
Gone and Back—The Anthropogenic History of Coccotylus brodiei (Turner) Kützing and Furcellaria lumbricalis (Hudson) J.V. Lamouroux in the Gulf of Gdańsk (Southern Baltic Sea)


Abstract
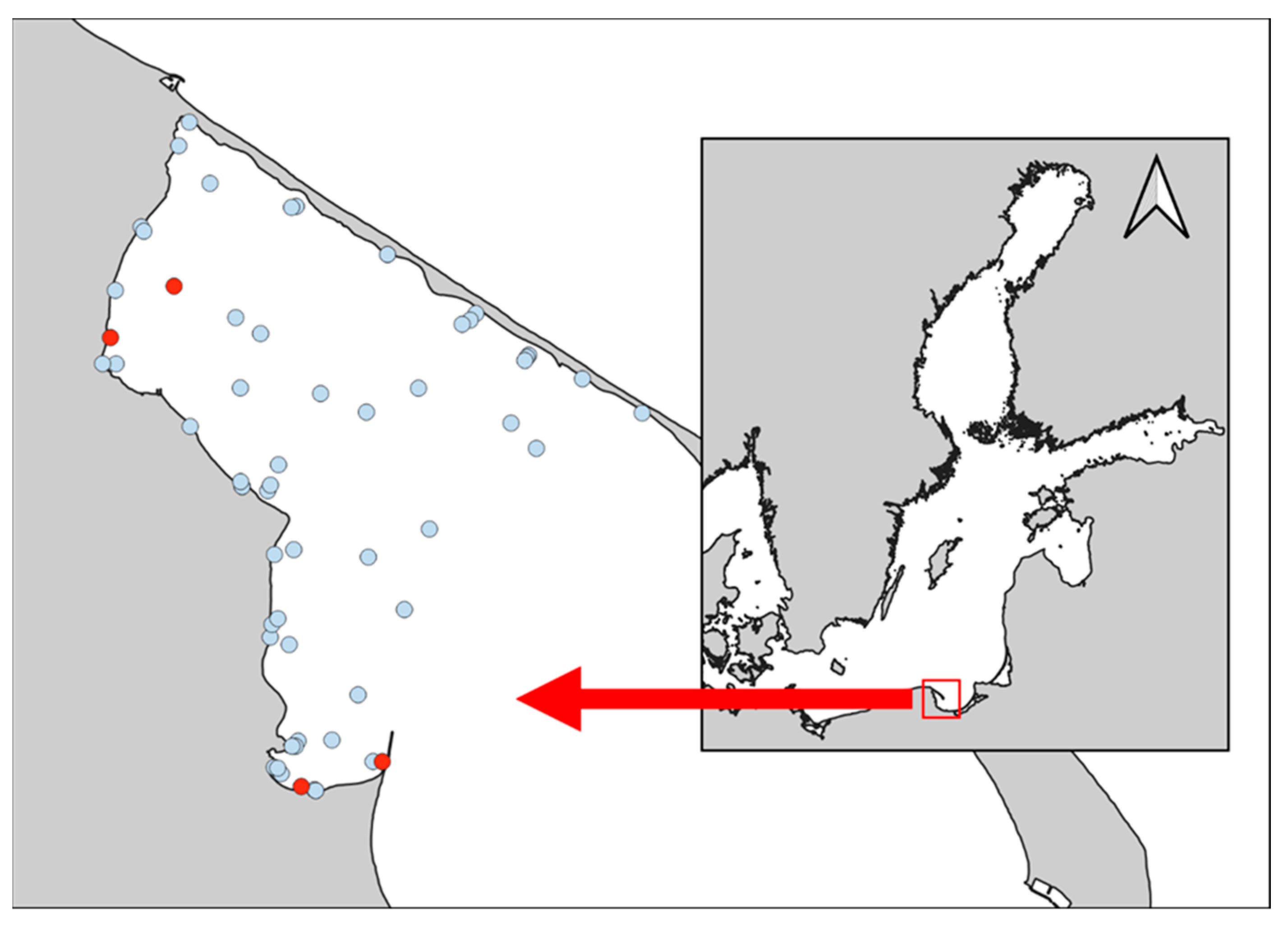
:1. Introduction
2. Materials and Methods
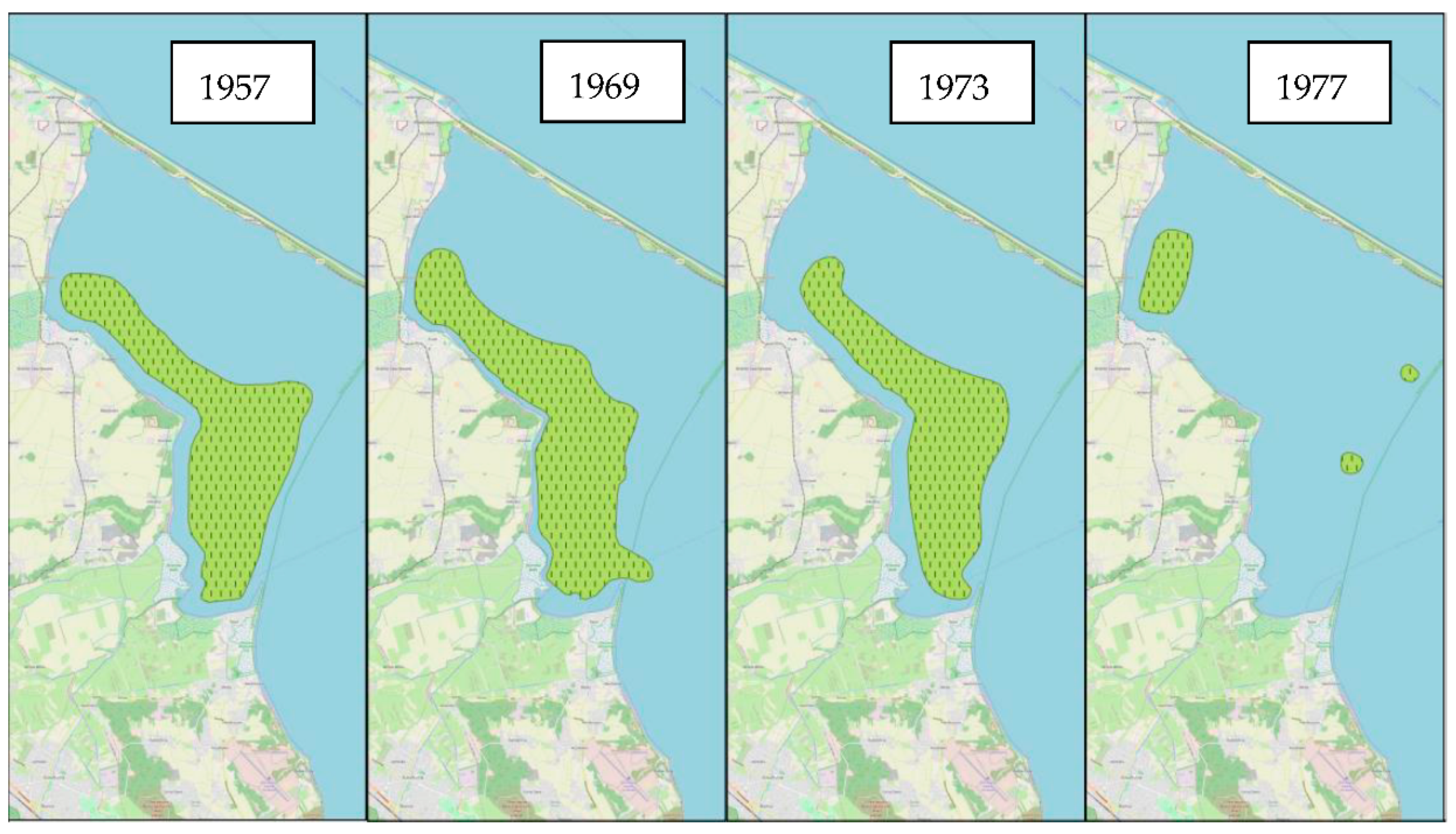
3. Results
4. Discussion
4.1. The State of Populations in the 20th Century
4.2. The State of Populations in the 21st Century
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kentzer, T.; Borowczak, E.; Szczepkowska, E. Investigations on the growth intensity and its modification due to impurities in certain baltic algae. Biol. Morza 3 Studia I Mater. Oceanogr. 1976, 15, 169–186. (In Polish) [Google Scholar]
- Ślesińska, B. The species composition of plants taken when collecting furcellaria from the Puck Bay. Zesz. Nauk. Wydziału Biol. I Nauk. O Ziemii Uniw. Gdańskiego Oceanogr. 1976, 3, 139–148. (In Polish) [Google Scholar]
- Klekot, L. Quantitative analysis of the underwater meadows of the Puck Bay. Oceanologia 1980, 12, 125–140. (In Polish) [Google Scholar]
- Pliński, M. The distribution and biomass of phytobenthos in the internal part of Puck Bay. Studia I Mater. Oceanol. 1982, 39, 195–217. [Google Scholar]
- Kruk-Dowgiałło, L. Long-term changes in the structure of underwater meadows of the Puck Lagoon. Acta Ichthyologica. Et Piscatoria. Suppl. 1991, 21, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Gic-Grusza, G.; Kryla-Straszewska, L.; Urbański, J.; Warzocha, J.; Węsławski, J.M. Atlas of Polish Marine Area Bottom Habitats: Environmental Valorization of Marine Habitats; Węsławski, J.M., Ed.; Broker-Innowacji: Gdynia, Poland, 2009. [Google Scholar]
- Pastuszak, M.; Stålnacke, P.; Pawlikowski, K.; Witek, Z. Response of Polish rivers (Vistula, Oder) to reduced pressure from point sources and agriculture during the transition period (1988–2008). J. Mar. Syst. 2012, 94, 157–173. [Google Scholar] [CrossRef]
- Savchuk, O.P. Large-scale nutrient dynamics in the Baltic Sea, 1970–2016. Front. Mar. Sci. 2018, 5, 95. [Google Scholar] [CrossRef]
- Zalewska, T.; Kraśniewski, W. Assessment of the State of the Environment of the Polish Maritime Areas of the Baltic on the Basis of Monitoring Data from 2018 against the Background of the Decade 2008–2017; Inspekcja Ochrony Środowiska—Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2019. (In Polish) [Google Scholar]
- Grawiński, E.; Kozińska, A.; Paździor, E. Studies on the pathology of South Baltic Sea fish—A review. Życie Weter. 2013, 88, 851–860. [Google Scholar]
- Lang, T.; Kotwicki, L.; Czub, M.; Grzelak, K.; Weirup, L.; Straumer, K. The health status of fish and benthos communities in chemical munitions dumpsites in the baltic sea. In Towards the Monitoring of Dumped Munitions Threat (MODUM); Beldowski, J., Been, R., Turmus, E.K., Eds.; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- Kruk-Dowgiałło, L.; Brzeska, P.; Opioła, R.; Kukliński, M. Macroalgae and Angiosperms. Przewodniki Metodyczne do Badań Terenowych i Analiz Laboratoryjnych Elementów Biologicznych Wód Przejściowych i Przybrzeżnych; Kruk-Dowigiałło, L., Ed.; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2010; ISBN 978-83-61227-36-6. (In Polish) [Google Scholar]
- Pliński, M.; Hindak, F. Chlorophyta (green algae). Filamentous green algae (Ulotrichophycae, Conjugatophycae and Charophyceae). In Flora Zatoki Gdańskiej i Wód Przyległych (Bałtyk Południowy); Pliński, M., Ed.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2012; Volume 7/2. (In Polish) [Google Scholar]
- Pliński, M.; Surosz, W. Red algae and brown algae. In Flora Zatoki Gdańskiej i wód Przyległych (Bałtyk Południowy); Pliński, M., Ed.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2013; Volume 6. (In Polish) [Google Scholar]
- Pliński, M.; Szmeja, J. Seed Plants—Spermatophyta (Magnoliopsida & Liliopsida). In Flora Zatoki Gdańskiej i Wód Przyległych (Bałtyk Południowy); Pliński, M., Ed.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2013; Volume 8. (In Polish) [Google Scholar]
- Braune, W.; Guiry, M.D. Seaweeds. A Colour Guide to Common Benthic Green, Brown and Red Algae of the World’s Oceans; A.r.g. Gantner Verlag: Ruggell, Liechtenstein, 2011; ISBN 9780995567337. [Google Scholar]
- Bunker, F.; Brodie, J.A.; Maggs, C.A.; Bunker, A.R. Seaweeds of Britain and Ireland, 2nd ed.; Marine Conservation Society: Edinburgh, UK, 2017; ISBN 9780995567337. [Google Scholar]
- Lundsteen, S.; Nielsen, R. Coccotylus brodiei, C. truncatus and other Phyllophoraceae (Rhodophyta) in Danish waters. Forum om Marin Bundfauna og –Flora 2015, 2–120. Available online: http://bios.au.dk/fileadmin/bioscience/Forskning/Roskilde/2015-12-08_forum_bundfauna_Coccotylus.pdf (accessed on 25 September 2021).
- Guiry, M.D.; Guiry, G.M. AlgaeBase; National University of Ireland: Galway, Ireland, 2021; Available online: https://www.algaebase.org (accessed on 3 January 2021).
- le Gall, L.; Saunders, G.W. Dna Barcoding Is a Powerful Tool to Uncover Algal Diversity: A Case Study of the Phyllophoraceae (Gigartinales, Rhodophyta) in the Canadian Flora. J. Phycol. 2010, 46, 374–389. [Google Scholar] [CrossRef]
- Rosenvinge, L.K. The Marine Algae of Denmark. Contributions to Their Natural History. Part IV. Rhodophyceae IV. (Gigatinales, Rhodymeniales, Nemastomatales). Det K. Dan. Vidensk. Selsk. Skr. 7 Række 1931, 7, 491–626. [Google Scholar]
- Kylin, H. Die Rhodophyceen Der Schwedischen Westküste. Acta Univ. Lund. N.F. 1944, 40, 1–104. [Google Scholar]
- Zinova, A.D. Etermination Book of the Red Algae of the Northern Seas of the U.S.S.R.; Izdatelstvo Akademii Nauk SSSR: Leningrad, Moscow, 1955. [Google Scholar]
- Newroth, P.R.; Taylor, A.R.A. The nomenclature of the north atlantic species of Phyllophora Greville. Phycologia 1971, 10, 93–97. [Google Scholar] [CrossRef]
- Ciszewski, P.; Demel, K.; Ringer, Z.; Szatybelko, M. Zasoby widlika w Zatoce Puckiej oszacowane metoda nurkowania. Prace Mor. Inst. Ryb. Gdyni II Czesc A, Oceanogr.-Ichtiol. 1962, II, 9–36. [Google Scholar]
- Reprywatyzacja. Available online: http://www.reprywatyzacja.info.pl/b_wsk_fin_od_1939.htm (accessed on 18 December 2021).
- Trokowicz, D.; Andrulewicz, E. Wydobycie i przetwórstwo wodorostów z Zatoki Puckiej. Wiad. Ryb. Pismo Mor. Inst. Ryb. Państw. Inst. Badaw. 2019, 11–12, 20–22. [Google Scholar]
- Weinberger, F.; Paalme, T.; Wikström, S.A. Seaweed resources of the Baltic Sea, Kattegat and German and Danish North Sea coasts. Bot. Mar. 2020, 63, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Kireeva, M.S. Commercial resources of algae in the coastal seas of Soviet Union. Okeanologiya 1965, 5, 14–21. (In Russian) [Google Scholar]
- Trei, T. The physiognomy and structure of the sublittoralmacrophyte communities in Kassari Bay (an area between the isles of Hiiumaa and Saaremaa). Kiel. Meeresforsch-Ungen Sonderh. 1978, 4, 117–121. [Google Scholar]
- Kruk-Dowgiałło, L.; Szaniawska, A. Gulf of Gdańsk and Puck Bay. In Ecology of Baltic Coastal Waters; Schiewer, U., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 139–165. ISBN 978-3-540-73524-3. [Google Scholar]
- Fonseca, M.S.; Thayer, G.W.; Chester, A.J.; Foltz, C. Impact of scallop harvesting on eelgrass (Zostera marina) meadows: Implications for management. N. Am. J. Fish. Manag. 1984, 4, 286–293. [Google Scholar] [CrossRef]
- Creed, J.C.; Amado, G.M.; Filhô, F. Disturbance and recovery of the macroflora of a seagrass (Halodule Wrightii Ascherson) meadow in the Abrolhos Marine National Park, Brazil: An experimental evaluation of anchor damage. J. Exp. Mar. Biol. Ecol. 1999, 235, 285–306. [Google Scholar] [CrossRef]
- d’Avack, E.A.S.; Tillin, H.; Jackson, E.A.S.; Tyler-Walters, H. Assessing the Sensitivity of Seagrass Bed Biotopes to Pressures Associated with Marine Activities; JNCC: Peterborough, UK, 2014.
- Neckles, H.A.; Short, F.T.; Barker, S.; Kopp, B.S. Disturbance of eelgrass Zostera marina by commercial mussel Mytilus edulis harvesting in Maine: Dragging impacts and habitat recovery. Mar. Ecol. Prog. Ser. 2005, 285, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Bird, N.L.; Chen, L.C.-M.; McLachlan, J. Effects of temperature, light and salinity on growth in culture of Chondrus crispus, Furcellaria lumbricalis, Gracilaria tikvahiae (Gigartinales, Rhodophyta), and Fucus serratus (Fucales, Phaeophyta). Bot. Mar. 1979, 22, 521–528. [Google Scholar] [CrossRef]
- Rayment, W.J. Furcellaria lumbricalis a red seaweed. In Marine Life Information Network: Biology and Sensitivity Key Information Reviews; Tyler-Walters, H., Hiscock, K., Eds.; Marine Biological Association of the United Kingdom: Plymouth, UK, 2008; Available online: https://www.marlin.ac.uk/species/detail/1616 (accessed on 13 June 2020).
- Martin, G.; Paalme, T.; Torn, K. Growth and production rates of loose-lying and attached forms of the red algae Furcellaria lumbricalis and Coccotylus truncatus in Kassari Bay, the West Estonian Archipelago Sea. Hydrobiologia 2006, 554, 107–115. [Google Scholar] [CrossRef]
- Bird, C.J.; Saunders, G.W.; McLachlan, J. Biology of Furcellaria lumbricalis (Hudson) Lamouroux (Rhodophyta: Gigartinales), a commercial carrageenophyte. J. Appl. Phycol. 1991, 3, 61–82. [Google Scholar] [CrossRef]
- Kersen, P.; Orav-Kotta, H.; Kotta, J.; Kukk, H. Effect of abiotic environment on the distribution of the attached and drifting red algae Furcellaria Lumbricalis in the Estonian Coastal Sea. Estonian J. Ecol. 2009, 58, 245. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.; Snoeijs, P. Patterns of macroalgal diversity, community composition and long-term changes along the Swedish west coast. Hydrobiologia 2004, 459, 83–102. [Google Scholar] [CrossRef]
- Schories, D.; Selig, U.; Schubert, H. Species and synonym list of the german marine macroalgae based on historical and recent records (Arten- und Synomliste der Makroalgen in den Deutschen Küstengewässern—Auswertung von historischen und rezenten Befunden. Rostock. Meeresbiol. Beitr. 2009, 21, 7–135. [Google Scholar]
- Osowiecki, A.; Łysiak-Pastuszak, E.; Kruk-Dowgiałło, L.; Błeńska, M.; Brzeska, P.; Kraśniewski, W.; Lewandowski, Ł.; Krzymiński, W. Development of tools for ecological quality assessment in the Polish marine areas according to the Water Framework Directive. Part IV—Preliminary assessment. Oceanol. Hydrobiol. Stud. 2012, 41, 1–10. [Google Scholar] [CrossRef]
- Kornaś, J.; Medwecka-Kornaś, A. Associations Végétales Sous-Marines Dans Le Golfe de Gdańsk (Baltique Polonaise). Vegetatio 1950, 2, 120–127. [Google Scholar] [CrossRef]
- Sparre, A. The Climate of Denmark, Summaries of Observations from Light Vessels (IV), Salinity, Means, Extremes and Frequency; Danish Meteorological Institute: Copenhagen, Denmark, 1984; Volume 11. [Google Scholar]
- Jodziewicz, B. Citizen Science as a Tool for the Study of Rare Algae Species (Nauka Obywatelska Jako Narzędzie do Studiowania Rzadkich Gatunków Glonów). Bachelor’s Thesis, University of Gdańsk, Gdańsk, Poland, 2021. [Google Scholar]
- Brzeska-Roszczyk, P.; Kruk-Dowgiałło, L. Preliminary results of monitoring studies on macrophytes in the area of brine discharge from the creation of gas storage caverns (Puck Bay, Baltic). Biul. Inst. Mor. 2017, 32, 50–65. [Google Scholar] [CrossRef]
- Bełdowska, M.; Jędruch, A.; Zgrundo, A.; Ziółkowska, M.; Graca, B.; Gębka, K. The influence of cold season warming on the mercury pool in coastal benthic organisms. Estuar. Coast. Shelf Sci. 2016, 171, 99–105. [Google Scholar] [CrossRef]
- Sokołowski, A.; Ziółkowska, M.; Zgrundo, A. Habitat-related patterns of soft-bottom macrofaunal assemblages in a brackish, low-diversity system (Southern Baltic Sea). J. Sea Res. 2015, 103, 93–102. [Google Scholar] [CrossRef]
- Zgrundo, A. EN Ruppia maritima, L. Rupia morska. In Czerwona Księga Roślin Naczyniowych Pomorza Gdańskiego TOM 1 Zagrożone Gatunki Nadmorskich Plaż, Wydm i Solnisk oraz wód Słonawych Strefy Przymorskiej; Lazarus, M., Afranowicz-Cieślak, R., Eds.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2020; pp. 180–183. ISBN 978-83-8206-047-8. (In Polish) [Google Scholar]
- Zgrundo, A.; Złoch, I.; Kobos, J. Macroflora of the Puck Bay. In The Puck Bay Vol. 3; Bolałek, J., Burska, D., Eds.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland. (In Polish)
- Zgrundo, A.; Lazarus, M. VU Zannichellia palustris L. Zamętnica błotna. In Czerwona Księga Roślin Naczyniowych Pomorza Gdańskiego TOM 1 Zagrożone Gatunki Nadmorskich Plaż, Wydm i Solnisk oraz wód Słonawych Strefy Przymorskiej; Lazarus, M., Afranowicz-Cieślak, R., Eds.; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2020; pp. 183–187. ISBN 978-83-8206-047-8. (In Polish) [Google Scholar]
- Sousa, W.P. Intertidal mosaics: Patch size, propagule availability, and spatially variable patterns of succession. Ecology 1984, 65, 1918–1935. [Google Scholar] [CrossRef]
- Díez, I.; Santolaria, A.; Secilla, A.; Gorostiaga, J.M. Recovery stages over long-term monitoring of the intertidal vegetation in the ‘abra de bilbao’ area and on the adjacent coast (N. Spain). Eur. J. Phycol. 2009, 44, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Ko, Y.W.; Yang, K.M.; Sung, G.; Kim, J.H. Effects of disturbance timing on community recovery in an intertidal habitat of a Korean rocky shore. Algae 2017, 32, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Sztobryn, M.; Wójcik, R.; Mietus, M. Występowanie zlodzenia na Bałtyku—Stan obecny i spodziewane zmiany w przyszłości. In Warunki Klimatyczne i Oceanograficzne w Polsce i na Bałtyku Południowym. Spodziewane Zmiany i Wytyczne do Opracowania Strategii Adaptacyjnych w Gospodarce Krajowej, Seria Monografie; Wibig, J., Jakusik, E., Eds.; IMGW-PIB: Warszawa, Poland, 2012; pp. 189–215. [Google Scholar]
- Panetta, A.M.; Stanton, M.L.; Harte, J. Climate warming drives local extinction: Evidence from observation and experimentation. Sci. Adv. 2018, 4, eaaq1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, G.; Harley, C.; Faria, J.; Vale, M.; Hawkins, S.; Neto, A.; Arenas, F. Direct and indirect effects of climate change squeeze the local distribution of a habitat-forming seaweed. Mar. Ecol. Prog. 2019, 626, 43–52. [Google Scholar] [CrossRef]
- Coelho, S.M.; Rijstenbil, J.W.; Brown, M.T. Impacts of anthropogenic stresses on the early development stages of seaweeds. J. Aquat. Ecosyst. Stress Recover. 2000, 7, 317–333. [Google Scholar] [CrossRef]
- Schiel, D.R.; Lilley, S.A. Impacts and negative feedbacks in community recovery over eight years following removal of habitat-forming macroalgae. J. Exp. Mar. Biol. Ecol. 2011, 407, 108–115. [Google Scholar] [CrossRef]
- Graiff, A.; Bartsch, I.; Ruth, W.; Wahl, M.; Karsten, U. Season exerts differential effects of ocean acidification and warming on growth and carbon metabolism of the seaweed Fucus vesiculosus in the Western Baltic Sea. Front. Mar. Sci. 2015, 2, 112. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.J.; Kim, K.Y. Effects of future climate conditions on photosynthesis and biochemical component of Ulva pertusa (Chlorophyta). Algae 2016, 31, 49–59. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zgrundo, A.; Złoch, I. Gone and Back—The Anthropogenic History of Coccotylus brodiei (Turner) Kützing and Furcellaria lumbricalis (Hudson) J.V. Lamouroux in the Gulf of Gdańsk (Southern Baltic Sea). Water 2022, 14, 2181. https://doi.org/10.3390/w14142181
Zgrundo A, Złoch I. Gone and Back—The Anthropogenic History of Coccotylus brodiei (Turner) Kützing and Furcellaria lumbricalis (Hudson) J.V. Lamouroux in the Gulf of Gdańsk (Southern Baltic Sea). Water. 2022; 14(14):2181. https://doi.org/10.3390/w14142181
Chicago/Turabian StyleZgrundo, Aleksandra, and Ilona Złoch. 2022. "Gone and Back—The Anthropogenic History of Coccotylus brodiei (Turner) Kützing and Furcellaria lumbricalis (Hudson) J.V. Lamouroux in the Gulf of Gdańsk (Southern Baltic Sea)" Water 14, no. 14: 2181. https://doi.org/10.3390/w14142181