2022). Assessment of Nutrients Recovery Capacity and Biomass Growth of Four Microalgae Species in Anaerobic Digestion Effluent. Water, 14(2), 221. https://doi.org/10.3390/w14020221