
Fishing Methods Matter: Comparing the Community and Trait Composition of the Dai (Bagnet) and Gillnet Fisheries in the Tonle Sap River in Southeast Asia


 ,
,  and
and 
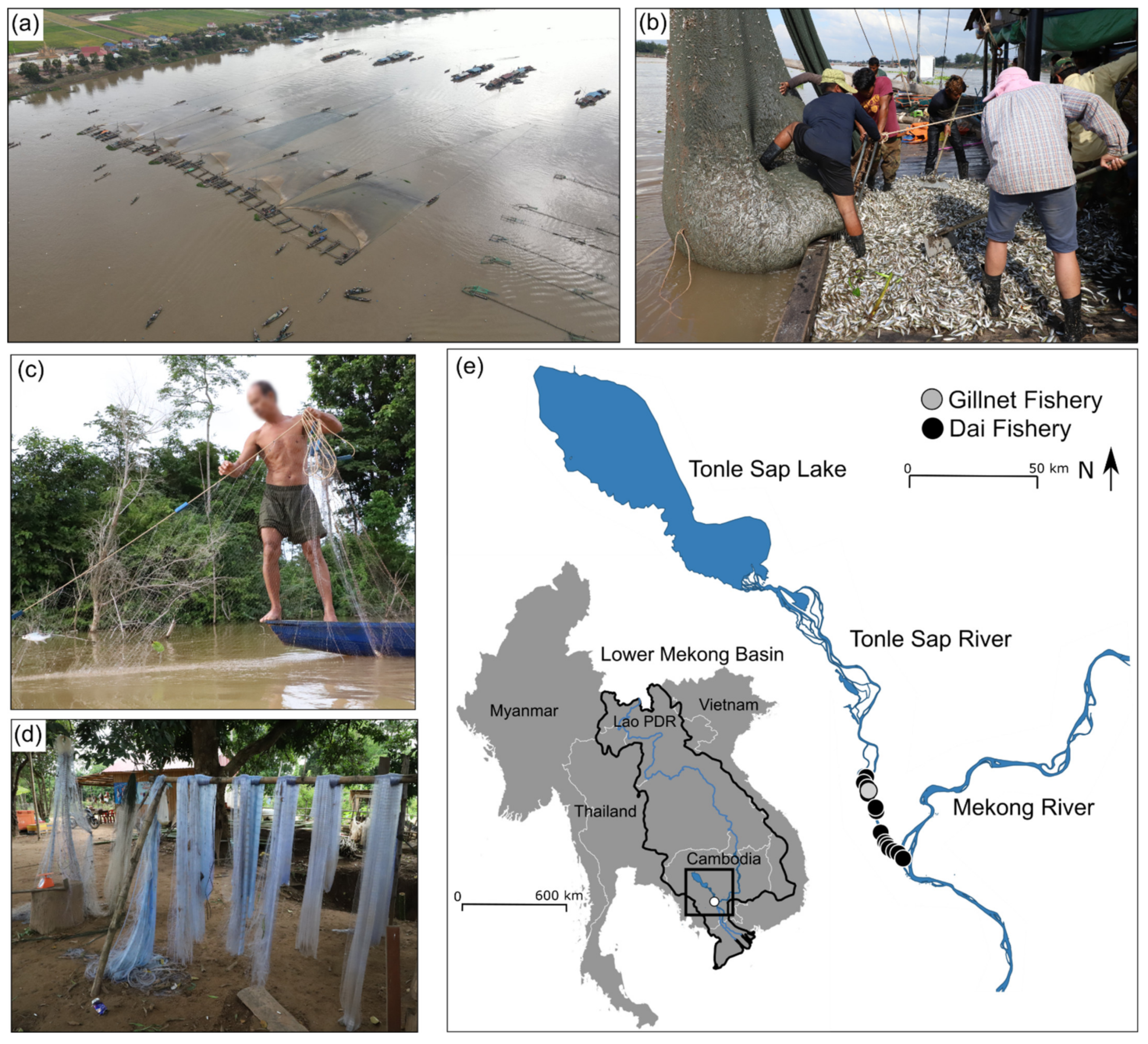{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
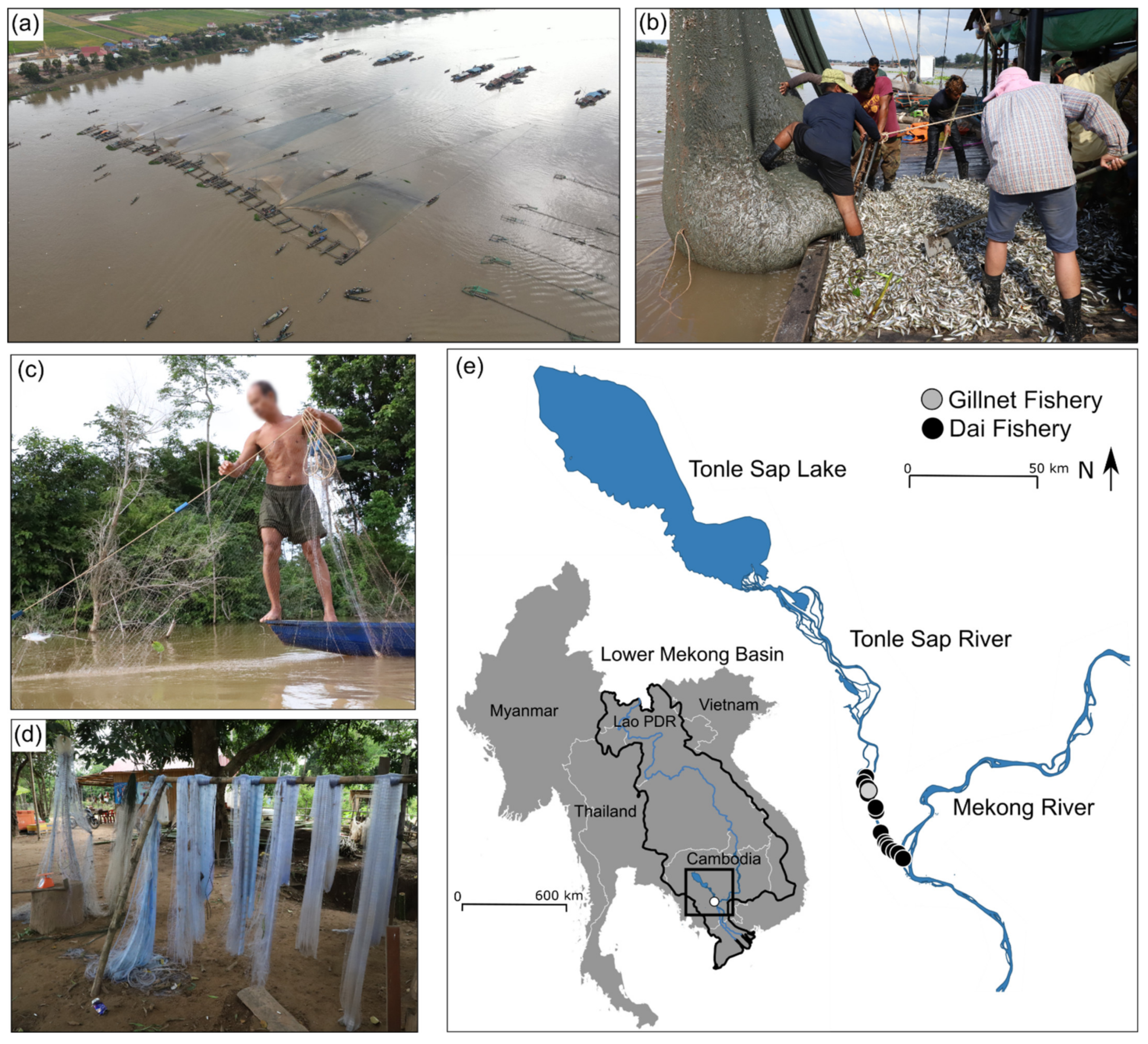
2.1. Fish Catch and Trait Data
2.2. Comparing Annual Catch to Flooding Magnitude
2.3. Community Analyses to Compare the Dai and Gillnet Fisheries
2.4. Resource Selection Ratios to Quantify Gillnet Selectivity Relative to the Dai Fishery
2.5. Trait Changes in the Dai and Gillnet Fishery
3. Results
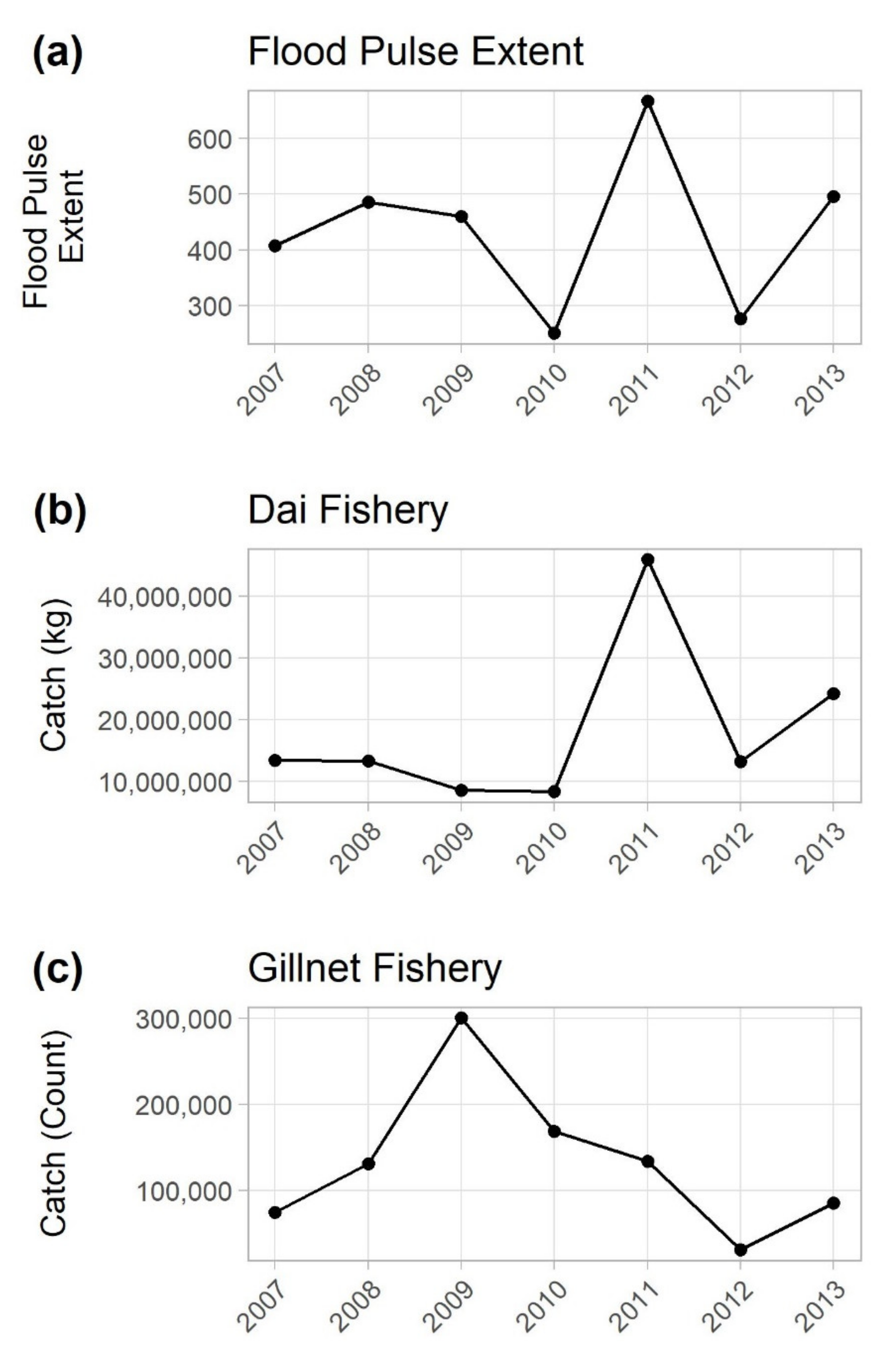
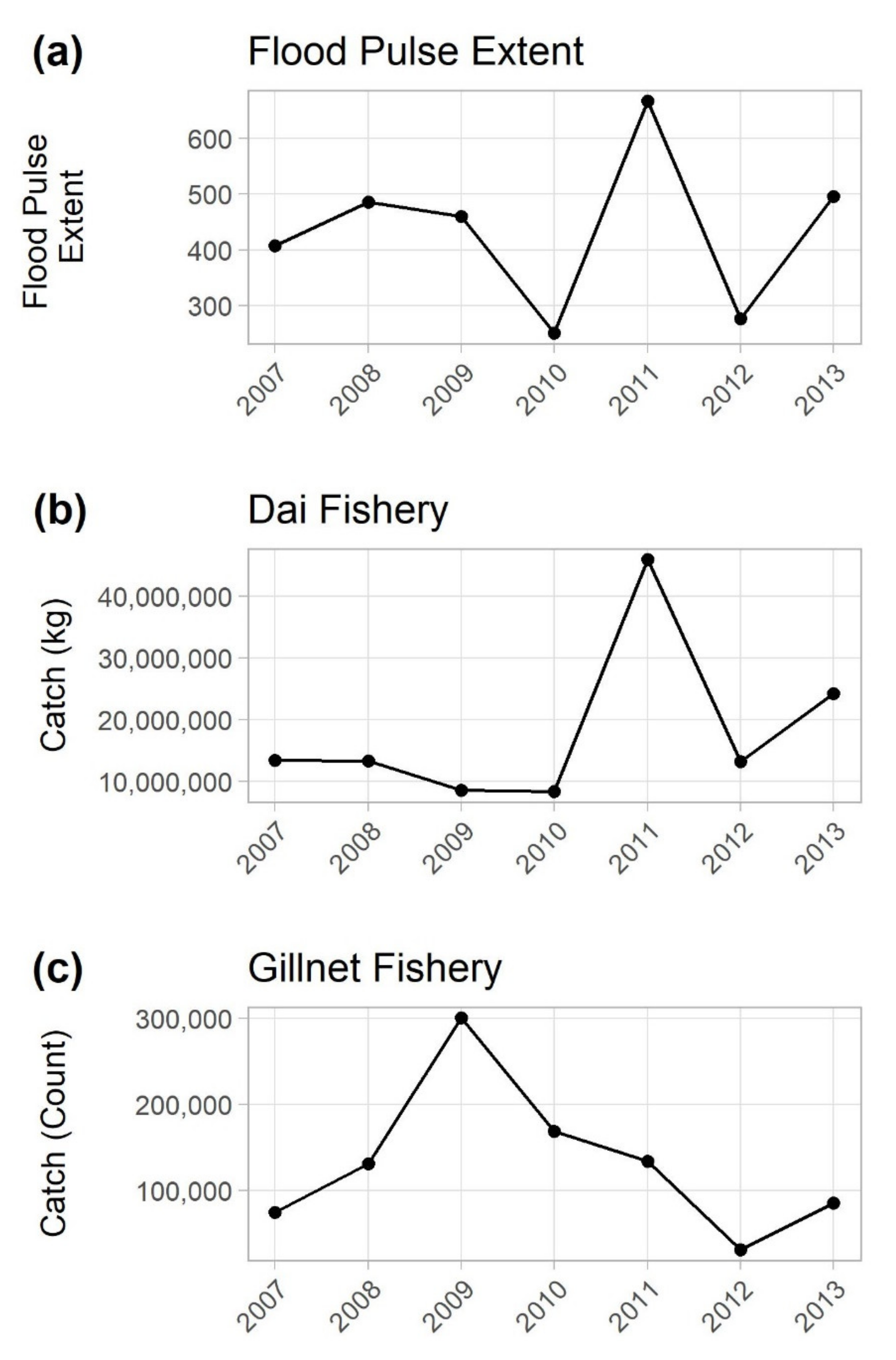
3.1. Comparing Annual Catch to Flooding Magnitude
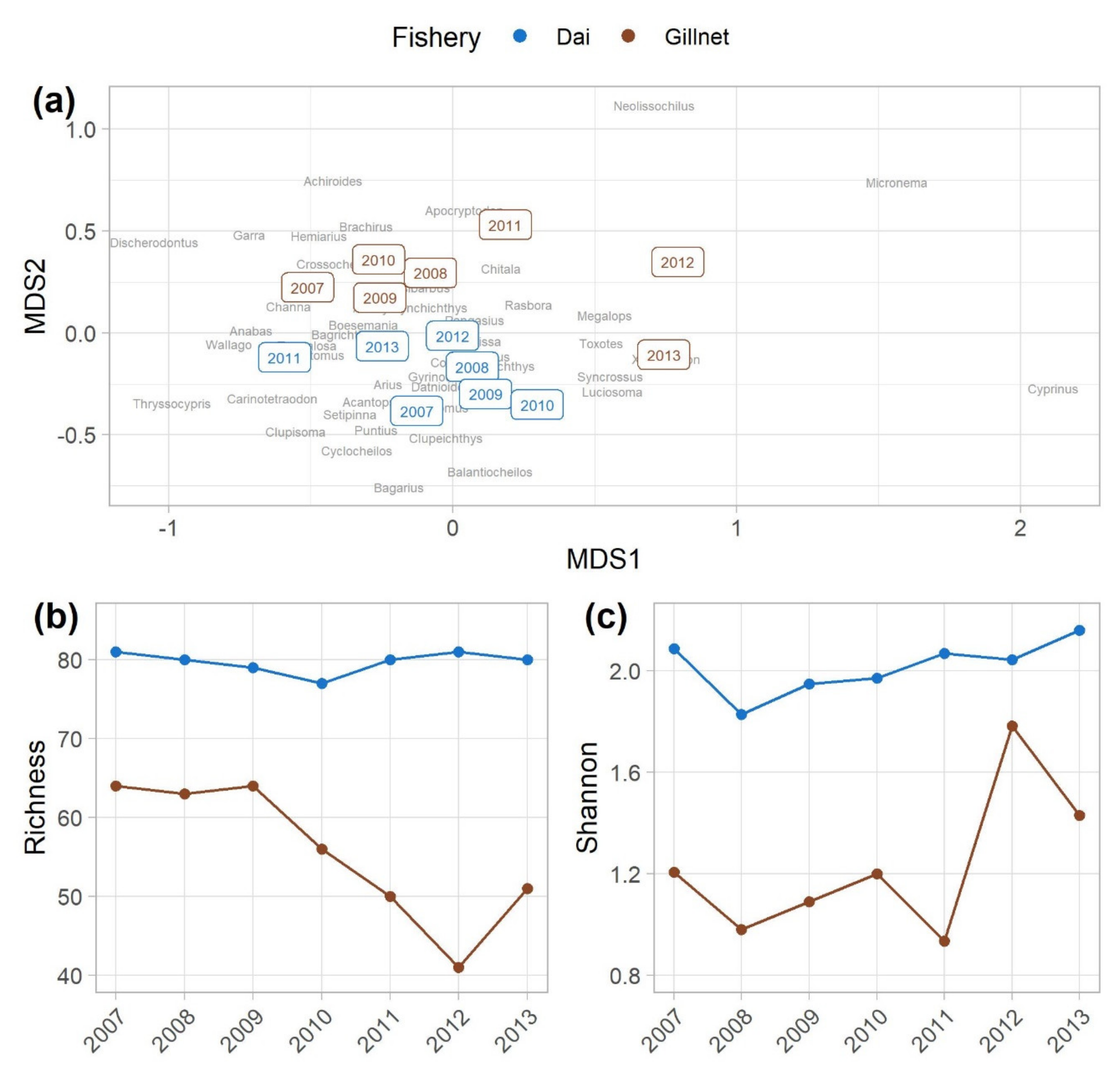
3.2. Community Analyses to Compare the Dai and Gillnet Fisheries
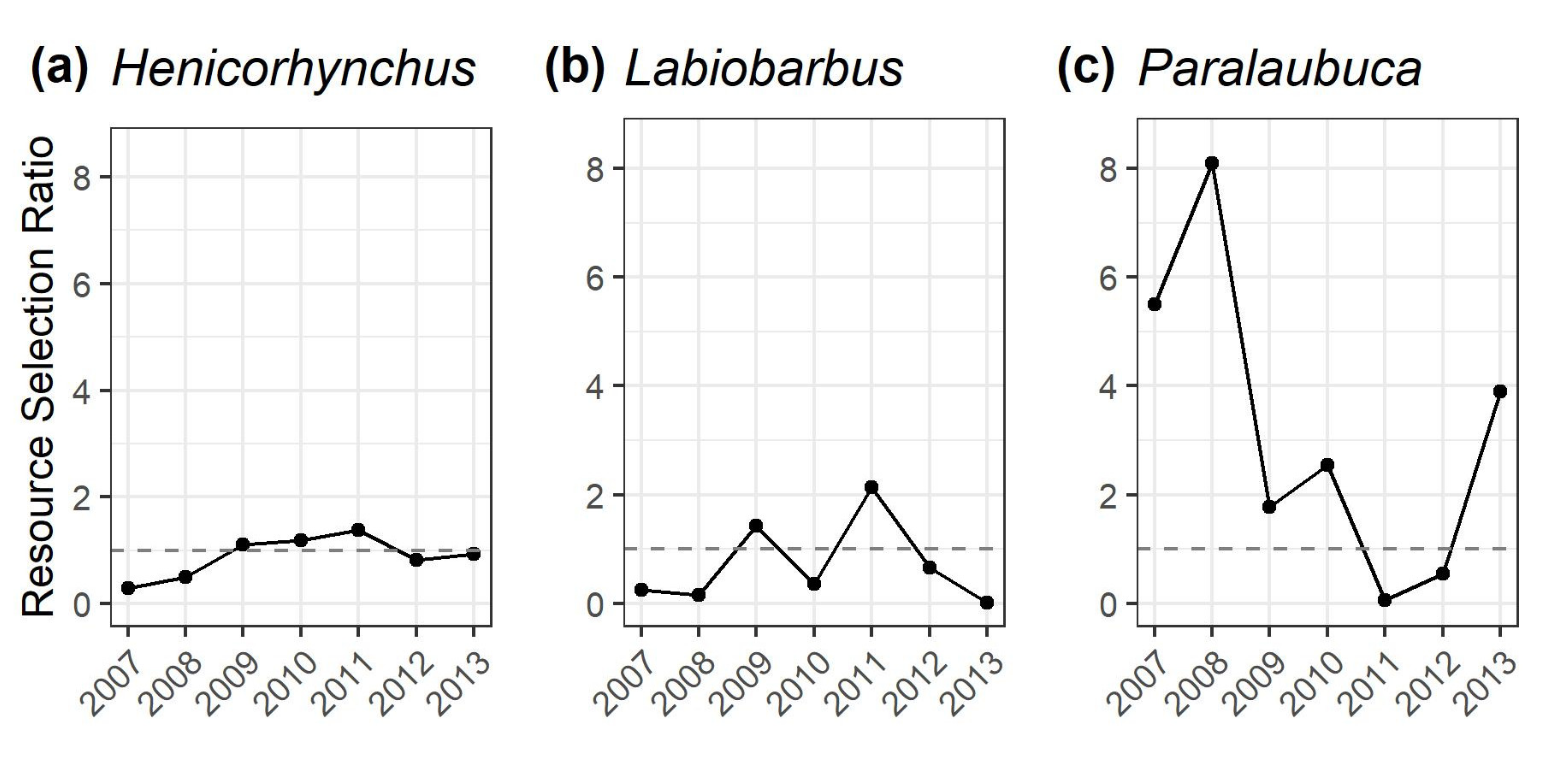
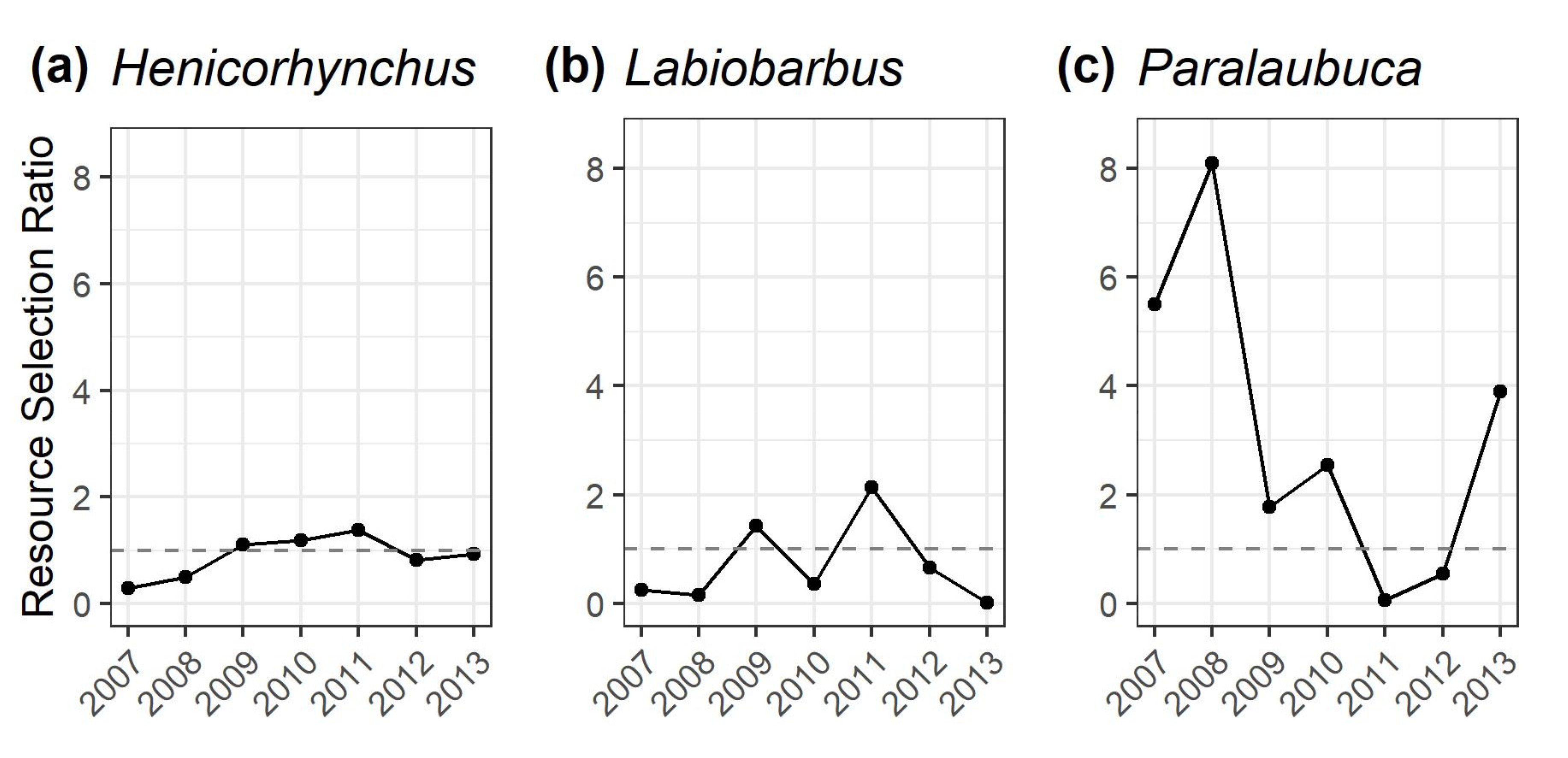
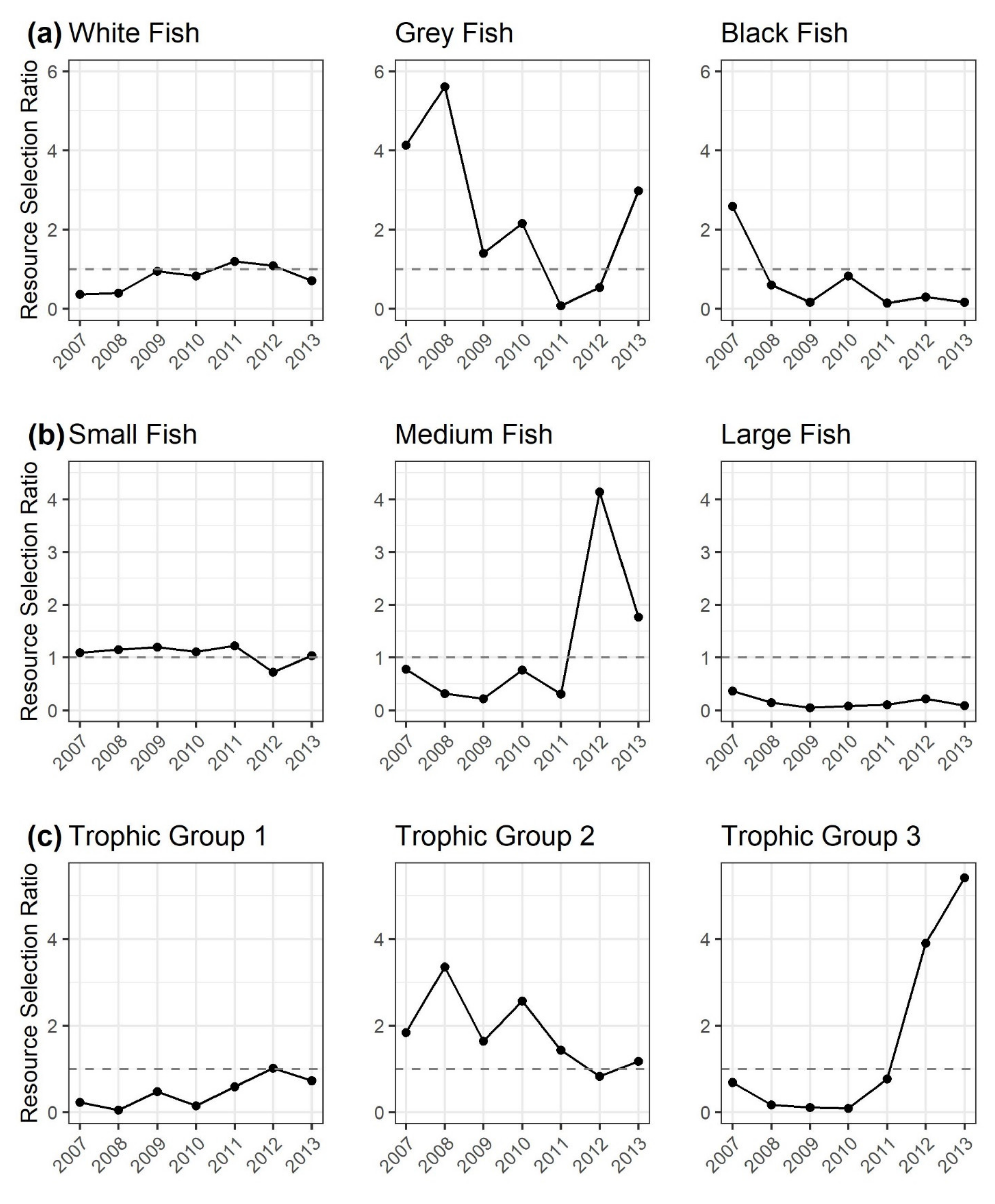
3.3. Resource Selection Ratios to Quantify Gillnet Selectivity Relative to the Dai Fishery
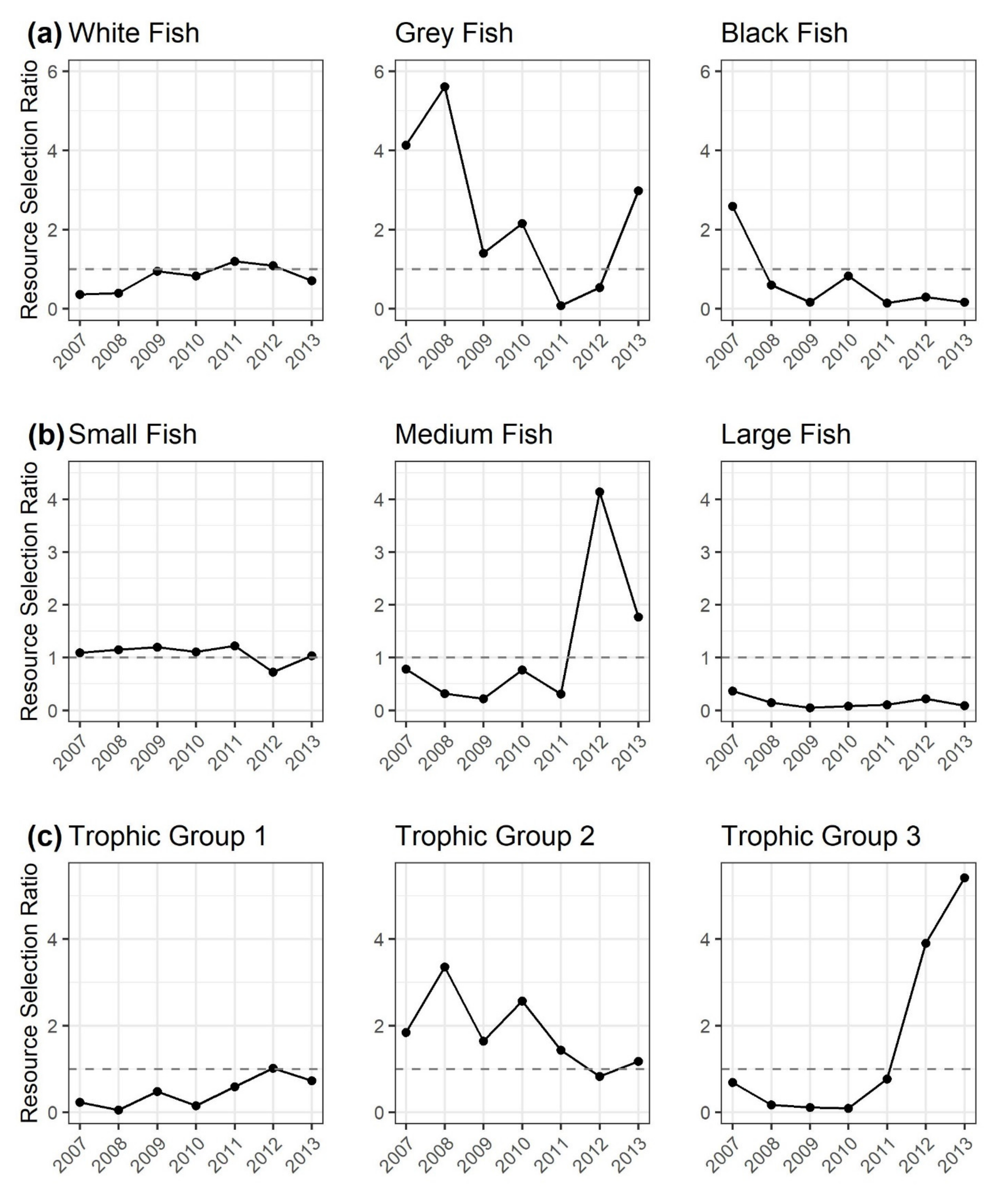
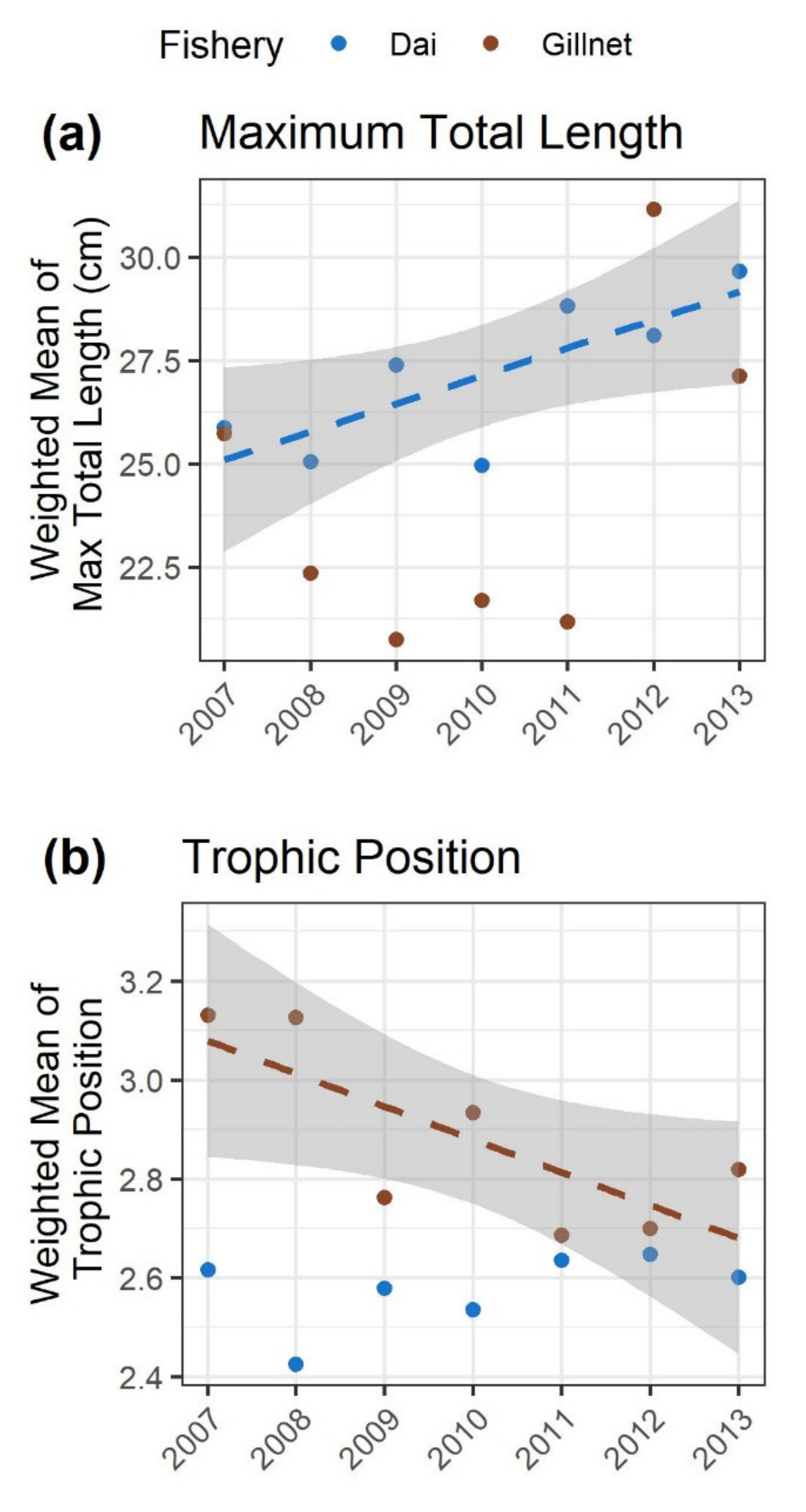
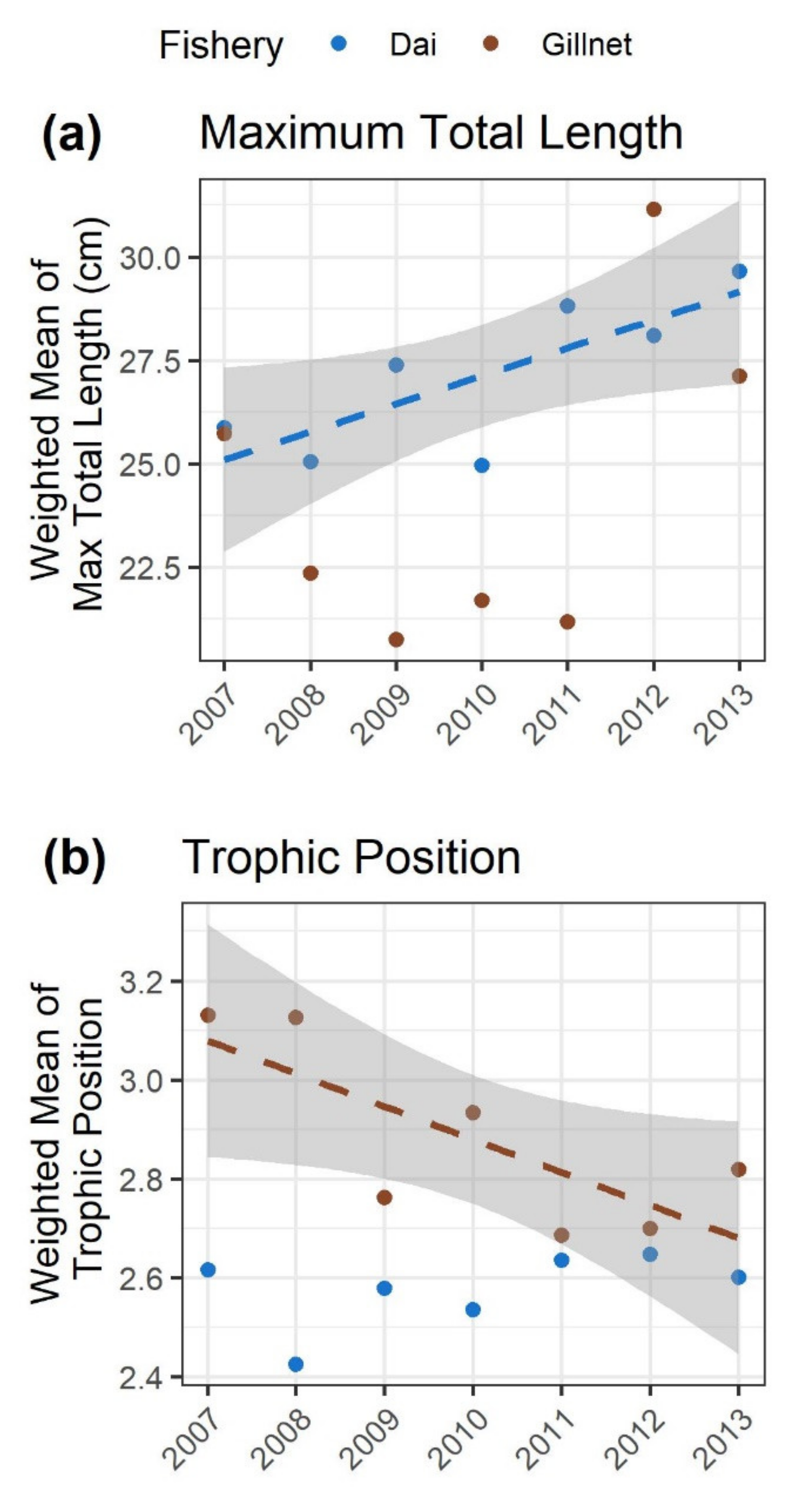
3.4. Trait Changes in the Dai and Gillnet Fishery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humphries, A.T.; Gorospe, K.D.; Carvalho, P.G.; Yulianto, I.; Kartawijaya, T.; Campbell, S.J. Catch composition and selectivity of fishing gears in a multi-species Indonesian coral reef fishery. Front. Mar. Sci. 2019, 6, 378. [Google Scholar] [CrossRef]
- Huse, I.; Løkkeborg, S.; Soldal, A.V. Relative selectivity in trawl, longline and gillnet fisheries for cod and haddock. ICES J. Mar. Sci. 2000, 57, 1271–1282. [Google Scholar] [CrossRef]
- Hortle, B.K.G.; Pengbun, N.; Rady, H.; Sopha, L. Trends in the Cambodian Dai fishery: Floods and fishing pressure. Catch Cult. 2004, 10, 7–9. [Google Scholar]
- Hortle, K.G. Fisheries of the Mekong River Basin, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; ISBN 9780123740267. [Google Scholar]
- Baran, E.; Cheng, P.; Ly, V.; Nasielski, J.; Saray, S.; Touch, B.; Tress, J.; Kaing, K.; Tan, S. Fish Resources. In Atlas of Cambodia Socio-Economic Development and Environment; Save Cambodia’s Wildlife: Phnom Penh, Cambodia, 2014; pp. 37–48. [Google Scholar]
- Pin, K.; Nut, S.; Hogan, Z.; Chandra, S.; Saray, S.; Touch, B.; Chheng, P.; Ngor, P.B. Cambodian freshwater fish assemblage structure and distribution patterns: Using a large-scale monitoring network to understand the dynamics and management implications of species clusters in a global biodiversity hotspot. Water 2020, 12, 2506. [Google Scholar] [CrossRef]
- Poulsen, A.F.; Valbo-Jørgensen, J. Fish Migrations and Spawning Habits in the Mekong Mainstream a Survey Using Local Knowledge; Mekong River Commission: Vientiane, Laos, 2000. [Google Scholar]
- Poulsen, A.F.; Poeu, O.; Viravong, S.; Suntornratana, U.; Tung, N.T. Fish Migrations of the Lower Mekong River Basin: Implications for Development, Planning and Environmental Management; Mekong River Commission: Vientiane, Laos, 2002. [Google Scholar]
- Baran, E. Fish Migration Triggers in the Lower Mekong Basin and other Freshwater Tropical Systems; Mekong River Commission: Vientiane, Laos, 2006. [Google Scholar]
- McIntyre, P.B.; Liermann, C.A.R.; Revenga, C. Linking freshwater fishery management to global food security and biodiversity conservation. Proc. Natl. Acad. Sci. USA. 2016, 113, 12880–12885. [Google Scholar] [CrossRef] [Green Version]
- Hortle, K.G.; Bamrungrach, P. Fisheries Habitat and Yield in the Lower Mekong Basin; Mekong River Commission: Vientiane, Laos, 2015. [Google Scholar]
- Baran, E.; Myschowoda, C. Have Fish Catches Been Declining in the Mekong River Basin? In Modern Myths of the Mekong; Water & Development Publications Helsinki University of Technology: Espoo, Finland, 2008; pp. 55–64. ISBN 9789512291021. [Google Scholar]
- Campbell, I.C. Perceptions, data, and river management: Lessons from the Mekong River. Water Resour. Res. 2007, 43, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ngor, P.B.; McCann, K.S.; Grenouillet, G.; So, N.; McMeans, B.C.; Fraser, E.; Lek, S. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Sci. Rep. 2018, 8, 8947. [Google Scholar] [CrossRef] [Green Version]
- Hogan, Z.; Baird, I.G.; Radtke, R.; Vander Zanden, M.J. Long distance migration and marine habitation in the tropical Asian catfish, Pangasius krempfi. J. Fish Biol. 2007, 71, 818–832. [Google Scholar] [CrossRef]
- Baird, I.; Hogan, Z.; Phylaivanh, B.; Moyle, P. A Communal Fishery for the Migratory Catfish Pangasius macronema in the Mekong River. Asian Fish. Sci. 2001, 14, 25–41. [Google Scholar]
- Deap, L.; Degen, P.; Van Zalinge, N. Fishing gears of the Cambodian Mekong; MRC: Phnom Penh, Cambodia, 2003. [Google Scholar]
- Welcomme, R.L.; Valbo-Jorgensen, J.; Halls, A. Inland Fisheries Evolution and Management. Case Studies from Four Continents; FAO Fisheries and Aquaculture Technical Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Baran, E.; Van Zalinge, N.; Ngor, P.B. Floods, Floodplains and Fish production in the Mekong Basin: Present and Past Trends. In Proceedings of the Second Asian Wetlands Symposium, Penang, Malaysia, 27–30 August 2001; p. 1166. [Google Scholar]
- Halls, A.S.; Paxton, B.R.; Hall, N.; Peng Bun, N.; Lieng, S.; Pengby, N.; So, N. The Stationary Trawl (Dai) Fishery of the Tonle Sap-Great Lake, Cambodia; Mekong River Commission: Vientiane, Laos, 2013; No. 32. [Google Scholar]
- Sabo, J.L.; Ruhi, A.; Holtgrieve, G.W.; Elliott, V.; Arias, M.E.; Ngor, P.B.; Räsänen, T.A.; Nam, S. Designing river flows to improve food security futures in the Lower Mekong Basin. Science 2017, 358, eaao1053. [Google Scholar] [CrossRef] [Green Version]
- Phen, C.; Nam, S. Assessment of Gillnets and Other Fishing Gear Used in the Mekong River between Kratie and the Lao PDR Border; Mekong River Commission: Phnom Penh, Cambodia, 2011. [Google Scholar]
- Millar, R.B.; Holst, R. Estimation of gillnet and hook selectivity using log-linear models. ICES J. Mar. Sci. 1997, 54, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Carol, J.; García-Berthou, E. Gillnet selectivity and its relationship with body shape for eight freshwater fish species. J. Appl. Ichthyol. 2007, 23, 654–660. [Google Scholar] [CrossRef]
- Lele, S.R.; Merrill, E.H.; Keim, J.; Boyce, M.S. Selection, use, choice and occupancy: Clarifying concepts in resource selection studies. J. Anim. Ecol. 2013, 82, 1183–1191. [Google Scholar] [CrossRef]
- Boyce, M.S.; McDonald, L.L. Relating populations to habitats using resource selection functions. Trends Ecol. Evol. 1999, 14, 268–272. [Google Scholar] [CrossRef]
- Morato, R.G.; Connette, G.M.; Stabach, J.A.; De Paula, R.C.; Ferraz, K.M.P.M.; Kantek, D.L.Z.; Miyazaki, S.S.; Pereira, T.D.C.; Silva, L.C.; Paviolo, A.; et al. Resource selection in an apex predator and variation in response to local landscape characteristics. Biol. Conserv. 2018, 228, 233–240. [Google Scholar] [CrossRef]
- Freitas, C.; Olsen, E.M.; Knutsen, H.; Albretsen, J.; Moland, E. Temperature-associated habitat selection in a cold-water marine fish. J. Anim. Ecol. 2016, 85, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Nam, S.; Vann, L.S.; Somany, P.; Sinh, L.X.; Robert, P. Assessment of Diversity and Bioecological Characgeristics of Low Value/Small Sized Fish; Aquafish CRSP Project Final Technical Report: Corvallis, OR, USA, 2009. [Google Scholar]
- Kolding, J.; Van Zwieten, P.A.M. Sustainable fishing of inland waters. J. Limnol. 2014, 73, 132–148. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; García Molinos, J.; Zhang, H.; Zhang, M.; Xu, J. Contemporary changes in structural dynamics and socioeconomic drivers of inland fishery in China. Sci. Total Environ. 2019, 648, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, A.F.; Hortle, K.G.; Valbo-Jorgensen, J.; Chan, S.; Chhuon, C.K.; Viravong, S.; Bouakhamvongsa, K.; Suntornratana, U.; Yoorong, N.; Nguyen, T.T.; et al. Distribution and Ecology of some Important Riverine Fish Species of the Mekong River Basin; Mekong River Commission: Vientiane, Laos, 2004. [Google Scholar]
- Lluch-Belda, D.; Magallon, F.J.; Schwartzlose, R.A. Large fluctuations in the sardine fishery in the Gulf of California: Possible causes. CalCOFI Rep. 1986, 27, 136–140. [Google Scholar]
- Essington, T.E.; Moriarty, P.E.; Froehlich, H.E.; Hodgson, E.E.; Koehn, L.E.; Oken, K.L.; Siple, M.C.; Stawitz, C.C. Fishing amplifies forage fish population collapses. Proc. Natl. Acad. Sci. USA 2015, 112, 6648–6652. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.E.; Cochrane, T.A.; Piman, T.; Kummu, M.; Caruso, B.S.; Killeen, T.J. Quantifying changes in flooding and habitats in the Tonle Sap Lake (Cambodia) caused by water infrastructure development and climate change in the Mekong Basin. J. Environ. Manag. 2012, 112, 53–66. [Google Scholar] [CrossRef]
- Winemiller, K.O.; McIntyre, P.B.; Castello, L.; Fluet-Chouinard, E.; Giarrizzo, T.; Nam, S.; Baird, I.G.; Darwall, W.; Lujan, N.K.; Harrison, I.; et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 2016, 351, 128–129. [Google Scholar] [CrossRef] [Green Version]
- Soukhaphon, A.; Baird, I.G.; Hogan, Z.S. The impacts of hydropower dams in the Mekong River basin: A review. Water 2021, 13, 265. [Google Scholar] [CrossRef]
- Dudgeon, D. Asian river fishes in the Anthropocene: Threats and conservation challenges in an era of rapid environmental change. J. Fish Biol. 2011, 79, 1487–1524. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.; Ngor, P.B.; Hogan, Z.S.; So, N.; Brosse, S.; Lek, S. Temporal dynamics of fish assemblages as a reflection of policy shift from fishing concession to co-management in one of the world’s largest tropical flood pulse fisheries. Water 2020, 12, 2974. [Google Scholar] [CrossRef]
- Hecht, J.S.; Lacombe, G.; Arias, M.E.; Dang, T.D.; Piman, T. Hydropower dams of the Mekong River basin: A review of their hydrological impacts. J. Hydrol. 2019, 568, 285–300. [Google Scholar] [CrossRef]
- Vu, A.V.; Hortle, K.G.; Nguyen, D.N. Factors driving long term declines in inland fishery yields in the Mekong delta. Water 2021, 13, 1005. [Google Scholar] [CrossRef]
- Nam, S.; Song, S.L. Fisheries Management and Development in Tonle Sap Great Lake, Cambodia; Fisheries Administration: Phnom Penh, Cambodia, 2011. [Google Scholar]
- Ngor, P.B.; Oberdorff, T.; Phen, C.; Baehr, C.; Grenouillet, G.; Lek, S. Fish assemblage responses to flow seasonality and predictability in a tropical flood pulse system. Ecosphere 2018, 9, e02366. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.M.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 October 2020).
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef]
- Welcomme, R.L. Inland Fisheries: Ecology and Management; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of inland waters. BioScience 2005, 55, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Koning, A.A.; Mcintyre, P.B. Grassroots reserves rescue a river food web from cascading impacts of overharvest. Front. Ecol. Environ. 2021, 19, 152–158. [Google Scholar] [CrossRef]
- Harrell, F.E.J.; Dupont, C. Hmisc: Harrell Miscellaneous. R Package Version 4.4-2. 2020. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 15 January 2021).
- Gu, W.; Swihart, R.K. Absent or undetected? Effects of non-detection of species occurrence on wildlife-habitat models. Biol. Conserv. 2004, 116, 195–203. [Google Scholar] [CrossRef]
- Rotherham, D.; Johnson, D.D.; Kesby, C.L.; Grey, C.A. Sampling estuarine fish and invertebrates with a beam trawl provides a different picture of populations and assemblages than multi-mesh gillnets. Fish. Res. 2012, 123–124, 49–55. [Google Scholar] [CrossRef]
- Jurvelius, J.; Kolari, I.; Leskelä, A. Quality and status of fish stocks in lakes: Gillnetting, seining, trawling and hydroacoustics as sampling methods. Hydrobiologia 2011, 660, 29–36. [Google Scholar] [CrossRef]
- McCann, K.S.; Gellner, G.; McMeans, B.C.; Deenik, T.; Holtgrieve, G.; Rooney, N.; Hannah, L.; Cooperman, M.; Nam, S. Food webs and the sustainability of indiscriminate fisheries. Can. J. Fish. Aquat. Sci. 2016, 73, 656–665. [Google Scholar] [CrossRef]
- Halls, A.S.; Hortle, K.G. Flooding is a key driver of the Tonle Sap dai fishery in Cambodia. Sci. Rep. 2021, 11, 3806. [Google Scholar] [CrossRef]
- Hogan, Z.S.; Moyle, P.B.; May, B.; Vander Zanden, M.J.; Baird, I.G. The imperiled giants of the Mekong. Am. Sci. 2004, 92, 228–237. [Google Scholar] [CrossRef]
- Campbell, T.; Pin, K.; Ngor, P.B.; Hogan, Z. Conserving Mekong megafishes: Current status and critical threats in Cambodia. Water 2020, 12, 1820. [Google Scholar] [CrossRef]
- Karakulak, F.S.; Erk, H. Gill net and trammel net selectivity in the northern Aegean Sea, Turkey. Sci. Mar. 2008, 72, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Handford, P.; Bell, G.; Reimchen, T. A Gillnet Fishery Considered as an Experiment in Artificial Selection. J. Fish. Board Can. 1977, 34, 954–961. [Google Scholar] [CrossRef]
- Hard, J.J.; Gross, M.R.; Heino, M.; Hilborn, R.; Kope, R.G.; Law, R.; Reynolds, J.D. Evolutionary consequences of fishing and their implications for salmon. Evol. Appl. 2008, 1, 388–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audzijonyte, A.; Kuparinen, A.; Gorton, R.; Fulton, E.A. Ecological consequences of body size decline in harvested fish species: Positive feedback loops in trophic interactions amplify human impact. Biol. Lett. 2013, 9, 20121103. [Google Scholar] [CrossRef] [PubMed]
- Fisheries Administration, Ministry of Agriculture, Forestry and Fisheries. FiA Law on Fisheries; Fisheries Administration, Ministry of Agriculture, Forestry and Fisheries: Phnom Phen, Cambodia, 2006. [Google Scholar]
- Bieg, C.; Mccann, K.S.; Mcmeans, B.C.; Rooney, N.; Holtgrieve, G.W.; Lek, S.; Bun, N.P.; Kc, K.B.; Fraser, E. Linking humans to food webs: A framework for the classification of global fisheries. Front. Ecol. Environ. 2018, 16, 412–420. [Google Scholar] [CrossRef]
- Loeung, D. The Bagnet (Dai) Fishery in the Tonle Sap River. In Freshwater Fisheries of Cambodia; Department of Fisheries: Phnom Phen, Cambodia, 1995; pp. 135–149. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelson, S.J.; Hogan, Z.; Jerde, C.L.; Chandra, S.; Ngor, P.B.; Koning, A. Fishing Methods Matter: Comparing the Community and Trait Composition of the Dai (Bagnet) and Gillnet Fisheries in the Tonle Sap River in Southeast Asia. Water 2021, 13, 1904. https://doi.org/10.3390/w13141904
Kelson SJ, Hogan Z, Jerde CL, Chandra S, Ngor PB, Koning A. Fishing Methods Matter: Comparing the Community and Trait Composition of the Dai (Bagnet) and Gillnet Fisheries in the Tonle Sap River in Southeast Asia. Water. 2021; 13(14):1904. https://doi.org/10.3390/w13141904
Chicago/Turabian StyleKelson, Suzanne J., Zeb Hogan, Christopher L. Jerde, Sudeep Chandra, Peng Bun Ngor, and Aaron Koning. 2021. "Fishing Methods Matter: Comparing the Community and Trait Composition of the Dai (Bagnet) and Gillnet Fisheries in the Tonle Sap River in Southeast Asia" Water 13, no. 14: 1904. https://doi.org/10.3390/w13141904
APA StyleKelson, S. J., Hogan, Z., Jerde, C. L., Chandra, S., Ngor, P. B., & Koning, A. (2021). Fishing Methods Matter: Comparing the Community and Trait Composition of the Dai (Bagnet) and Gillnet Fisheries in the Tonle Sap River in Southeast Asia. Water, 13(14), 1904. https://doi.org/10.3390/w13141904