Effect of Land Use on the Benthic Diatom Community of the Danube River in the Region of Budapest

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
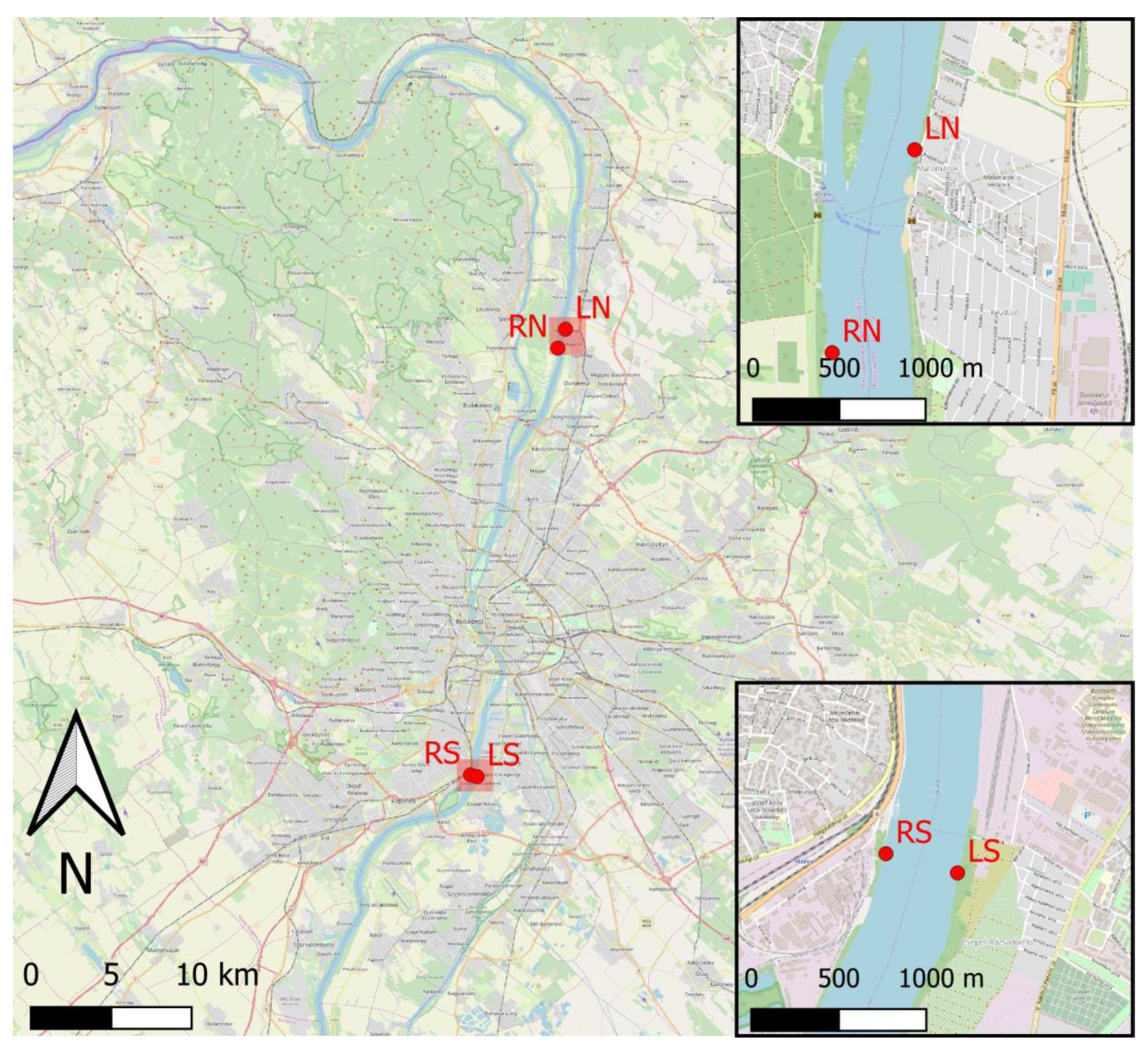
2.1. Study Site
2.2. Sampling
2.3. Environmental Variables
2.4. Diatom Sample Processing
2.5. Diatom Index and Trait Data
2.6. Data Analysis
3. Results
3.1. Environmental Variables
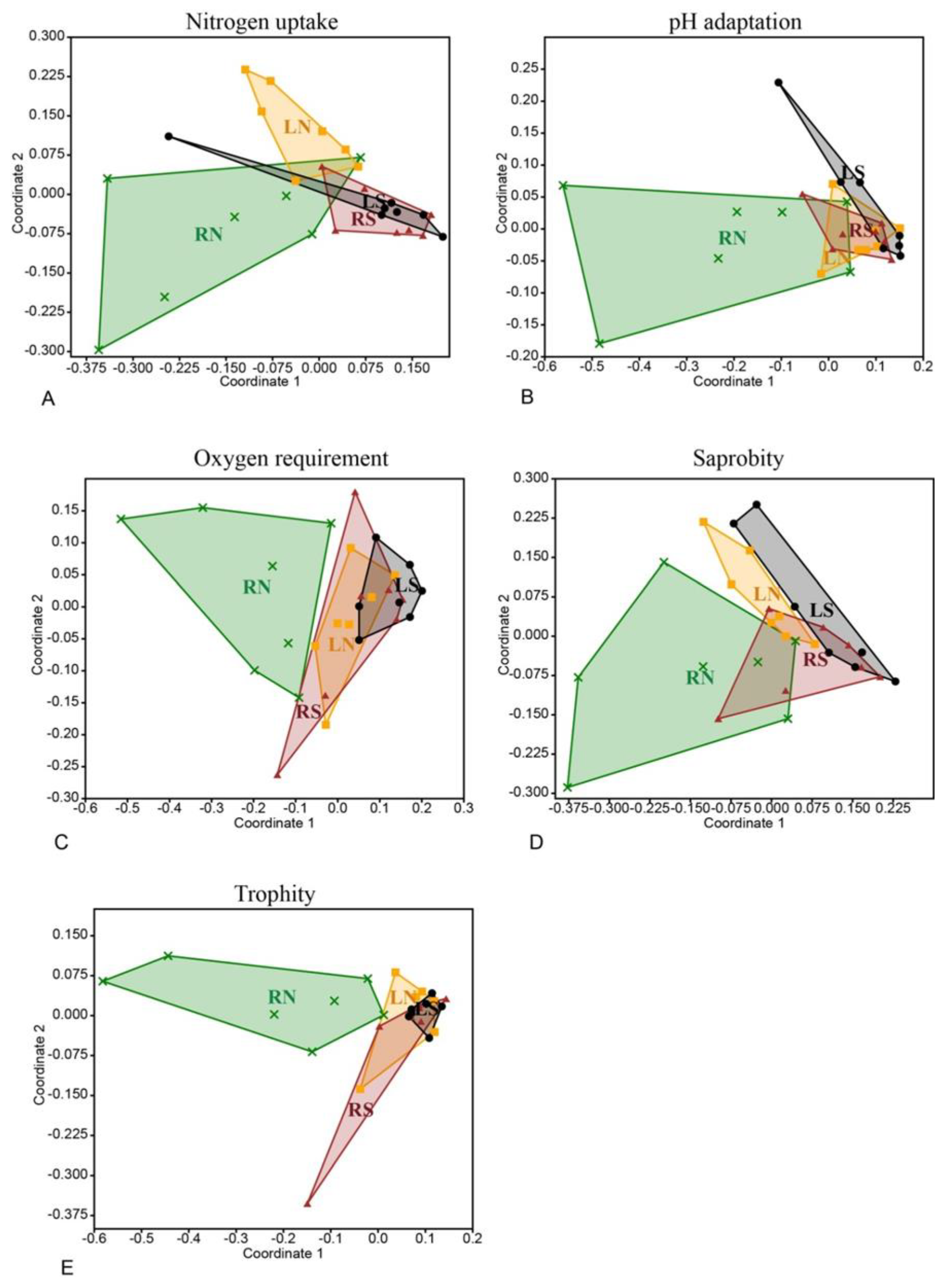
3.2. Diatom Assemblages
3.2.1. Species
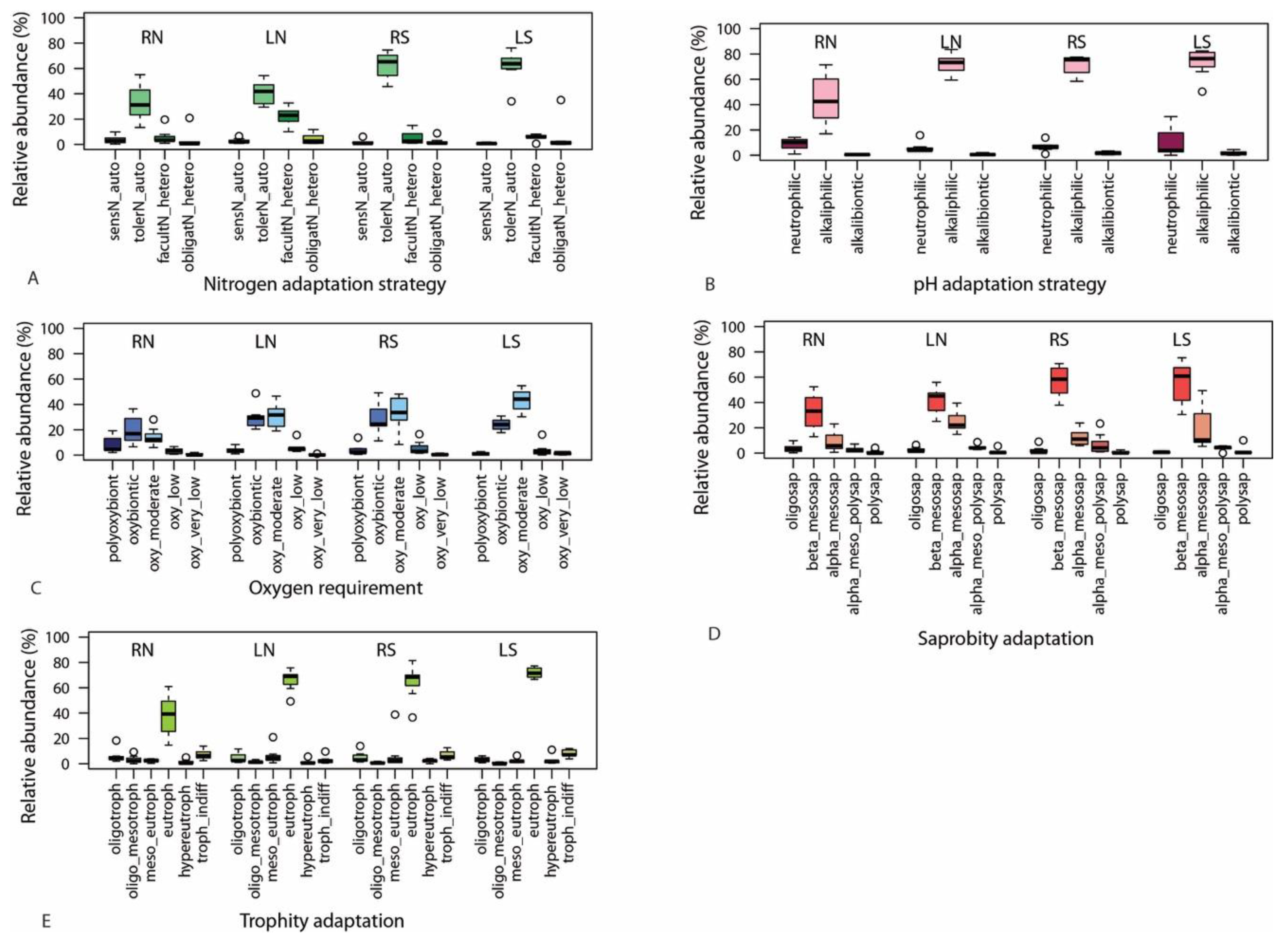
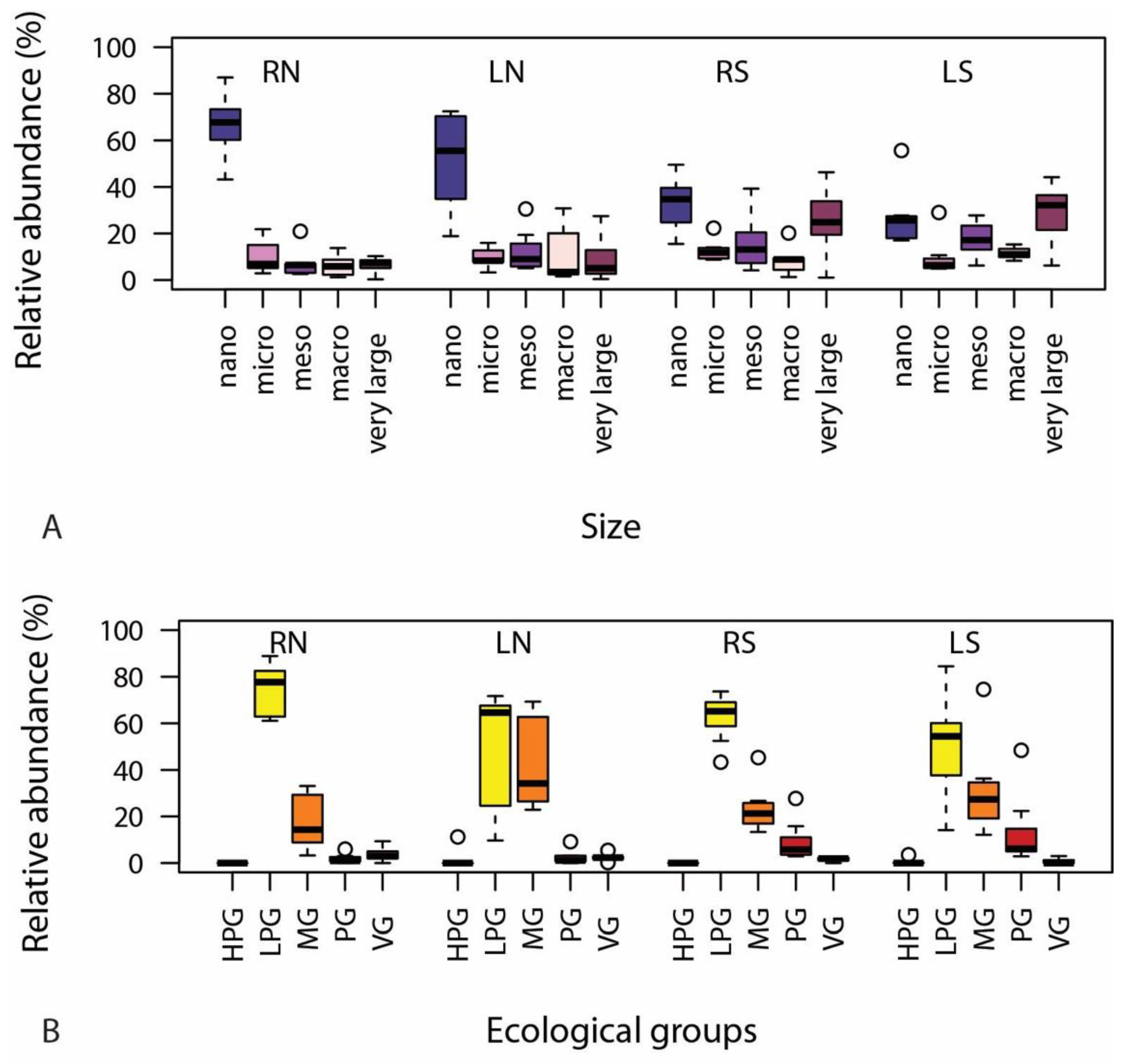
3.2.2. Traits
4. Discussion
4.1. Water Quality Based on Environmental Variables
4.2. Ecological Status Indicated by Diatom Assemblages and the Usability of Traits
4.2.1. Species Composition
4.2.2. Traits
4.2.3. Diatom Index
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paul, M.J.; Meyer, J.L. Streams in the urban landscape. Annu. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. North Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Meyer, J.L.; Paul, M.J.; Taulbee, W.K. Stream ecosystem function in urbanizing landscapes. J. North Am. Benthol. Soc. 2005, 24, 602–612. [Google Scholar] [CrossRef]
- Alexander, M. Biodegradation and Bioremediation, 2nd ed.; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Douziech, M.; Conesa, I.R.; Benítez-López, A.; Franco, A.; Huijbregts, M.; van Zelm, R. Quantifying variability in removal efficiencies of chemicals in activated sludge wastewater treatment plants—A meta–analytical approach. Environ. Sci. Process. Impacts 2018, 24, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Weiwei, B.; Zhu, B.; Yuan, X.; Zhang, Y.; Yang, M.; Qiang, Z. Occurrence, removal and risk of organic micropollutants in wastewater treatment plants across China: Comparison of wastewater treatment processes. Water Res. 2018, 130, 38–46. [Google Scholar] [CrossRef]
- Ntengwe, F.W. Pollutant loads and water quality in streams of heavily populated and industrialised towns. Phys. Chem. Earth 2006, 31, 832–839. [Google Scholar] [CrossRef]
- Szczepocka, E.; Szulc, B. The use of benthic diatoms in estimating water quality of variously polluted rivers. Oceanol. Hydrobiol. Stud. 2009, 38, 17–26. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera, 1st ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Burliga, A.L.; Kociolek, J.P. Diatoms (Bacillariophyta) in Rivers. In River Algae, 1st ed.; Necchi, O., Jr., Ed.; Springer Nature: Cham, Switzerland, 2016; pp. 73–128. [Google Scholar] [CrossRef]
- Mann, D.G.; Vanormelingen, P. An inordinate fondness? The number, distributions, and origins of diatom species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef]
- Lobo, E.A.; Heinrich, C.G.; Schuch, M.; Wetzel, C.E.; Ector, L. Diatoms as Bioindicators in Rivers. In River Algae, 1st ed.; Necchi, O., Jr., Ed.; Springer Nature: Cham, Switzerland, 2016; pp. 245–272. [Google Scholar] [CrossRef]
- Kelly, M.G.; King, L.; Jones, R.I.; Barker, P.A.; Jamieson, B.J. Validation of diatoms as proxies for phytobenthos when assessing ecological status in lakes. Hydrobiologia 2008, 610, 125–129. [Google Scholar] [CrossRef]
- Ector, L.; Kingston, J.C.; Charles, D.F.; Denys, L.; Douglas, M.S.V.; Manoylov, K.; Michelutti, N.; Rimet, F.; Smol, J.P.; Stevenson, R.J.; et al. Freshwater diatoms and their role as ecological indicators. In Proceedings of the 17th International Diatom Symposium, Ottawa, ON, Canada, 25–31 August 2002; pp. 469–480. [Google Scholar]
- European Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23rd October 2000 establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 2000, 327, 1–72. [Google Scholar]
- Berthon, V.; Bouchez, A.; Rimet, F. Using diatom life-forms and ecological guilds to assess organic pollution and trophic level in rivers: A case study of rivers in south-eastern France. Hydrobiologia 2011, 673, 259–271. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Tapolczai, K.; Bouchez, A.; Stenger-Kovács, C.S.; Padisák, J.; Rimet, F. Trait-based ecological classifications for benthic algae: Review and perspectives. Hydrobiologia 2016, 776, 1–17. [Google Scholar] [CrossRef]
- Passy, S.I. Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquat. Bot. 2007, 86, 171–178. [Google Scholar] [CrossRef]
- Rimet, F.; Bouchez, A. Life-forms, cell-sizes and ecological guilds of diatoms in European rivers. Knowl. Manag. Aquat. Ecosyst. 2012, 406, 1. [Google Scholar] [CrossRef]
- van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Alygizakis, N.A.; Besselink, H.; Paulus, G.K.; Oswald, P.; Hornsta, L.M.; Oswaldova, M.; Medema, G.; Thomaidis, N.S.; Behnisch, P.A.; Slobodnik, J. Characterization of wastewater effluents in the Danube River Basin with chemical screening, in vitro bioassays and antibiotic resistant genes analysis. Environ. Int. 2019, 127, 420–429. [Google Scholar] [CrossRef]
- International Commission for the Protection of the Danube River (ICPDR), Joint Danube Survey. 2019. Available online: http://www.icpdr.org/main/activities–projects/joint–danube–survey (accessed on 17 October 2019).
- Hungarian Central Statistical Office. Gazetteer of Hungary 1 January 2018; Hungarian Central Statistical Office: Budapest, Hungary, 2018; pp. 1–235. [Google Scholar]
- Detailed Gazetteer of Hungary, Dunakeszi. 2019. Available online: http://www.ksh.hu/apps/hntr.telepules?p_lang=HU&p_id=18616 (accessed on 17 October 2019).
- Tatai, Z.S.; Zétényi, D.; Niedetzky, A.; Szőke, B.; Bódi-Nagy, A.; Pogány, A.; Orosz, I.; Szabó, K.; Becsák, P. Budapest Környezeti Állapotértékelése 2017; Budapest Főváros Önkormányzatának Főpolgármesteri Hivatala Városigazgatóság Főosztály: Budapest, Hungary, 2017; pp. 1–259. [Google Scholar]
- Schwarz, U.; Holubova, K.; Cuban, R.; Matok, P.; Busovsky, J. Joint Danube Survey 3—Extended Report on: Hydromorphology; International Commission for the Protection of the Danube River: Vienna, Austria, 2014. [Google Scholar]
- Belletti, B.; Rinaldi, M.; Buijse, A.D.; Gurnell, A.M.; Mosselman, E. A review of assessment methods for river hydromorphology. Environ. Earth Sci. 2015, 73, 2079–2100. [Google Scholar] [CrossRef]
- European Committee for Standardisation. EN 13946 Water Quality. Guidance Standard for the Routine Sampling and Pretreatment of Benthic Diatoms from Rivers; European Committee for Standardisation: Brussels, Belgium, 2002; pp. 1–18. [Google Scholar]
- Eaton, A.D.; Clesceri, L.S.; Rice, E.W.; Greenberg, A.E. Standard Methods for the Examination of Water and Wastewater, 21th ed.; APHA; AWWA; WEF: Washington, DC, USA, 2005. [Google Scholar]
- Hungarian Hydrological Forecasting Service. 2019. Available online: www.hydroinfo.hu (accessed on 17 October 2019).
- Cemagref. Etude des Méthodes Biologiques Quantitativesd’appréciation de La Qualité des Eaux; Rapport Division Qualité des Eaux Lyon, Agence Financière de Bassin Rhône-Méditerranée-Corse: Pierre-Bénite, France, 1982; pp. 1–218. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Lange, K.; Townsend, C.R.; Matthae, C.D. A trait–based framework for stream algal communities. Ecol. Evol. 2016, 6, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System, Open Source Geospatial Foundation Project; Open Source Geospatial Foundation: Chicago, IL, USA, 2019. [Google Scholar]
- Gurnell, A.; Lee, M.; Souch, C. Urban Rivers: Hydrology, Geomorphology, Ecology and Opportunities for Change. Geogr. Compass 2007, 1–5, 1118–1137. [Google Scholar] [CrossRef]
- Hamchevici, C.; Dumitrache, F.; Sena, F.; Umlauf, G.; Postolache, C.; Udrea, I. Joint Danube Survey 3—Chapter (Full Report) on: General Physico–Chemical Determinands and Nutrients; International Commission for the Protection of the Danube River: Vienna, Austria, 2014. [Google Scholar]
- Budapest Sewage Works Pte Ltd., Wastewater Treatment. Available online: http://www.fcsm.hu/en/services/wastewater_treatment/wastewater_treatment/ (accessed on 17 October 2019).
- Budapest Waterworks, Route of Wastewater. Available online: https://www.vizmuvek.hu/en/company_information/water_supply_wastewater_drainage_and_treatment/wastewater_drainage_treatment/technology/route_of_wastewater (accessed on 17 October 2019).
- Budapest Sewage Works Pte Ltd., Technical Data. 2019. Available online: http://www.fcsm.hu/en/company_information/data/technical_data/ (accessed on 17 October 2019).
- Kovács, J.; Kovács, S.; Hatvani, I.G.; Magyar, N.; Tanos, P.; Korponai, J.; Blaschke, A.P. Spatial optimization of monitoring networks on the examples of a river, a lake-wetland system and a sub-surface water system. Water Resour. Manag. 2015, 29, 5275–5294. [Google Scholar] [CrossRef]
- Bolong, N.; Ismail, A.; Salim, M.; Matsuura, T. A review of the effects of emerging contaminants in wastewater and options for their removal. Desalination 2009, 239, 229–246. [Google Scholar] [CrossRef]
- Makovinská, J.; Hlúbiková, D.; Fidlerová, D. Joint Danube Survey 3—Phytobenthos; International Commission for the Protection of the Danube River: Vienna, Austria, 2014. [Google Scholar]
- Peterson, D.A.; Porter, S.D. Biological and Chemical Indicators of Eutrophication in the Yellowstone River and Major Tributaries during August 2000; National Water Monitoring Council: Washington, DC, USA, 2002; pp. 1–14. [Google Scholar]
- Carayon, D.; Tison-Rosebery, J.; Delmas, F. Defining a new autoecological trait matrix for French stream benthic diatoms. Ecol. Indic. 2019, 103, 650–658. [Google Scholar] [CrossRef]
- Pérès, F.; Barthès, A.; Ponton, E.; Coste, M.; Ten-Hague, L.; Le–Cohu, R. Achnanthidium delmontii sp. nov.; a new species from French rivers. Fottea 2012, 12, 189–198. [Google Scholar] [CrossRef]
- Falasco, E.; Piano, E.; Bona, F. Diatom flora in Mediterranean streams: Flow intermittency threatens endangered species. Biodivers. Conserv. 2016, 25, 2965–2986. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Cantonati, M., Kelly, M.G., Lange–Bertalot, H., Eds.; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1. In Teil: Naviculaceae. Süßwasserflora von Mitteleuropa 2/1, 1st ed.; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustva Fischer Verlag: Jena, Germany, 1986; pp. 1–876. [Google Scholar]
- Ács, E.; Kiss, K.T. Colonization processes of diatoms on artificial substrates in the River Danube near Budapest (Hungary). Hydrobiologia 1993, 269–270, 307–315. [Google Scholar] [CrossRef]
- Trábert, Z.S.; Kiss, K.T.; Várbíró, G.; Dobosy, P.; Grigorszky, I.; Ács, É. Comparison of the utility of a frequently used diatom index (IPS) and the diatom ecological guilds in the ecological status assessment of large rivers. Fundam. Appl. Limnol. Arch. Hydrobiol. 2017, 189, 87–103. [Google Scholar] [CrossRef]
- Bahls, L.L. Periphyton Bioassessment Methods for Montana Streams; Water Quality Bureau, Department of Health and Environmental Sciences: Helena, MT, USA, 1993; pp. 1–132. [Google Scholar]
- Hambrook Berkman, J.A.; Porter, S.D. An overview of algal monitoring and research in the U.S. Geological Survey’s National Water Quality Assessment (NAWQA) Program. Diatom 2004, 20, 13–22. [Google Scholar] [CrossRef]
- Lange, K.; Liess, A.; Piggott, J.J.; Townsend, C.R.; Matthaei, C.D. Light, nutrients and grazing interact to determine stream diatom community composition and functional group structure. Freshw. Biol. 2011, 56, 264–278. [Google Scholar] [CrossRef]
- Ács, E.; Kiss, K.T. Effects of the water discharge on periphyton abundance and diversity in a large river (River Danube, Hungary). Hydrobiologia 1993, 249, 125–133. [Google Scholar] [CrossRef]
- Ács, E.; Kiss, K.T.; Szabó, K.; Makk, J. Short-term colonization sequence of periphyton on glass slides in a large river (River Danube, near Budapest). Acta Bot. Croat. 2000, 100, 135–156. [Google Scholar] [CrossRef]
- Ács, E.; Szabó, K.; Kiss, K.T.; Hindák, F. Benthic algal investigations in the Danube river and some of its main tributaries from Germany to Hungary. Biologia 2003, 58, 545–554. [Google Scholar]
- Litchman, E.; de Tezanos Pinto, P.; Klausmeier, C.A.; Thomas, M.K.; Yoshiyama, K. Linking traits to species diversity and community structure in phytoplankton. Hydrobiologia 2010, 653, 15–28. [Google Scholar] [CrossRef]
- Wynn, K.H.; Bauch, N.J.; Driver, N.E. Gore Creek Watershed, Colorado—Assessment of Historical and Current Water Quantity, Water Quality, and Aquatic Ecology, 1968–1998; US Geological Survey: Denver, CO, USA, 2001; pp. 1–72.
- Ács, E.; Borics, G.; Kiss, K.T.; Várbíró, G. Módszertani Útmutató a Fitobentosz Élőlénycsoport VKI Szerinti Gyűjtéséhez, Feldolgozásához és Kiértékeléséhez. Kézirat; MTA Ökológiai Kutatóközpont: Tihany, Magyarország, 2015; pp. 1–69. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Point | Vegetation Cover | Bank Modification | Land Cover in 500 m Buffer | Sum |
|---|---|---|---|---|
| Upstream right bank | 1 | 1 | 1 | 3 |
| Upstream left bank | 2 | 2 | 2 | 6 |
| Downstream right bank | 3 | 3 | 3 | 9 |
| Downstream left bank | 2 | 2 | 3 | 7 |
| OMNIDIA Code | Average Dissimilarity | Contribution % | Cumulative % | Mean RN | Mean LN | Mean LS | Mean RS | |
|---|---|---|---|---|---|---|---|---|
| Achnanthidium delmontii Pérès, Le Cohu & Barthès | ADMO | 9.278 | 14.28 | 14.28 | 33.8 | 7.56 | 1.26 | 5.62 |
| Cocconeis euglypta Ehrenberg | CEUG | 8.342 | 12.84 | 27.12 | 6.02 | 7.12 | 24.5 | 24.7 |
| Amphora pediculus (Kützing) Grunow | APED | 5.854 | 9.013 | 36.14 | 17.1 | 20.8 | 13.8 | 11.8 |
| Nitzschia inconspicua Grunow | NINC | 4.11 | 6.327 | 42.46 | 3.51 | 15.4 | 1.66 | 0.861 |
| Discostella pseudostelligera (Hustedt) Houk & Klee | DPSG | 2.426 | 3.734 | 46.2 | 0.833 | 1.33 | 6.4 | 3.91 |
| Navicula cryptotenella Lange-Bertalot | NCTE | 2.1 | 3.232 | 49.43 | 2.19 | 2.08 | 7.14 | 4.42 |
| Amphora inariensis Krammer | AINA | 1.963 | 3.022 | 52.45 | 6.03 | 4.77 | 3.85 | 4.33 |
| Nitzschia paleacea (Grunow) Grunow | NPAE | 1.731 | 2.665 | 55.12 | 2.62 | 0.829 | 3.18 | 1.13 |
| Navicula tripunctata (O.F. Müller) Bory | NTPT | 1.58 | 2.432 | 57.55 | 0.224 | 1.25 | 4.69 | 2.64 |
| Navicula antonii Lange-Bertalot | NANT | 1.48 | 2.278 | 59.83 | 2.34 | 3.29 | 3.18 | 1.75 |
| Ecological Status Boundary | IPS Value |
|---|---|
| High/good | 15.6 |
| Good/moderate | 13.0 |
| Moderate/poor | 8.7 |
| poor/bad | 4.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trábert, Z.; Duleba, M.; Bíró, T.; Dobosy, P.; Földi, A.; Hidas, A.; Kiss, K.T.; Óvári, M.; Takács, A.; Várbíró, G.; et al. Effect of Land Use on the Benthic Diatom Community of the Danube River in the Region of Budapest. Water 2020, 12, 479. https://doi.org/10.3390/w12020479
Trábert Z, Duleba M, Bíró T, Dobosy P, Földi A, Hidas A, Kiss KT, Óvári M, Takács A, Várbíró G, et al. Effect of Land Use on the Benthic Diatom Community of the Danube River in the Region of Budapest. Water. 2020; 12(2):479. https://doi.org/10.3390/w12020479
Chicago/Turabian StyleTrábert, Zsuzsa, Mónika Duleba, Tibor Bíró, Péter Dobosy, Angéla Földi, András Hidas, Keve Tihamér Kiss, Mihály Óvári, Anita Takács, Gábor Várbíró, and et al. 2020. "Effect of Land Use on the Benthic Diatom Community of the Danube River in the Region of Budapest" Water 12, no. 2: 479. https://doi.org/10.3390/w12020479
APA StyleTrábert, Z., Duleba, M., Bíró, T., Dobosy, P., Földi, A., Hidas, A., Kiss, K. T., Óvári, M., Takács, A., Várbíró, G., Záray, G., & Ács, É. (2020). Effect of Land Use on the Benthic Diatom Community of the Danube River in the Region of Budapest. Water, 12(2), 479. https://doi.org/10.3390/w12020479