Abstract
The European lobster, Homarus gammarus, was sampled from September 2016 to August 2017, using pots and gillnets in the Eastern Adriatic Sea. Official landings were also analyzed (2008–2020). The majority of landings were from the Northern Adriatic and showed an increase of 18.5% over the study period. Results revealed an extremely low catch rate that fluctuated significantly by season. Dominance by specimens in the size classes of 90–110 mm carapace length (CL) was observed. Experimental design revealed spatial heterogeneity, with larger individuals caught further from the coast (>3 nm) and undersized specimens caught near the coast (<3 nm). With increasing CL and total length (TL), an increase in weight was higher in males than females. TL and abdomen width (AW) increased linearly with CL and were more pronounced in females. At 80 mm CL, females started to develop secondary sex characteristics with wider AW than males. The results suggest that the modern legal framework is appropriate for lobster fisheries and a longer fishing season is not advisable, despite a positive catch trend in the northern part of the sea. Additional efforts are desirable to reduce fishing pressure in the coastal part. The morphometric relationships could reveal population characteristics required as inputs in stock assessment analysis for effective management.
1. Introduction
Modern fisheries management design and implementation should draw on insights into biological, environmental, and socioeconomic issues, and their interconnections at the local, regional, or national scales [1]. Fisheries management tools are imperative for the Mediterranean Sea, due to its heterogeneity and uncertainty surrounding the implementation of measures to date [2,3,4]. It should be stressed that the coastal Mediterranean fisheries, whether artisanal, recreational, subsistence or a combination of the above, play an important socioeconomic role in Europe [5,6].
Small-scale fishers (SSF) along the Mediterranean coast operate with a variety of fishing gears on a seasonal basis, and catches are composed of a wide range of species [7]. Although the European lobster, Homarus gammarus, is not a target species of SSF, it is often a bycatch in gillnets targeting different fish and other marine organisms [8,9,10]. Both H. gammarus and spiny lobster (Palinurus elephas) represent a valuable income in SSF during the warmer months, which coincides with their fishing seasons. Since it is difficult to evaluate SSF landings and they are often underestimated, official landings statistics both in the Mediterranean and Adriatic Seas should be considered with caution. Lotze et al. [11] reconstructed official commercial invertebrate landings in the Adriatic Sea by the Italian fleet from 1970 to 2006, and simulated that landings of H. gammarus were around 48 t per year in the 1990s, but declined to below 20 t per year in the 2010s, suggesting the depletion of stocks in the Adriatic Sea. The total landings of the European lobster throughout its distribution range fluctuated from 4181 to 5607 tons per year [12], in the period from 2007 to 2016. Around 80% of landings came from the North-Eastern Atlantic, while landings in Mediterranean countries never reached those levels [13]. New estimations confirmed the general underestimates of the FAO data in the French Mediterranean [14], suggesting that they might be generally accurate for the whole distribution. It was clearly demonstrated that specific lobster stocks in the Mediterranean suffered from overexploitation [15,16,17], with sharp declines in landings reported for more than a century [18,19]. The European lobster is targeted mainly by trap fisheries in Northern Europe [19,20] and with baited pots and different gillnets throughout the coastal Mediterranean Sea [14]. In the Mediterranean, the EU rules define a minimum landing size of 105 mm carapace length or 300 mm total length [21].
European lobster has a broad geographic distribution, from Northern Norway to the Mediterranean Sea [22]. This species inhabits the whole Adriatic Sea, though population abundance is higher in the northern waters [23]. Studies on its biology and ecology in the Adriatic Sea as well as throughout the Mediterranean, are severely lacking in comparison with Northwestern European waters [22,24,25,26,27,28,29]. Genetic analysis suggests a division between the stocks in the Atlantic and the Mediterranean Sea [30,31], and also between Mediterranean populations [32]. Recently, Pavičić et al. [23] confirmed a genetic division among different Mediterranean basins, with the panmictic characteristics of Adriatic populations. They also showed that anthropogenic impacts did not significantly affect H. gammarus genetic diversity. Exploitation patterns and biological data of H. gammarus around Corsica were reported by Pere et al. [14]. In the absence of published data, parameters for complete population assessment of any crustacean species (i.e., sex, growth, age, and mortality), morphometric relationships, and allometry are useful tools to reveal population characteristics during growth [33,34], to estimate population biomass, to compare morphological traits between different species or populations across the distribution range [35,36,37], and to estimate functional maturity [27,38]. Further, the ratio of abdominal width to carapace length (AW/CL) can be potentially used in female lobsters to establish size at maturity [39]. This is important since female morphology might respond more strongly to local selection pressures than male morphology [24].
The primary aim of this study was to gather additional biological data and assess the population characteristics of H. gammarus in the Eastern Adriatic Sea, since to date there is no complete analysis of the morphometric relationships in the area, as well as in its entire distribution range. In that sense, data regarding length frequency distribution, catch per unit effort, morphometric relationships, and allometry, together with the W/CL index (condition factor), were analyzed.
2. Materials and Methods
2.1. Study Site
For management purposes and data collection, the Croatian marine fishing area was administratively divided into smaller units (fishing zones—open sea A, B, C, D, internal E, F, G, and Ecological and Fisheries Protection Zone H, I, J, as a protected fishing area within Croatia’s Exclusive Economic Zone). Official landing statistics for H. gammarus, obtained from the Croatian Fisheries Directorate (for the years 2008–2020) by fishing zone and fishing gear, were used for the recent trend evaluation. The majority of official landings occurs in the Northern Adriatic (fishing zones A and E).
Sampling of Homarus gammarus was conducted over a one-year period from September 2016 to August 2017. Along the Eastern Adriatic coast, sampling was conducted in four larger areas, mainly (92.4% of the total sample) off the west coast of Istria (fishing zone A up to 35 m of depth; sandy-muddy bottom), while the remaining samples were collected off the coast of the Mali Lošinj Island (fishing zone E up to 50 m of depth, rocky–sandy–muddy bottom), the Hvar Island (fishing zone G up to 70 m of depth; hard bottom), and the Vis Island (fishing zone C up to 90 m of depth; hard bottom) (Figure 1). For the purpose of spatial analysis, fishing zone A was divided into three subzones based on distance from the coast, as follows—zone 1 (3 nautical miles, nm from the coast), zone 2 (from 3 to 8 nm), and zone 3 (from 8 to 12 nm).
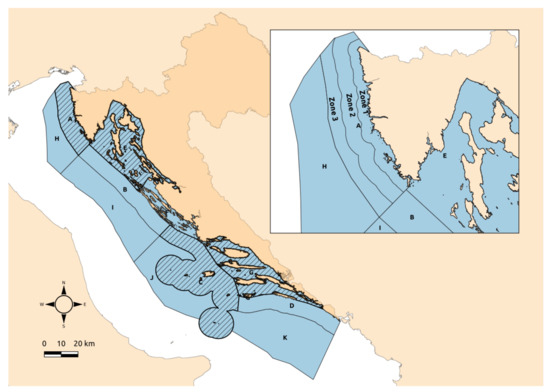
Figure 1.
Sampling area of the European lobster, Homarus gammarus, in the Eastern Adriatic Sea (fishing zones A, E, C, and G); fishing zone A is divided into subzones according to distance from the coast (zones 1, 2, and 3).
Generally, the shallow Northern Adriatic is a unique marine ecosystem characterized by specific hydrographic conditions and a relatively high biodiversity. This is the most productive part of the Adriatic Sea due to the influence of the River Po [40,41], with strongly pronounced multi-year and annual fluctuations in environmental parameters (temperature, salinity, and nutrients) [42,43].
2.2. Data Collection
A total of 1273 individuals of the H. gammarus, sampled in the Northern and Central Adriatic Sea from September 2016 to August 2017, were used for the morphometric analysis. Of this total, 1176 individuals were sampled experimentally using baited pots of 40 mm square mesh side, in cooperation with local fishers in fishing zone A, to determine the share of undersized specimens in relation to the distance from the coast. The pots were constructed using a nylon square gillnet affixed to a frame with a single entrance made of wire. The pots were usually placed on the sea bottom in groups of 12 pots per 5 positions (25–35 m depth). Fish remains were used as bait. The remaining 97 individuals, used only for morphometric analysis, were sampled using gillnets (60 mm square mesh side), in cooperation with local small-scale fishers in fishing zones C, E, and G, at a depth range of 40–80 m.
Catch per unit effort (CPUE) was recorded as the weight of lobsters per pot. All lobsters were measured on board or on the coast. Biometric measurements (carapace length (CL), total length (TL), and abdomen width (AW)) were measured using Vernier calipers (to the nearest 0.1 cm), specimens were weighed (W) (recorded to 0.1 g), and sex was determined macroscopically (male/female). CL was measured from the posterior margin of the eye socket to the posterior end of the carapace. TL was measured from tip of the rostrum to the end of the abdomen. AW was measured at the widest point of the second abdominal segment [24,38]. W/CL index or condition index [34] was estimated from individuals used in the morphometric analysis, and was calculated as total weight (W) divided by carapace length (CL).
2.3. Data Analysis
2.3.1. Morphometric Relationships and W/CL Index
To examine the morphometric relationships and allometry between male and female European lobster, only individuals for which sex could be determined were included in the analysis (N = 936). Morphometric relationships are used to predict the value of Y from X, while allometry is used to test how one variable corresponds to another [44]. The following morphometric relationships were examined for males and females—W vs. CL, TL vs. CL, W vs. TL, and AW vs. CL. The relationships W vs. CL and W vs. TL are described with the power function y = axb [45], while TL vs. CL and AW vs. CL are described by a linear function. Morphometric relationships were examined by ordinary least-squares regression [44], and Student’s t-test regression slopes were used for comparisons by sex [46]. Additionally, reduced major axis regression (standardized major axis regression) on log-transformed data was used to estimate the slopes and to test for allometry [44]. Student’s t-test was used to test for allometry (b = 1 for length–length relationship, b = 3 for length–weight relationship) [47].
2.3.2. Multivariate Analysis
To assess for seasonal variation of CPUE and the W/CL index, the sampling period was analyzed by season; autumn (October–December), winter (January–March), spring (April–June), and summer (July–September). Permutational analysis of variance (PERMANOVA, [48]) was used to test for differences between seasons (fixed, 4 levels—autumn, winter, spring, and summer) of CPUE data and between fishing zones (fixed, 3 levels—1, 2, and 3) of CL, TL, and W data. Where appropriate, post-hoc pair-wise comparisons were used for further testing. PERMANOVA was also used to test for differences between season and sex (fixed, 2 levels—female and male) of the W/CL index. Since the W/CL index is influenced by size [34,49], CL was used as a covariate for further testing of seasonal variation in the W/CL index. PERMANOVA was performed in the PRIMER-E software [50] with the add-on package PERMANOVA+ [51].
Ordinary least-squares regression and reduced major axis regression analyses and graphs were performed in SigmaPlot v.14.0 (Systat Software, Inc., San Jose, CA, USA, www.systatsoftware.com accessed on 9 April 2021) and PAST v.4.05 [52]. R software was used for producing graphs [53].
3. Results
According to the Croatian Fisheries Directorate, the average annual landings of H. gammarus in the last 13 years were around 3308.12 kg (from 1989.95 kg in 2008 to 4723.88 kg in 2016). Landings fluctuated around the average in the last couple years, while an increasing trend of 18.5% was observed for the entire period. H. gammarus is mostly captured in fishing zones A and E, covering the coastal zone in the Northern Adriatic, with a continuous increase in catches (Fisheries Directorate 2020) (Figure 2A,B). Landed quantities from other fishing zones were negligible and low (fishing zones H, I and J) or non-existent (fishing zone K).
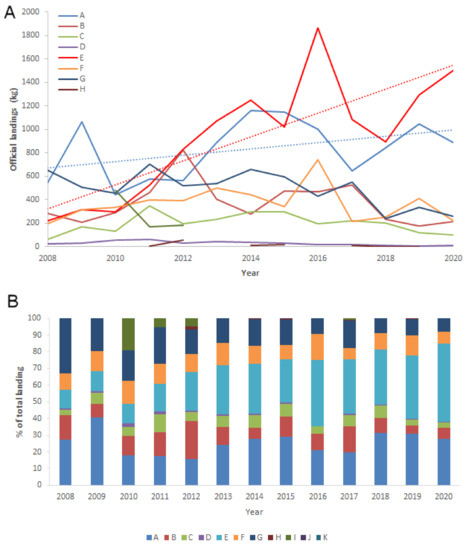
Figure 2.
(A) Official landings of the European lobster, Homarus gammarus (Croatian Fisheries Directorate 2008–2020) with the indicated trends in fishing zones A and E. (B) Percentage of landed catch in each fishing zone.
Lobsters below the minimum landing size (MLS) of 105 mm CL accounted for 52.83% of the total sample (Figure 3) and were returned to the sea. Lobsters below MLS in autumn, winter, spring, and summer accounted for 59.8%, 50.4%, 61%, and 52.4% of the catch, respectively. Sex ratio was slightly in favor of males (males 52.7%, females 47.3%). CL size of females ranged from 63 to 180 mm (104.01 ± 20.96 mm, mean ± standard deviation) and that of males ranged from 55 to 189 mm (103.78 ± 21.05 mm). Weight of females and males ranged from 180 to 2480 g (mean 735.72 ± 454.62 g) and 105 to 4118 g (725.55 ± 491.47 g), respectively. CPUE fluctuated from 4.6 to 239 g/pot. The mean value of CPUE was lowest in autumn (43.7 g/pot), while CPUEs were similar in spring (102 g/pot) and summer (101.2 g/pot) (Figure 4). One-way PERMANOVA showed that CPUE values were significantly different between seasons (Pseudo-F = 12.16, p = 0.001). All pair-wise comparisons between seasons were significantly different, except between spring and summer (p > 0.05).
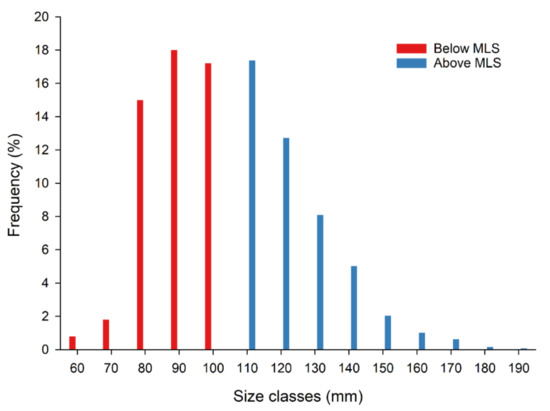
Figure 3.
Carapace length (CL) size distribution of the total sample of the European lobster (Homarus gammarus) in relation to the minimum landing size (MLS = 105 mm CL).
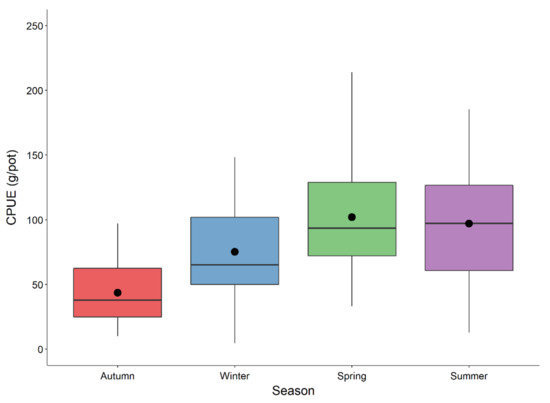
Figure 4.
Seasonal variation of catch per unit effort CPUE (g/pot) of the European lobster (Homarus gammarus) in the Adriatic Sea.
Generally, females and males gradually increased in CL, TL, and W from the coast to open waters. Female CL in fishing zones 1, 2, and 3 were 100.05 ± 19.54 mm, 111.92 ± 16.82 mm, and 117.21 ± 21.52 mm, while male CL in the same zones were 98.84 ± 17.52 mm, 115.79 ± 20.68 mm, and 113.67 ± 22.71 mm, respectively. Female TL in fishing zones 1, 2, and 3 was 294.69 ± 52.85 mm, 325.27 ± 47.0 mm, and 343.82 ± 63.31 mm, while male TL was 286.25 ± 45.86 mm, 325.76 ± 49.41 mm, and 334.12 ± 61.70 mm, respectively. Female weight in fishing zones 1, 2, and 3 was 633.99 ± 387.84 g, 860.38 ± 397.85 g, and 1222.56 ± 588.99 g, while male weight was 595.28 ± 337.54 g, 941.18 ± 518.43 g, and 1122 ± 611.87 g, respectively (Figure 5).
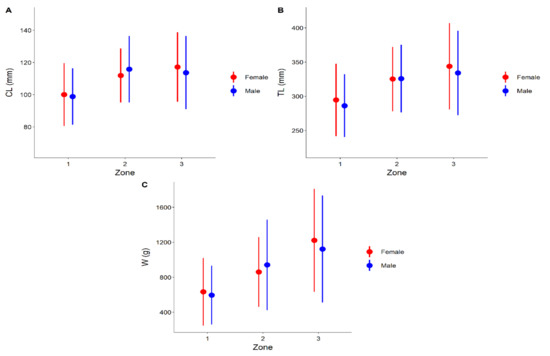
Figure 5.
Variation in CL (A), TL (B), and W (C) (mean ± standard deviation) of female and male European lobster by fishing zone. CL, carapace length; W, weight; and TL, total length.
For all three measurements (CL, TL, and W), a two-way PERMANOVA showed a significant difference between fishing zones (for CL Pseudo-F = 37.86, p = 0.001; for TL Pseudo-F = 42.09, p = 0.001; for W Pseudo-F = 78.72, p = 0.001), though these differences were not significant by sex or in the zone–sex interaction. Pair-wise comparisons revealed significant differences between fishing zones 1 and 2 and fishing zones 1 and 3, but not between fishing zones 2 and 3.
Morphometric relationships of ordinary least-squares regressions of males and females are presented in Table 1 and Figure 6. The relationships CL vs. W and TL vs. W are described with a power function (for all relationships R2 > 0.89). By increasing both CL and TL, W increased more in males than in females (Figure 6A,B). Other dimensions (TL and AW) increased linearly with CL (Figure 6C,D). With increasing CL and TL, AW increased more in females than males. At 80 mm CL, females started to develop secondary sexual characters, and a wider AW than in males. The slopes of all ordinary least-squares regressions were significantly different between females and males (Table 1). Results of the reduced major axis regressions for all relationships with log-transformed data between females and males are presented in Table 2. The relationships log CL vs. log W and log CL vs. log TL for both sexes showed negative allometry, while the log TL vs. log W relationships for both sexes showed positive allometry. All relationships between sexes exhibited the same type of allometry, except for log AW vs. log CL relationships, which showed positive allometry for females and negative allometry for males.

Table 1.
Parameters of ordinary least-squares regressions between males and females of the European lobster (Homarus gammarus).
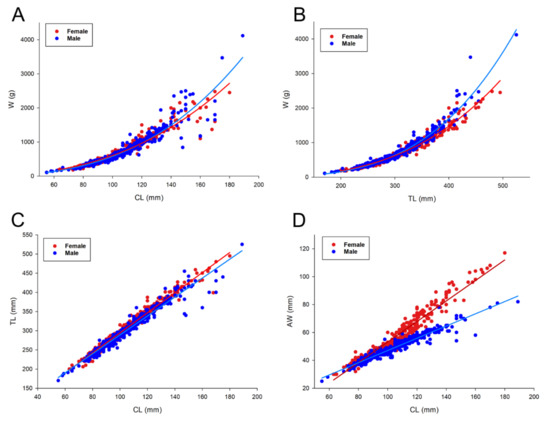
Figure 6.
Morphometric relationships of male and female European lobster (Homarus gammarus) from the Eastern Adriatic Sea—(A) CL vs. W; (B) TL vs. W; (C) CL vs. TL; and (D) CL vs. AW. CL, carapace length; W, weight; TL, total length; and AW, abdomen width.
Table 2.
Parameters of reduced major axis regressions between male and females and allometry tests of the European lobster (Homarus gammarus).
Mean values of the W/CL index were higher in females in all seasons, except spring. Namely, the mean value of the W/CL (g/mm) index of females and males, respectively, was 6.27 and 6.09 in fall; 7.03 and 5.88 in winter; 5.91 and 6.11 in spring; and 7.01 and 6.87 in summer (Figure 7). Results of two-way PERMANOVA on the W/CL index showed a significant effect of season, but with no significant differences for sex or sex–season interaction. After controlling for the significant effect of CL (covariate), the effect of season on the W/CL index did not remain significant.
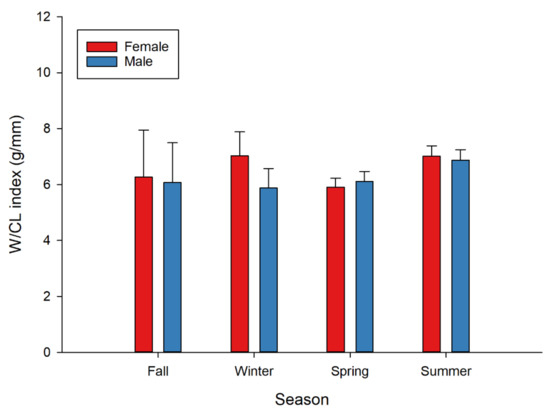
Figure 7.
Seasonal variation in the W/CL index (condition factor) of female and male European lobster (Homarus gammarus). Error bars represent 95% confidence intervals.
4. Discussion
All Homarus gammarus stocks along European coasts are progressively declining as a consequence of intensive fisheries, high fishing effort and high efficiency of fishing gears (specifically the improved design of pots) [54]. However, European lobster fisheries are generally unregulated, or minimally regulated only for a minimum size or for landing of berried females [19]. As fishery management moves globally from a single-species to an ecosystem-based emphasis, it is important to understand the function of this species in the marine ecosystem. Unfortunately, in Mediterranean waters, there is a lack of biological and ecological data on H. gammarus as an exploited species, particularly there is a lack of knowledge on how the abundance and consequently landings change throughout its distribution area, in the light of high anthropogenic pressure, particularly of climate change. It is well-known that environmental factors contribute to the interannual variability of key parameters for all marine stocks, and changes in these factors might affect their population dynamics [55]. Moreover, the reliability of landings reported by all Mediterranean countries, including those in the EU are marked by high uncertainty, and there are also reports of general underestimates of FAO data in the Mediterranean [14]. Croatian reported landings of the European lobster, ranging from 1.9 to 4.7 tons per year, showed an increase in recent years; however, these data should be considered with caution since they are based solely on fisher logbooks. These data are often far from reality, since underestimating the total catch is widespread, due to misreporting out-of-season catches and not quantifying discards regarding lobsters caught below the minimum size [2,56,57]; while IUU landings are substantial [58]. All this highlights the lack of relevant data to determine the state of H. gammarus stocks in the Mediterranean Sea, and the high uncertainty concerning the adoption of measures for its effective management.
In addition to overexploitation, a pattern of low H. gammarus abundance is evident throughout the Mediterranean, where environmental conditions are less favorable for this species, in comparison to the Atlantic [14]. This is becoming increasingly significant due to climate change [59,60]. Excessively high sea temperatures might lead to reduced population abundance [30], particularly affecting juvenile survival and recruitment during summer. Such a situation could encourage migration from a warmer to a relatively colder area that might offer more favorable conditions for the reproduction and development of juveniles. The Northern Adriatic could be considered to be a potentially particularly advantageous area, which hosts several species that are adapted to boreal climatic conditions—like the H. gammarus—and is configured as a cul-de-sac [61,62]. In this study, CPUE fluctuated from 4.6 to 239 g/pot and differed significantly between seasons, achieving peak values in the warmer months, which correlates with the open fishing season from 5 May to 31 August [63]. Pere et al. [14] reported that the CPUE of H. gammarus varied significantly as a function of month, fishing area, and depth in a Mediterranean small-scale fishery. Our results also indicate extremely low catch rates, which could imply low abundance of this species in the study area, as was previously suggested by Pere et al. [14]. However, fluctuations in H. gammarus abundance might occur as a consequence of a combination of environmental and fishery-related processes, like trap efficiency and target species behavior ([26] and references therein), including feeding behavior and molting status [64]. It is long known that sea temperature affects behavior and availability of traps for homarid lobsters [65]. In the present study, undersized specimens were predominantly caught in spring and autumn (>59%), likely due to the increase in lobster feeding activity prior and after spawning, and the increased sea temperature [26,28]. Sex ratio was slightly in favor of males, which might suggest a consistent female avoidance of traps year round, due to constant food or shelter-seeking behavior [66]. We found that the longer distance from the coast, the larger were the individuals of H. gammarus of both sexes. Length frequency distribution suggested that the largest individuals were in the length class of 190 mm CL, with no individuals smaller than 60 mm CL. Similar observations were reported by Latrouite [67] and Prodöhl et al. [13] in the Atlantic, and Pere et al. [14] in the Corsican Mediterranean Sea, concluding that the cryptic behavior and limited movement of juveniles could explain the absence of small and young European lobsters in catches. However, the length distribution in this study, similar to Pere et al. [14] confirmed their conclusion of shifting towards larger individuals as compared to stocks from the North Atlantic [38,68,69]. These results could also be linked to gear selectivity [14,70], which was evident here, since more than half of all specimens below the MLS were caught in the 40 mm square mesh side sampling pots, as opposed to the 55 mm square mesh side required by law [71].
Official statistics data indicated that most of the catch was off the Northern Adriatic coast. The heterogeneous bottom and relatively shallow depths make this area the most suitable habitat. This is because the rocky substrate provides shelter from predators, while the soft sediment increases food availability [23] and the potential for growth and reproduction for the European lobster [72]. Generally, the eastern coast of the Northern Adriatic is steep, with minimal recent sedimentation and strong karst relief, which influences sediment distribution patterns [73]. It was reported that the most suitable habitat for the European lobster was at the boundary between sedimentary and rocky-bottoms, which coincided with seafloor depressions with a steep slope, with medium to high wave energy conditions, and were located within a range of water depths of 35–40 m [25]. H. gammarus exhibit low migration activity (within 3.8 km) related to the spatial configuration of the local lobster habitat, with a marked tendency for offshore movement [28,74].
Age determination is important in life history studies and effective stock management [75]. Since age determination in crustacean species is complicated [76], given the lack of hard body structures after every molt, morphometric relationships can instead be used for stock management [35,36,37]. Differences in morphometric traits among populations typically result from a combination of genetic variation and different environmental conditions [77], and can be heritable. In this case, populations might diverge due to responses to local selection pressures [78]. In marine species, these pressures are proposed as the predominant mechanism of divergence despite the existence of gene flow barriers [79].
Morphometric relationships and allometry were studied in many crustacean species, including crabs [80,81], spiny lobsters [34,82,83], American lobster [37,84,85], and European lobster [27,33,38]. In the present study, ordinary least-square regressions showed that W increased following a power function with both CL and TL, while TL and AW increased linearly with CL. Comparing the slopes of the ordinary least-square regressions, we observed significant differences of all relationships between males and females, indicating different growth rates by sex [38,86]. The negative allometry, i.e., a decreasing growth rate of TL vs. CL and CL vs. W for both sexes, reflects a morphological change from an elongated body towards a heavier one, as also reported by Martínez-Calderón et al. [34] for the Caribbean spiny lobster (Panulirus argus) and Radhakrishnan et al. [87] for Panulirus homarus. French McCay et al. [84] reported a coefficient b of 2.934 for an American lobster population, which is similar to our results that gave a coefficient b below 3 for both sexes, while Steinback et al. [85] reported a coefficient b of 3.347 for males and 2.972 for females for American lobster. Baggio [37] reported isometric growth of H. americanus (CL vs. W). We found positive allometry for log TL vs. log W for both sexes, while for the log CL vs. log AW relationship, allometry was positive for females and negative for males. Positive allometry was also found between TL and W in male European lobsters in the Northern Adriatic [33]. Variations in the W/CL index did not show a significant effect of season after controlling for CL as a covariate. A seasonal variation of this index was reported by Martínez-Calderón et al. [34], suggesting this was due to a combination of carry-over effects and variation in local factors, such as predation risk and food availability [88]. Further on, the usefulness of the W/CL index for comparing the nutritional status between groups or populations was reported by Martínez-Calderón et al. [34]. Monitoring the levels of morphometric condition might provide insight into the potential factors decoupling the relationships between the levels of successful settlement of juveniles and the abundance of adults.
In the present study, the AW vs. CL relationship showed that a significant difference in AW between the sexes was observed starting at 80 mm CL, as females emerged from molt with a proportionally wider abdomen than males. The AW/CL ratio was used to assess morphometric maturity that preceded functional maturity [27,39]. In this study, no inflection points were observed in the AW/CL ratio. However, using the AW/CL relationship to assess size at maturity should be considered with caution [89]. Geographical variability in size at maturity was reported by several authors. Woodruff [90] reported functional and morphometric size of 91.6 mm and 84.3 mm CL at maturity, respectively, for H. gammarus in Northumberland inshore fishery coastal waters, UK. Tully et al. [27] estimated the value of functional L50 to be higher than 92.5 mm, while Laurans et al. [69] reported an even greater functional maturity L50 value of 103–106 mm CL. Lizárraga-Cubedo et al. [38] found a smaller size at maturity for a population from the Firth of Forth (around 80 mm CL) than those from the Hebrides, Scotland. A number of factors were linked to variations in size at maturity for the Homarus species, including temperature [89,91,92], fishing pressure [93,94], intraspecific competition [95], and predation [92,96]. These differences could also be due to female catchability [54], although Laurans et al. [69] found no differences in the catchability of ovigerous and non-ovigerous females. Size at maturity is an indicator of the suitability of the MLS [38,69]. Currently, the European Union (EU) issued an MLS of 105 mm CL as the only management measure for the Mediterranean Sea, aiming to protect juveniles and ensure that sufficient spawning occurs before individuals are vulnerable to the fishery [92]. The same MLS is established in Croatia. Prohibition of landing berried females and an extended closed season (September to early May) are also management measures implemented in Croatia [63]. Although a selectivity analysis of pots intended for catching fish (40 mm) and large crabs (55 mm) was not conducted, it is likely that the ratio of H. gammarus specimens, under 105 mm CL, would be markedly lower than that obtained here. Our results suggest that the current legal framework in Croatian marine fisheries for H. gammarus keep landings in a positive trend, with biological characteristics that do not indicate a significant change within the analyzed population. However, an extension of the fishing season (to encompass April and September) is not advisable, despite the positive catch trend in recent years. Additional effort is desirable to reduce fishing pressure in the coastal part (<3 nm), not only for active fishing gears [21] but also for passive gears, such as different gillnets and pots.
Author Contributions
All authors conceived the research. S.M.-S., M.P. and D.V. contributed to sample design, sample collection, and analysis. S.M.-S., M.P. and D.V. analyzed data. Additional analysis was provided by A.V., S.M.-S. and M.P. wrote and revised the manuscript. All authors have reviewed and approved the final manuscript.
Funding
This study was partially supported by the Croatian Science Foundation (HRZZ) under project IP-2016-06-9884 (NurseFish).
Data Availability Statement
Data used in this manuscript are available from the corresponding author upon reasonable request.
Acknowledgments
The authors are grateful to local fishers for assistance in collecting samples.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Jennings, S. Indicators to support an ecosystem approach to fisheries. Fish Fish. 2005, 6, 212–232. [Google Scholar] [CrossRef]
- Lleonart, J.; Maynou, F. Fish stock assessments in the Mediterranean: State of the art. Sci. Mar. 2003, 67, 37–49. [Google Scholar] [CrossRef]
- Tzanatos, E.; Somarakis, S.; Tserpes, G.; Koutsikopoulos, C. Identifying and classifying small-scale fisheries métiers in the Mediterranean: A case study in the Patraikos Gulf, Greece. Fish. Res. 2006, 81, 158–168. [Google Scholar] [CrossRef]
- Tzanatos, E.; Castro, J.; Forcada, A.; Matić-Skoko, S.; Gaspar, M.; Koutsikopoulos, C. A Métier-Sustainability-Index (MSI25) to evaluate fisheries components: Assessment of cases from data-poor fisheries from southern Europe. ICES J. Mar. Sci. 2013, 70, 78–98. [Google Scholar] [CrossRef]
- Guyader, O.; Berthou, P.; Koutsikopoulos, C.; Alban, F.; Demanèche, S.; Gaspar, M.B.; Eschbaum, R.; Fahy, E.; Tully, O.; Reynal, L.; et al. Small scale fisheries in Europe: A comparative analysis based on a selection of case studies. Fish. Res. 2013, 140, 1–13. [Google Scholar] [CrossRef]
- Lloret, J.; Cowx, I.G.; Cabral, H.; Castro, M.; Font, T.; Gonçalves, J.M.S.; Gordoa, A.; Hoefnagel, E.; Matić-Skoko, S.; Mikkelsen, E.; et al. Small-scale coastal fisheries in European Seas are not what they were: Ecological, social and economic changes. Mar. Policy 2018, 98, 176–186. [Google Scholar] [CrossRef]
- Grati, F.; Aladžuz, A.; Azzurro, E.; Bolognini, L.; Carbonara, P.; Çobani, M.; Domenichetti, F.; Dragičević, B.; Dulčić, J.; Durović, M.; et al. Seasonal dynamics of small-scale fisheries in the Adriatic Sea. Mediterr. Mar. Sci. 2018, 19, 21–35. [Google Scholar] [CrossRef]
- Marin, J. Exploitation, biologie et dymanique du stock de langouste rouge de corse Panulirus elephas Fabricus. Ph.D. Thesis, Université d’Aix-Marseille II, Marseille, France, 1987. [Google Scholar]
- Quetglas, A.; Gaamour, A.; Reñones, O.; Missaoui, H.; Zarrouk, T.; Elabed, A.; Goñi, R. Common spiny lobster (Palinurus elephas Fabricius 1787) fisheries in the western Mediterranean: A comparison of Spanish and Tunisian fisheries. Bolleti Soc. D’hist. Nat. Balear. 2004, 47, 63–80. [Google Scholar]
- Gönülal, O. Spiny lobster (Palinurus elephas Fabricius, 1787) and common lobster (Homarus gammarus Linnaeus, 1758) fishing in the Agean Sea. In The Aegean Sea Marine Biodiversity, Fisheries, Conservation and Governance; Katağan, T., Tokaç, A., Beşiktepe, Ş., Öztürk, B., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2015; p. 714. ISBN 978-975-8825-33-2. [Google Scholar]
- Lotze, H.K.; Coll, M.; Dunne, J.A. Historical Changes in Marine Resources, Food-web Structure and Ecosystem Functioning in the Adriatic Sea, Mediterranean. Ecosystems 2011, 14, 198–222. [Google Scholar] [CrossRef]
- FAO Fisheries & Aquaculture—Species Fact Sheets—Homarus gammarus (Linnaeus, 1758). Available online: http://www.fao.org/fishery/species/2648/en (accessed on 26 February 2020).
- Prodöhl, P.A.; Jørstad, K.E.; Triantafyllidis, A.; Katsares, V.; Triantaphyllidis, C. European lobster-Homarus gammarus. Genetic Impact of Aquaculture Activities on Native Populations. Genimpact Final Scientific Report (EU contract n. RICA-CT-2005-022802). 2007, pp. 91–98. Available online: http://genimpact.imr.no/ (accessed on 9 April 2021).
- Pere, A.; Marengo, M.; Lejeune, P.; Durieux, E.D.H. Evaluation of Homarus gammarus (Crustacea: Decapoda: Nephropidae) catches and potential in a mediterranean small-scale fishery. Sci. Mar. 2019, 83, 69–77. [Google Scholar] [CrossRef]
- Goñi, R.; Latrouite, D. Review of the biology, ecology and fisheries of Palinurus spp. species of European waters: Palinurus elephas (Fabricius, 1787) and Palinurus mauritanicus (Gruvel, 1911). Cah. Biol. Mar. 2005, 46, 127–142. [Google Scholar]
- Lloret, J.; Riera, V. Evolution of a mediterranean coastal zone: Human impacts on the marine environment of Cape Creus. Environ. Manag. 2008, 42, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Pere, A. Déclin des Populations de Langouste Rouge et Baisse de la Ressource Halieutique en Corse: Causes et Perspectives. Ph.D. Thesis, Université de Corse, Corte, France, 2012; p. 478. [Google Scholar]
- Dow, R.L. The Clawed Lobster Fisheries. In The Biology and Management of Lobsters; Elsevier Inc.: Amsterdam, The Netherlands, 1980; Volume 2, pp. 265–316. ISBN 9780080917344. [Google Scholar]
- Cobb, J.S.; Castro, K.M. Homarus Species. In Lobsters: Biology, Management, Aquaculture and Fisheries; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 310–339. [Google Scholar]
- Browne, R.M.; Mercer, J.P.; Duncan, M.J. An historical overview of the Republic of Ireland’s lobster (Homarus gammarus Linnaeus) fishery, with reference to European and North American (Homarus americanus Milne Edwards) lobster landings. In Coastal Shellfish A Sustainable Resource; Springer: Amsterdam, The Netherlands, 2001; pp. 49–62. [Google Scholar]
- European Union. Council Regulation (EC) No 1967/2006 of 21 December 2006 Concerning Management Measures for the Sustainable Exploitation of Fishery Resources in the Mediterranean Sea, Amending Regulation (EEC) No 2847/93 and Repealing Regulation (EC) No 1626/94; European Union: Brussels, Belgium, 2006. [Google Scholar]
- Coleman, M.T.; Agnalt, A.-L.; Emmerson, J.; Laurens, M.; Porter, J.S.; Bell, M.C. From the Adriatic to Northern Norway—geographic differences in moult increment and moult probability of the European lobster (Homarus gammarus), across the natural range. ICES J. Mar. Sci. 2020. [Google Scholar] [CrossRef]
- Pavičić, M.; Žužul, I.; Matić-Skoko, S.; Triantafyllidis, A.; Grati, F.; Durieux, E.D.H.; Celić, I.; Šegvić-Bubić, T. Population Genetic Structure and Connectivity of the European Lobster Homarus gammarus in the Adriatic and Mediterranean Seas. Front. Genet. 2020, 11, 1540. [Google Scholar] [CrossRef] [PubMed]
- Debuse, V.J.; Addison, J.T.; Reynolds, J.D. Morphometric variability in UK populations of the European lobster. J. Mar. Biol. Assoc. U. K. 2001, 81, 469–474. [Google Scholar] [CrossRef]
- Galparsoro, I.; Borja, Á.; Bald, J.; Liria, P.; Chust, G. Predicting suitable habitat for the European lobster (Homarus gammarus), on the Basque continental shelf (Bay of Biscay), using Ecological-Niche Factor Analysis. Ecol. Model. 2009, 220, 556–567. [Google Scholar] [CrossRef]
- Lizárraga-Cubedo, H.A.; Tuck, I.; Bailey, N.; Pierce, G.J.; Zuur, A.F.; Bova, D. Scottish lobster fisheries and environmental variability. ICES J. Mar. Sci. 2015, 72, i211–i224. [Google Scholar] [CrossRef]
- Tully, O.; Roantree, V.; Robinson, M. Maturity, fecundity and reproductive potential of the European lobster (Homarus gammarus) in Ireland. J. Mar. Biol. Assoc. U. K. 2001, 81, 61–68. [Google Scholar] [CrossRef]
- Skerritt, D.J.; Robertson, P.A.; Mill, A.C.; Polunin, N.V.C.; Fitzsimmons, C. Fine-scale movement, activity patterns and home-ranges of European lobster Homarus gammarus. Mar. Ecol. Prog. Ser. 2015, 536, 203–219. [Google Scholar] [CrossRef]
- Phillips, B.F. Lobsters: Biology, Management, Aquaculture and Fisheries; Phillips, B.F., Ed.; John Wiley & Sons, Ltd.: Oxford, UK, 2013; ISBN 9781118517444. [Google Scholar]
- Triantafyllidis, A.; Apostolidis, A.P.; Katsares, V.; Kelly, E.; Mercer, J.; Hughes, M.; Jørstad, K.E.; Tsolou, A.; Hynes, R.; Triantaphyllidis, C. Mitochondrial DNA variation in the European lobster (Homarus gammarus) throughout the range. Mar. Biol. 2005, 146, 223–235. [Google Scholar] [CrossRef]
- Ellis, C.D.; Hodgson, D.J.; Daniels, C.L.; Collins, M.; Griffiths, A.G.F. Population genetic structure in European lobsters: Implications for connectivity, diversity and hatchery stocking. Mar. Ecol. Prog. Ser. 2017, 563, 123–137. [Google Scholar] [CrossRef][Green Version]
- Jenkins, T.L.; Ellis, C.D.; Triantafyllidis, A.; Stevens, J.R. Single nucleotide polymorphisms reveal a genetic cline across the north-east Atlantic and enable powerful population assignment in the European lobster. Evol. Appl. 2019, 12, 1881–1899. [Google Scholar] [CrossRef]
- Devescovi, M.; Lucu, Č. Growth patterns of the lobster Homarus gammarus in the North Adriatic. Period. Biol. 2000, 102, 189–193. [Google Scholar]
- Martínez-Calderón, R.; Lozano-Álvarez, E.; Briones-Fourzán, P. Morphometric relationships and seasonal variation in size, weight, and a condition index of post-settlement stages of the Caribbean spiny lobster. PeerJ 2018, 2018, 1–22. [Google Scholar] [CrossRef]
- Zheng, J.; Murphy, M.C.; Kruse, G.H. A length-based population model and stock-recruitment relationships for red king crab, Paralithodes camtschaticus, in Bristol Bay, Alaska. Can. J. Fish. Aquat. Sci. 1995, 52, 1229–1246. [Google Scholar] [CrossRef]
- Santos, M.N.; Gaspar, M.B.; Vasconcelos, P.; Monteiro, C.C. Weight-length relationships for 50 selected fish species of the Algarve coast (southern Portugal). Fish. Res. 2002, 59, 289–295. [Google Scholar] [CrossRef]
- Baggio, M. Estimating weight-length relationships without individual weight data: An application to the American lobster (Homarus americanus) fishery of Long Island Sound. Rev. Fish Biol. Fish. 2011, 21, 771–777. [Google Scholar] [CrossRef][Green Version]
- Lizárraga-Cubedo, H.A.; Tuck, I.; Bailey, N.; Pierce, G.J.; Kinnear, J.A.M. Comparisons of size at maturity and fecundity of two Scottish populations of the European lobster, Homarus gammarus. Fish. Res. 2003, 65, 137–152. [Google Scholar] [CrossRef]
- Conan, G.Y.; Comeau, M.; Moriyasu, M. Are Morphometrical Approaches Appropriate to Establish Size at Maturity for Male American Lobster, Homarus americanus? J. Crustac. Biol. 2001, 21, 937–947. [Google Scholar] [CrossRef]
- Aubry, F.B.; Cossarini, G.; Acri, F.; Bastianini, M.; Bianchi, F.; Camatti, E.; De Lazzari, A.; Pugnetti, A.; Solidoro, C.; Socal, G. Plankton communities in the northern Adriatic Sea: Patterns and changes over the last 30 years. Estuar. Coast. Shelf Sci. 2012, 115, 125–137. [Google Scholar] [CrossRef]
- Djakovac, T.; Degobbis, D.; Supić, N.; Precali, R. Marked reduction of eutrophication pressure in the northeastern Adriatic in the period 2000–2009. Estuar. Coast. Shelf Sci. 2012, 115, 25–32. [Google Scholar] [CrossRef]
- Shaltout, M.; Omstedt, A. Recent sea surface temperature trends and future scenarios for the Mediterranean Sea. Oceanologia 2014, 56, 411–443. [Google Scholar] [CrossRef]
- Vilibić, I.; Zemunik, P.; Šepić, J.; Dunić, N.; Marzouk, O.; Mihanović, H.; Denamiel, C.; Precali, R.; Djakovac, T. Present climate trends and variability in thermohaline properties of the northern Adriatic shelf. Ocean Sci. 2019, 15, 1351–1362. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259. [Google Scholar] [CrossRef] [PubMed]
- Ricker, W.E. Linear Regressions in Fishery Research. J. Fish. Res. Board Can. 1973, 30, 409–434. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Hoboken, NJ, USA, 1999; ISBN 9780130815422. [Google Scholar]
- Hartnoll, R.G. Growth. In The Biology of Crustacea: Embriology, Morphology and Genetics; Abele, L.G., Ed.; Academic Press: New York, NY, USA, 1982; pp. 11–196. ISBN 0-12-106402-6. [Google Scholar]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Baeza-Martínez, K.; Lozano-Álvarez, E. Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reef lagoon: Are changes over a 10-year span related to the emergence of Panulirus argus Virus 1 (PaV1)? J. Exp. Mar. Biol. Ecol. 2009, 370, 82–88. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2015. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 1 July 2019).
- Tully, O. Impact of the v-notch technical conservation measure on reproductive potential in a lobster (Homarus gammarus L.) fishery in Ireland. Mar. Freshw. Res. 2001, 52, 1551–1557. [Google Scholar] [CrossRef]
- Cushing, D.H. Climate and Fisheries; Academic Press: New York, NY, USA, 1982; ISBN 9780121997205. [Google Scholar]
- Matić-Skoko, S.; Stagličić, N.; Pallaoro, A.; Kraljević, M.; Dulčić, J.; Tutman, P.; Dragičević, B. Effectiveness of conventional management in Mediterranean type artisanal fisheries. Estuar. Coast. Shelf Sci. 2011, 91, 314–324. [Google Scholar] [CrossRef]
- Stagličić, N.; Matić-Skoko, S.; Pallaoro, A.; Grgičević, R.; Kraljević, M.; Tutman, P.; Dragičević, B.; Dulčić, J. Long-term trends in the structure of eastern Adriatic littoral fish assemblages: Consequences for fisheries management. Estuar. Coast. Shelf Sci. 2011, 94, 263–271. [Google Scholar] [CrossRef]
- Matić-Skoko, S.; Stagličić, N.; Blažević, D.; Šiljić, J.; Iritani, D. Croatian marine fisheries (Adriatic Sea): 1950-2013. In Global atlas of Marine Fisheries—A Critical Appraisal of Catches and Ecosystem Impacts; Pauly, D., Zeller, D., Eds.; Island Press: Washington, DC, USA, 2016; p. 232. [Google Scholar]
- Caputi, N.; Lestang, S.; Frusher, S.; Wahle, R.A. The Impact of Climate Change on Exploited Lobster Stocks. In Lobsters: Biology, Management, Aquaculture and Fisheries; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 84–112. [Google Scholar]
- Green, B.S.; Gardner, C.; Hochmuth, J.D.; Linnane, A. Environmental effects on fished lobsters and crabs. Rev. Fish Biol. Fish. 2014, 24, 613–638. [Google Scholar] [CrossRef]
- Ben Rais Lasram, F.; Guilhaumon, F.; Albouy, C.; Somot, S.; Thuiller, W.; Mouillot, D. The Mediterranean Sea as a ‘cul-de-sac’ for endemic fishes facing climate change. Glob. Chang. Biol. 2010, 16, 3233–3245. [Google Scholar] [CrossRef]
- Pranovi, F.; Caccin, A.; Franzoi, P.; Malavasi, S.; Zucchetta, M.; Torricelli, P. Vulnerability of artisanal fisheries to climate change in the Venice Lagoon. J. Fish Biol. 2013, 83, 847–864. [Google Scholar] [CrossRef]
- Ministry of Agriculture. Pravilnik o Zaštiti Riba i Drugih Morskih Organizama. Narodne Novine 42/2016. No. 1096. (In Croatian). Available online: https://narodne-novine.nn.hr/eli/sluzbeni/2016/42/1096 (accessed on 9 April 2021).
- Wahle, R.A.; Castro, K.M.; Tully, O.; Cobb, J.S. Homarus. In Lobsters: Biology, Management, Aquaculture and Fisheries; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 221–258. [Google Scholar]
- McCleese, D.W.; Wildner, D.G. Overview of the Inshore Lobster Resources in the Scotia-Fundy Region. In Canadian Atlantic Fisheries Scientific Advisory Committee (CAFSAC) Research Document 85; CAFSAC: Halifax, NS, Canada, 1958; p. 20. [Google Scholar]
- Krouse, J.S. Performance and selectivity of trap fisheries for crustaceans. In Marine Invertebrate Fisheries: Their Assessment and Management; Caddy, J.F., Ed.; Wiley: New York, NY, USA, 1989; pp. 307–325. [Google Scholar]
- Latrouite, D. Le homard (Homarus gammarus) du nord golfe de Gascogne-Manche ouest (divisions VIIe + VIIIa du CIEM). In Contrat Ifremer/MAPA—Réf. 99-I1-03-01 Rapport Final: 1–13; Ifremer: Brest, France, 2001. [Google Scholar]
- Bennet, D.B.; Howard, A.E. Estimates of lobster (Homarus gammarus) fecundity from East and West Britain. ICES Document CM 1987/K: 47; Oxford University Press: Oxford, UK, 1987. [Google Scholar]
- Laurans, M.; Fifas, S.; Demaneche, S.; Brérette, S.; Debec, O. Modelling seasonal and annual variation in size at functional maturity in the European lobster (Homarus gammarus) from self-sampling data. ICES J. Mar. Sci. 2009, 66, 1892–1898. [Google Scholar] [CrossRef][Green Version]
- Schoeman, D.S.; Cockcroft, A.C.; Van Zyl, D.L.; Goosen, P.C. South African rock lobster Jasus lalandii commercial fishery. S. Afr. J. Mar. Sci. 2010, 24, 37–48. [Google Scholar] [CrossRef]
- Ministry of Agriculture. Pravilnik o Obavljanju Gospodarskog Ribolova na Moru Mrežama Stajaćicama, Klopkastim, Udičarskim i Probodnim Ribolovnim Alatima te Posebnim Načinima Ribolova. Narodne Novine 84/2015. No. 1640. (In Croatian). Available online: https://narodne-novine.nn.hr/eli/sluzbeni/2015/84/1640 (accessed on 9 April 2021).
- Shelton, P.M.J.; Shelton, R.G.J.; Richards, P.R. Eye development in relation to molt stage in the European lobster Homarus gammarus. J. Cons. 1981, 39, 239–243. [Google Scholar] [CrossRef]
- Fütterer, D.; Paul, J. Recent and Pleistocene sediments off the Istrian coast (Northern Adriatic). Senckenbergiana Marit. 1976, 8, 1–21. [Google Scholar]
- Smith, I.P.; Jensen, A.C.; Collins, K.J.; Mattey, E.L. Movement of wild European lobsters Homarus gammarus in natural habitat. Mar. Ecol. Prog. Ser. 2001, 222, 177–186. [Google Scholar] [CrossRef]
- Uglem, I.; Belchier, M.; Svasand, T. Age determination of European lobsters (Homarus gammarus L.) by histological quantification of lipofuscin. J. Crustac. Biol. 2005, 25, 95–99. [Google Scholar] [CrossRef]
- Kilada, R.; Driscoll, J.G. Age determination in crustaceans: A review. Hydrobiologia 2017, 799, 21–36. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Merilä, J. Heritable variation and evolution under favourable and unfavourable conditions. Trends Ecol. Evol. 1999, 14, 96–101. [Google Scholar] [CrossRef]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Ward, R.D.; Woodwark, M.; Skibinski, D.O.F. A comparison of genetic diversity levels in marine, freshwater, and anadromous fishes. J. Fish Biol. 1994, 44, 213–232. [Google Scholar] [CrossRef]
- Ribeiro, F.B.; Matthews Cascon, H.; Bezerra, L.E.A. Morphometric sexual maturity and allometric growth of the crab Sesarma rectum Randall, 1840 (Crustacea: Sesarmidae) in an impacted tropical mangrove in northeast Brazil. Lat. Am. J. Aquat. Res. 2013, 41, 361–368. [Google Scholar] [CrossRef]
- Glamuzina, L.; Conides, A.; Mancinelli, G.; Dobroslavić, T.; Bartulović, V.; Matić-Skoko, S.; Glamuzina, B. Population Dynamics and Reproduction of Mediterranean Green Crab Carcinus aestuarii in Parila Lagoon (Neretva Estuary, Adriatic Sea, Croatia) as Fishery Management Tools. Mar. Coast. Fish. 2017, 9, 260–270. [Google Scholar] [CrossRef]
- Horsford, I.; Simon, H.; Archibald, M.; Webber, J.; Joseph, T. Biology, status, and current management of the Caribbean Spiny Lobster (Panulirus argus). Gulf Caribb. Fish. Inst. 2014, 66, 423–433. [Google Scholar]
- Kampouris, T.E.; Koutsoubas, D.; Milenkova, D.; Economidis, G.; Tamvakidis, S.; Batjakas, I.E. New data on the biology and fisheries of the threatened Palinurus elephas (Fabricius, 1787) (Decapoda, achelata, Palinuridae) from the north-west Aegean sea, Greece. Water 2020, 12, 2390. [Google Scholar] [CrossRef]
- French McCay, D.; Gibson, M.; Cobb, J. Scaling restoration of American lobsters: Combined demographic and discounting model for an exploited species. Mar. Ecol. Prog. Ser. 2003, 264, 177–196. [Google Scholar] [CrossRef]
- Steinback, S.R.; Allen, R.B.; Thunberg, E. The benefits of rationalization: The case of the American lobster fishery. Mar. Resour. Econ. 2008, 23, 37–63. [Google Scholar] [CrossRef]
- Sheehy, M.R.J.; Bannister, R.C.A.; Wickins, J.F.; Shelton, P.M.J. New perspectives on the growth and longevity of the European lobster (Homarus gammarus). Can. J. Fish. Aquat. Sci. 1999, 56, 1904–1915. [Google Scholar] [CrossRef]
- Radhakrishnan, E.V.; Thangaraja, R.; Vijayakumaran, M. Ontogenetic changes in morphometry of the spiny lobster, Panulirus homarus homarus (Linnaeus, 1758) from southern Indian coast. J. Mar. Biol. Assoc. India 2015, 57, 5–13. [Google Scholar] [CrossRef]
- Ball, B.; Linnane, A.; Munday, B.; Browne, R.; Mercer, J.P. The effect of cover on in situ predation in early benthic phase European lobster Homarus gammarus. J. Mar. Biol. Assoc. U. K. 2001, 81, 639–642. [Google Scholar] [CrossRef]
- Landers, D.F.; Keser, M.; Saila, S.B. Changes in female lobster (Homarus americanus) size at maturity and implications for the lobster resource in Long Island Sound, Connecticut. Mar. Freshw. Res. 2001, 52, 1283–1290. [Google Scholar] [CrossRef]
- Woodruff, J. Stock Assessment—An Evaluation of the Minimum Landing Size and the Pot Limitation Byelaw in the Northumberland Lobster Fishery; NIFCA: Northumberland, UK, 2017. [Google Scholar]
- Baudron, A.R.; Needle, C.L.; Rijnsdorp, A.D.; Tara Marshall, C. Warming temperatures and smaller body sizes: Synchronous changes in growth of North Sea fishes. Glob. Chang. Biol. 2014, 20, 1023–1031. [Google Scholar] [CrossRef]
- Le Bris, A.; Pershing, A.J.; Gaudette, J.; Pugh, T.L.; Reardon, K.M. Multi-scale quantification of the effects of temperature on size at maturity in the American lobster (Homarus americanus). Fish. Res. 2017, 186, 397–406. [Google Scholar] [CrossRef]
- The Shellfish Team. Lobster (Homarus gammarus) Cefas Stock Status Report 2014; Center for Environment Fisheries & Aquaculture Science: Lowestoft, UK, 2014.
- Haarr, M.L.; Sainte-Marie, B.; Comeau, M.; Tremblay, M.J.; Rochette, R. Female American lobster (Homarus americanus) size-at-maturity declined in Canada during the 20th and early 21st centuries. Can. J. Fish. Aquat. Sci. 2018, 75, 908–924. [Google Scholar] [CrossRef]
- Grabowski, J.H.; Clesceri, E.J.; Baukus, A.J.; Gaudette, J.; Weber, M.; Yund, P.O. Use of Herring Bait to Farm Lobsters in the Gulf of Maine. PLoS ONE 2010, 5, e10188. [Google Scholar] [CrossRef]
- Turner, R.; Hardy, M.; Green, J.; Polunin, N. Defining the Northumberland Lobster Fishery; Newcastle University: London, UK, 2009. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).