Interpopulation Variability in Dietary Traits of Invasive Bleak Alburnus alburnus (Actinopterygii, Cyprinidae) Across the Iberian Peninsula

 , , ,
, , ,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Field and Laboratory Work
2.3. Data Analyses
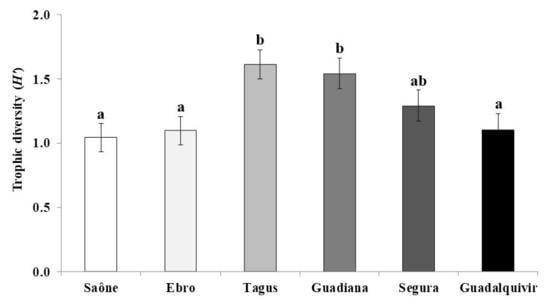
3. Results
4. Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reyjol, Y.; Hugueny, B.; Pont, D.; Bianco, P.G.; Beier, U.; Caiola, N.; Casals, F.; Cowx, I.; Economou, A.; Ferreira, T.; et al. Patterns in species richness and endemism of European freshwater fish. Glob. Ecol. Biogeogr. 2007, 16, 65–75. [Google Scholar] [CrossRef]
- Almeida, D.; Ribeiro, F.; Leunda, P.M.; Vilizzi, L.; Copp, G.H. Effectiveness of FISK, an invasiveness screening tool for non-native freshwater fishes, to perform risk identification assessments in the Iberian Peninsula. Risk Anal. 2013, 33, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.; Leunda, P.M. Non-native fish impacts on Mediterranean freshwater ecosystems: Current knowledge and research needs. Fish. Manag. Ecol. 2012, 19, 142–156. [Google Scholar] [CrossRef]
- Vinni, M.; Horppila, J.; Olin, M.; Ruuhijärvi, J.; Nyberg, K. The food, growth and abundance of five co-existing cyprinids in lake basins of different morphometry and water quality. Aquat. Ecol. 2000, 34, 421–431. [Google Scholar] [CrossRef]
- Vašek, M.; Kubečka, J. In situ diel patterns of zooplankton consumption by subadult/adult roach Rutilus rutilus, bream Abramis brama, and bleak Alburnus alburnus. Folia Zool. 2004, 53, 203–214. [Google Scholar]
- Keith, P.; Persat, H.; Feunteun, E.; Allardi, J. Les Poissons d’Eau Douce de France; Biotope Éditions Muséum National d’Histoire Naturelle: Paris, France, 2011; ISBN 9782914817691. [Google Scholar]
- Vinyoles, D.; Robalo, J.I.; De Sostoa, A.; Almodóvar, A.; Elvira, B.; Nicola, G.G.; Fernández-Delgado, C.; Santos, C.S.; Doadrio, I.; Sardà-Palomera, F.; et al. Spread of the alien bleak Alburnus alburnus (Linnaeus, 1758) (Actinopterygii, Cyprinidae) in the Iberian Peninsula: The role of reservoirs. Graellsia 2007, 63, 101–110. [Google Scholar] [CrossRef]
- Masó, G.; Latorre, D.; Tarkan, A.S.; Vila-Gispert, A.; Almeida, D. Interpopulation plasticity in growth and reproduction of invasive bleak, Alburnus alburnus (Cyprinidae, Actinopterygii), in northeastern Iberian Peninsula. Folia Zool. 2016, 65, 10–14. [Google Scholar] [CrossRef]
- Matono, P.; Da Silva, J.; Ilhéu, M. How Does an invasive cyprinid benefit from the hydrological disturbance of Mediterranean temporary streams? Diversity 2018, 10, 47. [Google Scholar] [CrossRef]
- Amat-Trigo, F.; Torralva, M.; Ruiz-Navarro, A.; Oliva-Paterna, F.J. Colonization and plasticity in population traits of the invasive Alburnus alburnus along a longitudinal river gradient in a Mediterranean river basin. Aquat. Invasions 2019, 14, 310–331. [Google Scholar] [CrossRef]
- Almeida, D.; Grossman, G.D. Utility of direct observational methods for assessing competitive interactions between non-native and native freshwater fishes. Fish. Manag. Ecol. 2012, 19, 157–166. [Google Scholar] [CrossRef]
- Vilizzi, L.; Copp, G.H.; Adamovich, B.; Almeida, D.; Chan, J.; Davison, P.I.; Dembski, S.; Ekmekçi, F.G.; Ferincz, Á.; Forneck, S.C.; et al. A global review and meta-analysis of applications of the freshwater Fish Invasiveness Screening Kit. Rev. Fish Biol. Fish. 2019, 29, 529–568. [Google Scholar] [CrossRef]
- Latorre, D.; Masó, G.; Hinckley, A.; Rubio-Gracia, F.; Vila-Gispert, A.; Almeida, D. Interpopulation plasticity in dietary traits of invasive bleak Alburnus alburnus (Linnaeus, 1758) in Iberian fresh waters. J. Appl. Ichthyol. 2016, 32, 1252–1255. [Google Scholar] [CrossRef]
- Almeida, D.; Fletcher, D.H.; Rangel, C.; García-Berthou, E.; Da Silva, E. Dietary traits of invasive bleak Alburnus alburnus (Actinopterygii, Cyprinidae) between contrasting habitats in Iberian fresh waters. Hydrobiologia 2017, 795, 23–33. [Google Scholar] [CrossRef]
- Copp, G.H.; Bianco, P.G.; Bogutskaya, N.G.; Eros, T.; Falka, I.; Ferreira, M.T.; Fox, M.G.; Freyhof, J.; Gozlan, R.E.; Grabowska, J.; et al. To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 2005, 21, 242–262. [Google Scholar] [CrossRef]
- Almeida, D.; Almodóvar, A.; Nicola, G.G.; Elvira, B.; Grossman, G.D. Trophic plasticity of invasive juvenile largemouth bass Micropterus salmoides in Iberian streams. Fish. Res. 2012, 113, 153–158. [Google Scholar] [CrossRef]
- Clavero, M.; García-Berthou, E. Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol. Appl. 2006, 16, 2313–2324. [Google Scholar] [CrossRef]
- Závorka, L.; Buoro, M.; Cucherousset, J. The negative ecological impacts of a globally introduced species decrease with time since introduction. Glob. Chang. Biol. 2018, 24, 4428–4437. [Google Scholar] [CrossRef]
- Ministry of Environment France. Available online: http://vigiprevi.meteofrance.com/PREV/V/index.html (accessed on 9 June 2020).
- Ministry of Environment Spain. Available online: http://www.aemet.es/es/serviciosclimaticos (accessed on 9 June 2020).
- European Commission. Commission Directive 2014/101/EU of 30 October 2014 amending Directive 2000/60/EC of the European Parliament and of the Council establishing a framework for Community action in the field of water policy. Off. J. Eur. Union-Legis. 2014, 311, 32–35. [Google Scholar]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’Eau Douce: Systématique, Biologie, Écologie; CNRS Éditions: Paris, France, 2010; ISBN 9782271069450. [Google Scholar]
- Alcaraz, C.; García-Berthou, E. Food of an endangered cyprinodont (Aphanius iberus): Ontogenetic diet shift and prey electivity. Environ. Biol. Fishes 2007, 78, 193–207. [Google Scholar] [CrossRef]
- Johnson, P.C.D.; Barry, S.J.E.; Ferguson, H.M.; Müller, P. Power analysis for generalized linear mixed models in ecology and evolution. Methods Ecol. Evol. 2015, 6, 133–142. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.r-project.org (accessed on 1 October 2019).
- Latorre, D.; Masó, G.; Hinckley, A.; Verdiell-Cubedo, D.; Tarkan, A.S.; Vila-Gispert, A.; Copp, G.H.; Cucherousset, J.; Da Silva, E.; Fernández-Delgado, C.; et al. Interpopulation variability in growth and reproduction of invasive bleak Alburnus alburnus (Linnaeus, 1758) across the Iberian Peninsula. Mar. Freshw. Res. 2018, 69, 1326–1332. [Google Scholar] [CrossRef]
- Bøhn, T.; Terje Sandlund, O.; Amundsen, P.-A.; Primicerio, R. Rapidly changing life history during invasion. Oikos 2004, 106, 138–150. [Google Scholar] [CrossRef]
- Oksanen, J.; Minchin, P.R. Continuum theory revisited: What shape are species responses along ecological gradients? Ecol. Model. 2002, 157, 119–129. [Google Scholar] [CrossRef]
- Gasith, A.; Resh, V.H. Streams in Mediterranean climate regions: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- Almeida, D.; Almodóvar, A.; Nicola, G.G.; Elvira, B. Feeding tactics and body condition of two introduced populations of pumpkinseed Lepomis gibbosus: Taking advantages of human disturbances? Ecol. Freshw. Fish 2009, 18, 15–23. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| River: | Saône | Ebro | Tagus | Guadiana | Segura | Guadalquivir | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Food Category | Oc. | Mass | Oc. | Mass | Oc. | Mass | Oc. | Mass | Oc. | Mass | Oc. | Mass |
| Algae and plant debris | 3 | <1 | 2 | 5 | 29 | 17 | 5 | 3 | 3 | <1 | 8 | 9 |
| Zooplankton a | 50 | 17 | 40 | 15 | 11 | 8 | 1 | <1 | 5 | 3 | 10 | 6 |
| Ephemeroptera and Plecoptera nymphs | 5 | 9 | 33 | 21 | 5 | 10 | 20 | 23 | 7 | 21 | 5 | 12 |
| Odonata nymphs | 1 | 2 | 11 | 6 | 1 | 5 | 5 | 9 | 1 | <1 | 1 | 3 |
| Diptera larvae | 69 | 39 | 27 | 15 | 3 | 15 | 3 | 1 | 59 | 31 | 3 | 16 |
| Trichoptera larvae | 6 | 1 | 5 | <1 | 1 | <1 | 15 | 13 | 6 | 11 | 1 | 5 |
| Other benthic invertebrates b | 12 | 4 | 3 | 3 | 53 | 13 | 1 | <1 | 12 | 1 | 71 | 14 |
| Nektonic and neustonic insects c | 1 | <1 | 1 | <1 | 1 | <1 | 1 | <1 | 1 | <1 | 20 | 19 |
| Flying insects d | 3 | <1 | 5 | 8 | 35 | 7 | 61 | 23 | 3 | 19 | – | – |
| Terrestrial arthropods e | 1 | <1 | 1 | <1 | – | – | 1 | 9 | 1 | <1 | 7 | 2 |
| Detritus | 31 | 27 | 25 | 26 | 42 | 24 | 24 | 18 | 31 | 13 | 21 | 14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latorre, D.; Masó, G.; Hinckley, A.; Verdiell-Cubedo, D.; Castillo-García, G.; González-Rojas, A.G.; Black-Barbour, E.N.; Vila-Gispert, A.; García-Berthou, E.; Miranda, R.; et al. Interpopulation Variability in Dietary Traits of Invasive Bleak Alburnus alburnus (Actinopterygii, Cyprinidae) Across the Iberian Peninsula. Water 2020, 12, 2200. https://doi.org/10.3390/w12082200
Latorre D, Masó G, Hinckley A, Verdiell-Cubedo D, Castillo-García G, González-Rojas AG, Black-Barbour EN, Vila-Gispert A, García-Berthou E, Miranda R, et al. Interpopulation Variability in Dietary Traits of Invasive Bleak Alburnus alburnus (Actinopterygii, Cyprinidae) Across the Iberian Peninsula. Water. 2020; 12(8):2200. https://doi.org/10.3390/w12082200
Chicago/Turabian StyleLatorre, Dani, Guillem Masó, Arlo Hinckley, David Verdiell-Cubedo, Gema Castillo-García, Anni G. González-Rojas, Erin N. Black-Barbour, Anna Vila-Gispert, Emili García-Berthou, Rafael Miranda, and et al. 2020. "Interpopulation Variability in Dietary Traits of Invasive Bleak Alburnus alburnus (Actinopterygii, Cyprinidae) Across the Iberian Peninsula" Water 12, no. 8: 2200. https://doi.org/10.3390/w12082200
APA StyleLatorre, D., Masó, G., Hinckley, A., Verdiell-Cubedo, D., Castillo-García, G., González-Rojas, A. G., Black-Barbour, E. N., Vila-Gispert, A., García-Berthou, E., Miranda, R., Oliva-Paterna, F. J., Ruiz-Navarro, A., da Silva, E., Fernández-Delgado, C., Cucherousset, J., Serrano, J. M., & Almeida, D. (2020). Interpopulation Variability in Dietary Traits of Invasive Bleak Alburnus alburnus (Actinopterygii, Cyprinidae) Across the Iberian Peninsula. Water, 12(8), 2200. https://doi.org/10.3390/w12082200