Stable Carbon Isotopes of Phytoplankton as a Tool to Monitor Anthropogenic CO2 Submarine Leakages

 ,
,  ,
,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
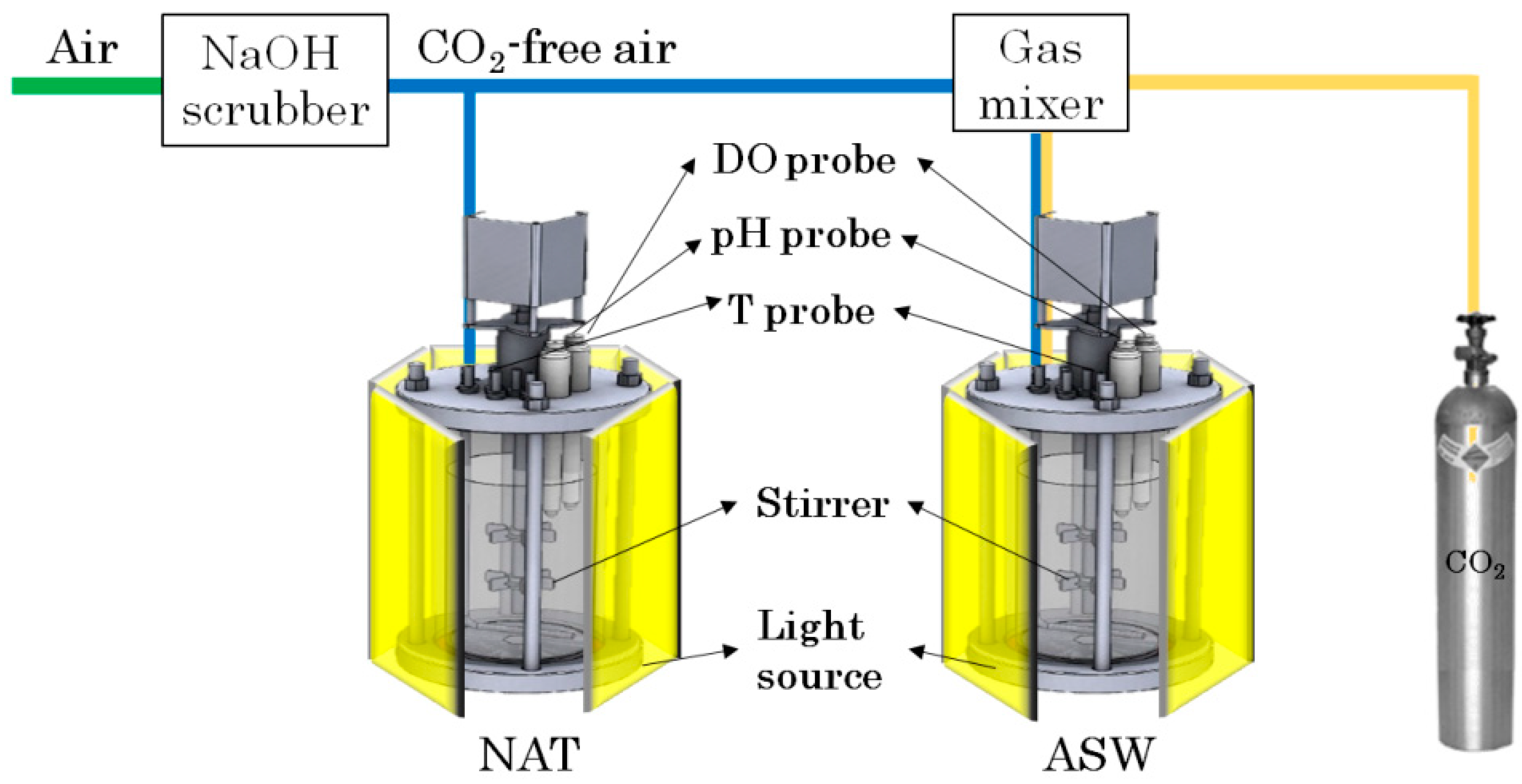
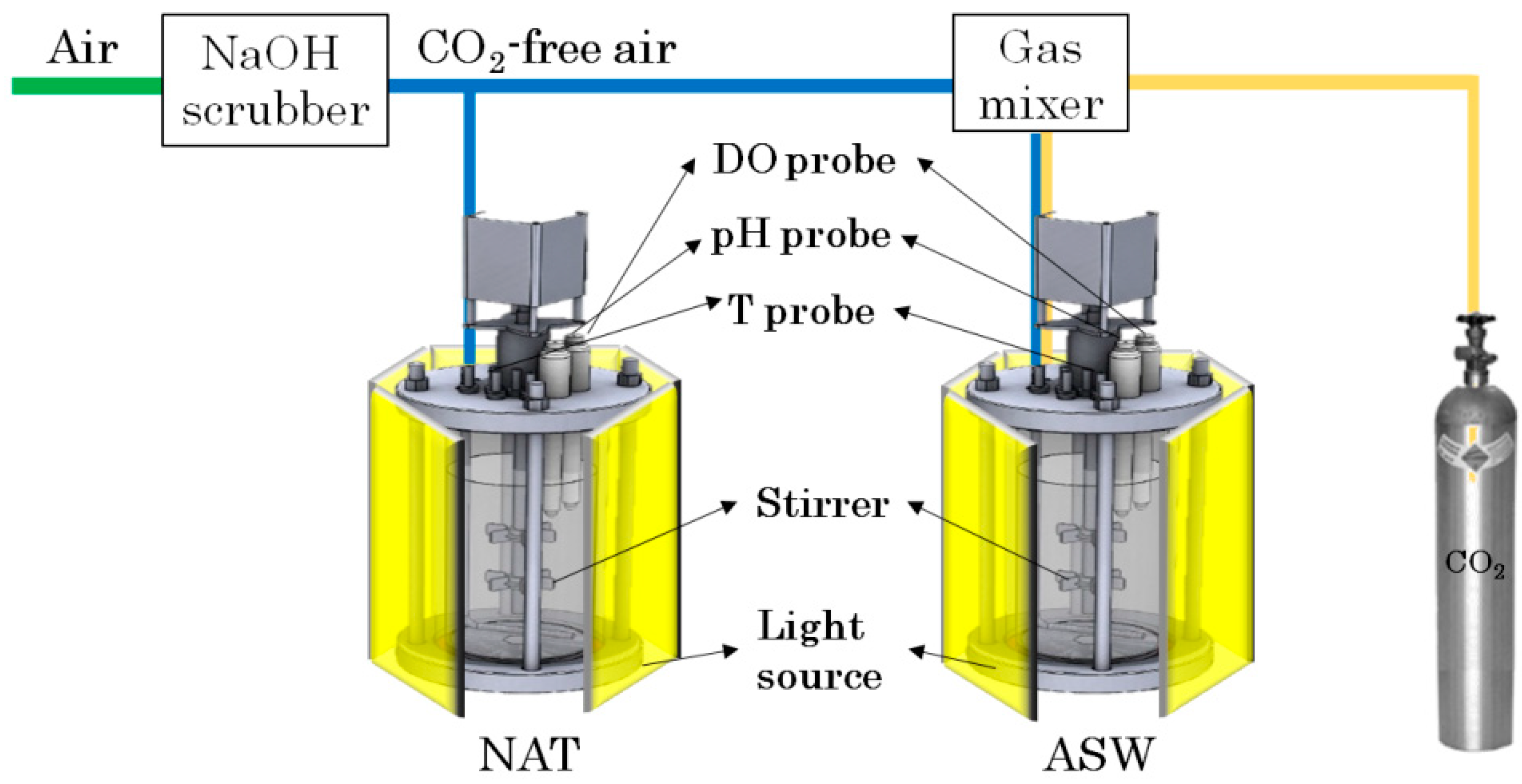
2.1. Experimental Setup
2.2. Sampling and Analysis for Phytoplankton Abundance and Media Characterisation
2.3. Sampling and Analysis of Medium and Phytoplankton Carbon Stable Isotopic Composition and Calculation of Isotopic Fractionation
2.4. Statistical Analysis
3. Results
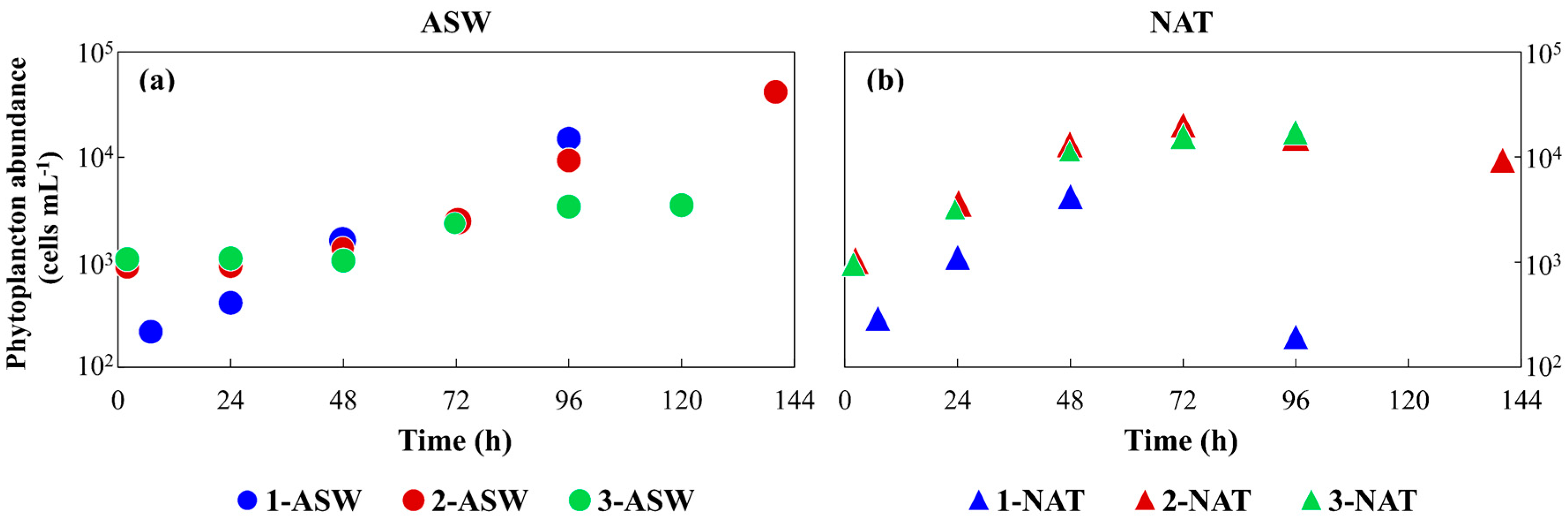
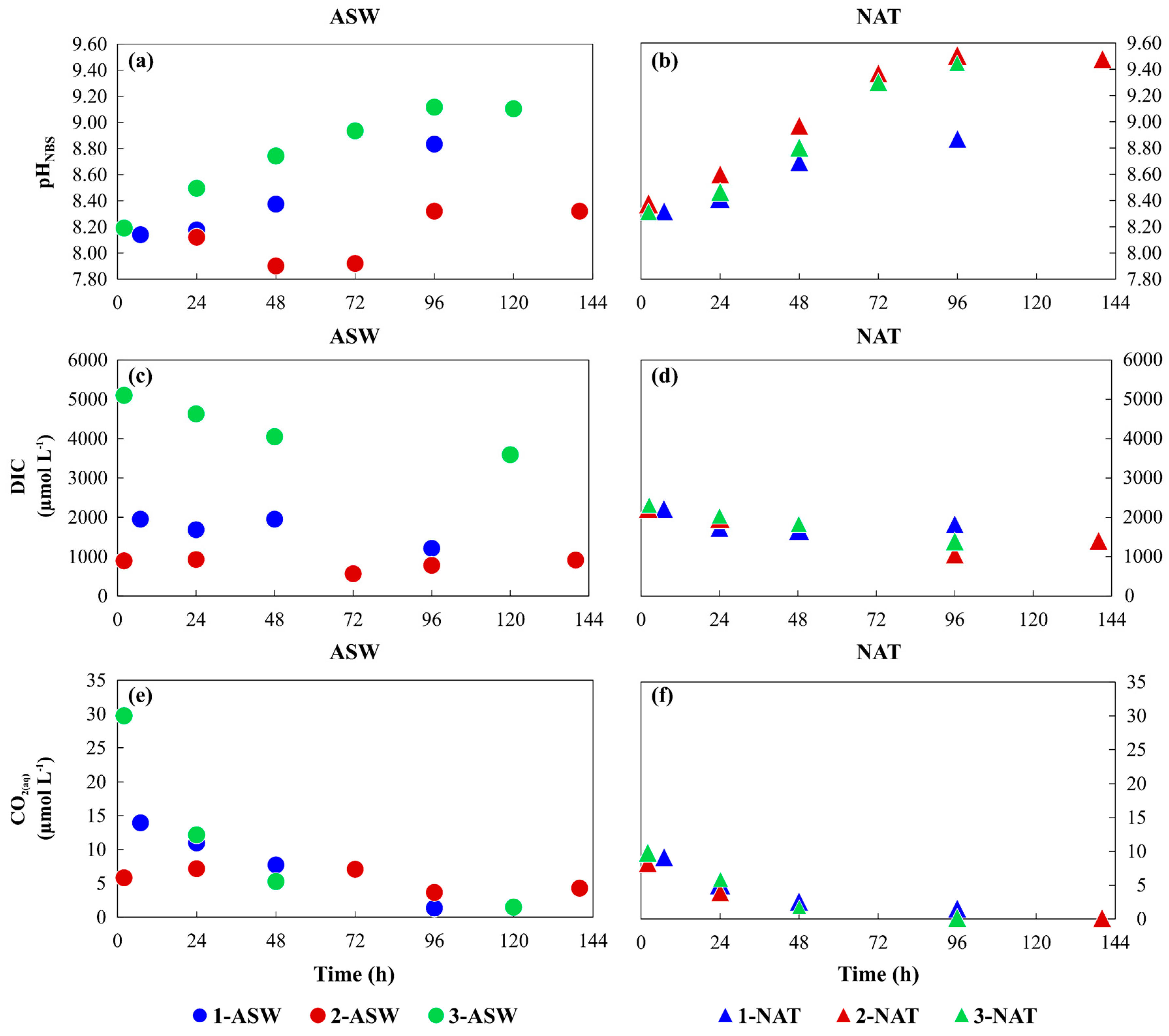
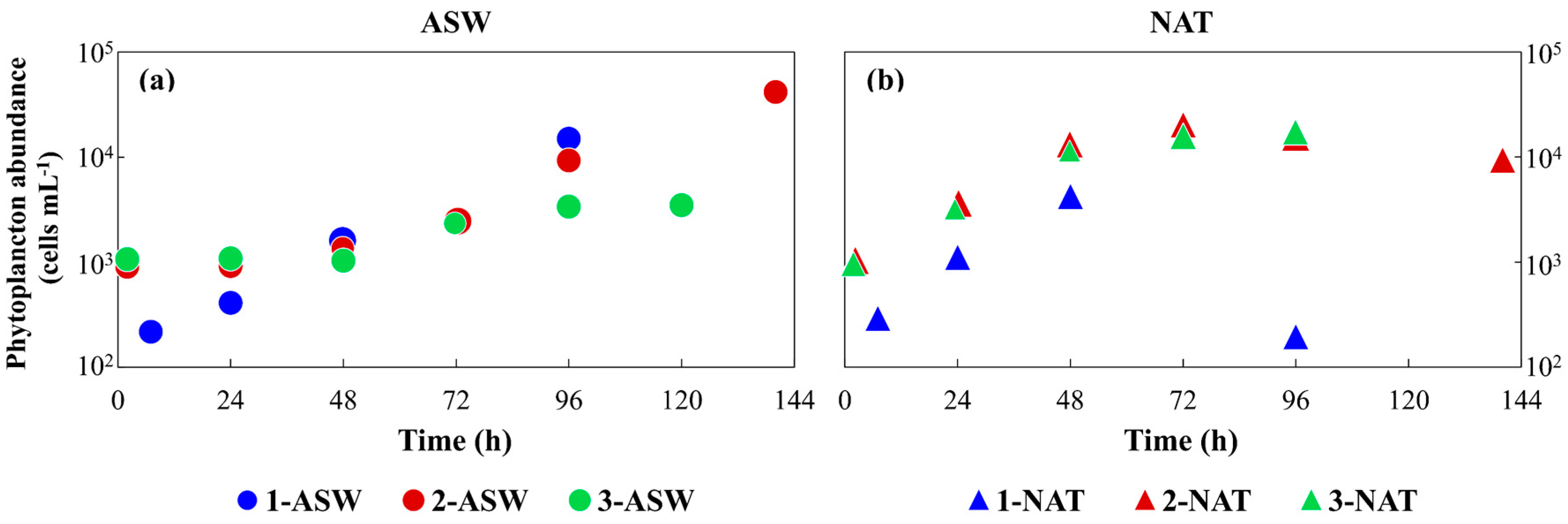
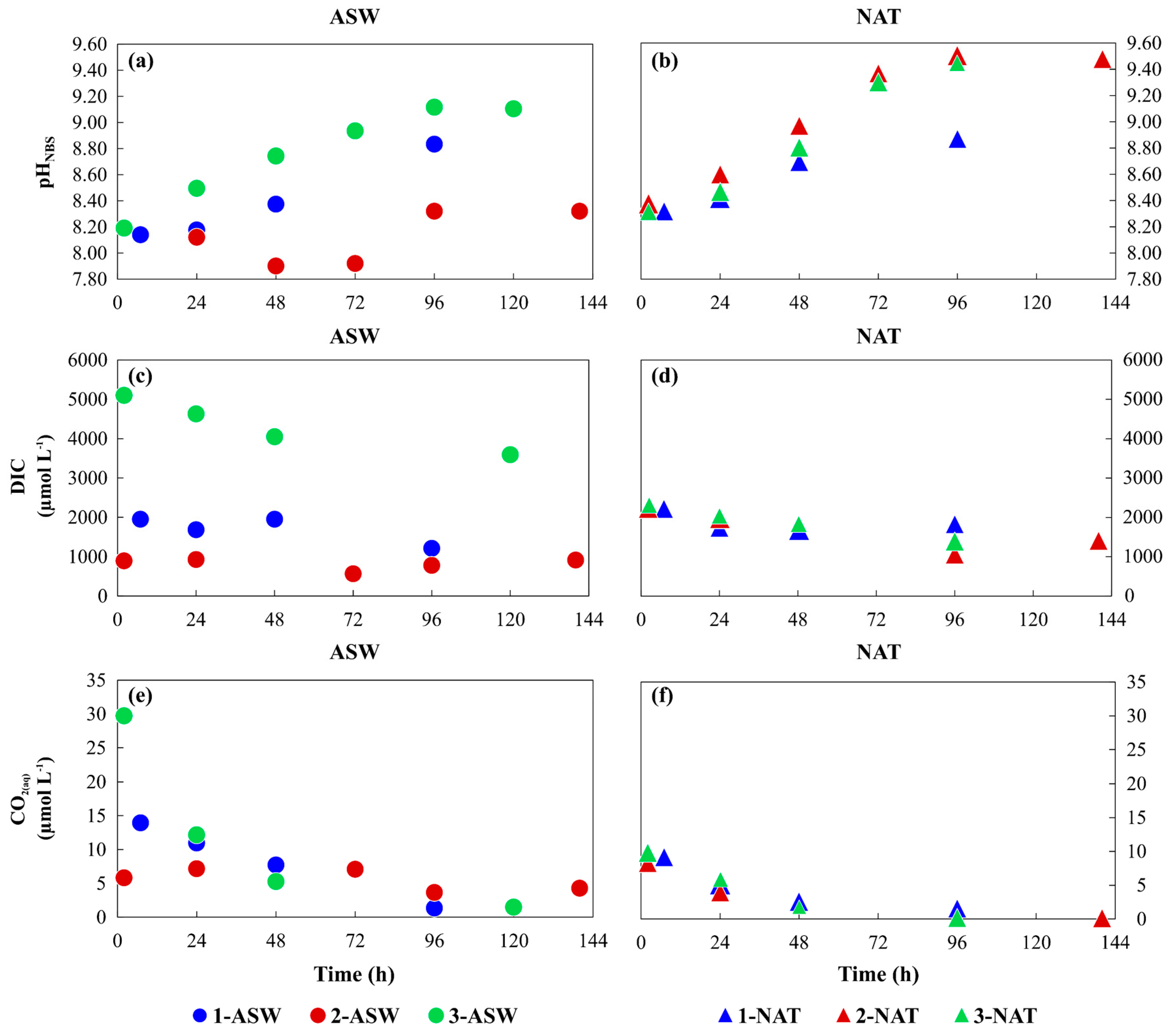
3.1. Phytoplankton Growth under Different Conditions and Media Characterisation
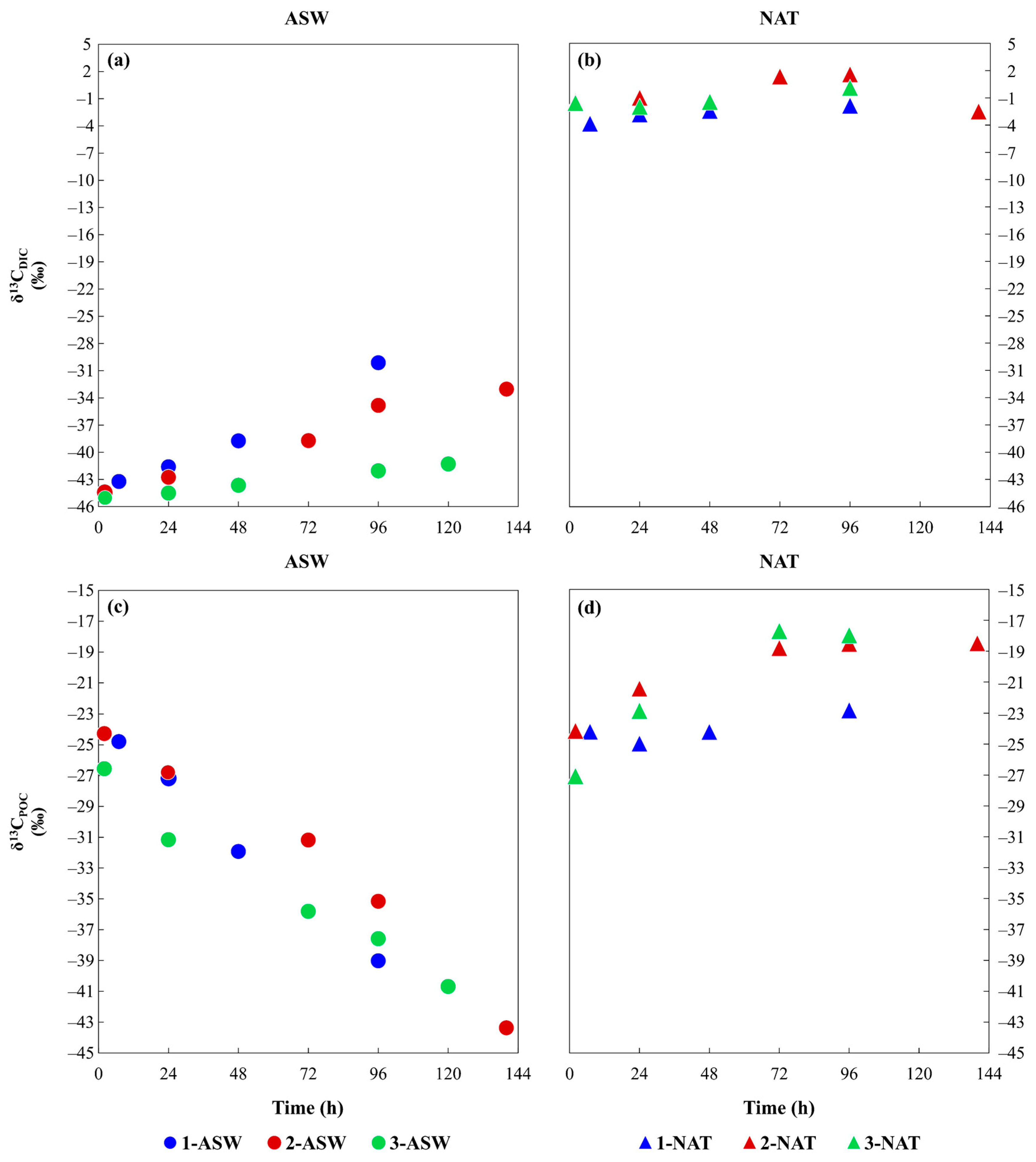
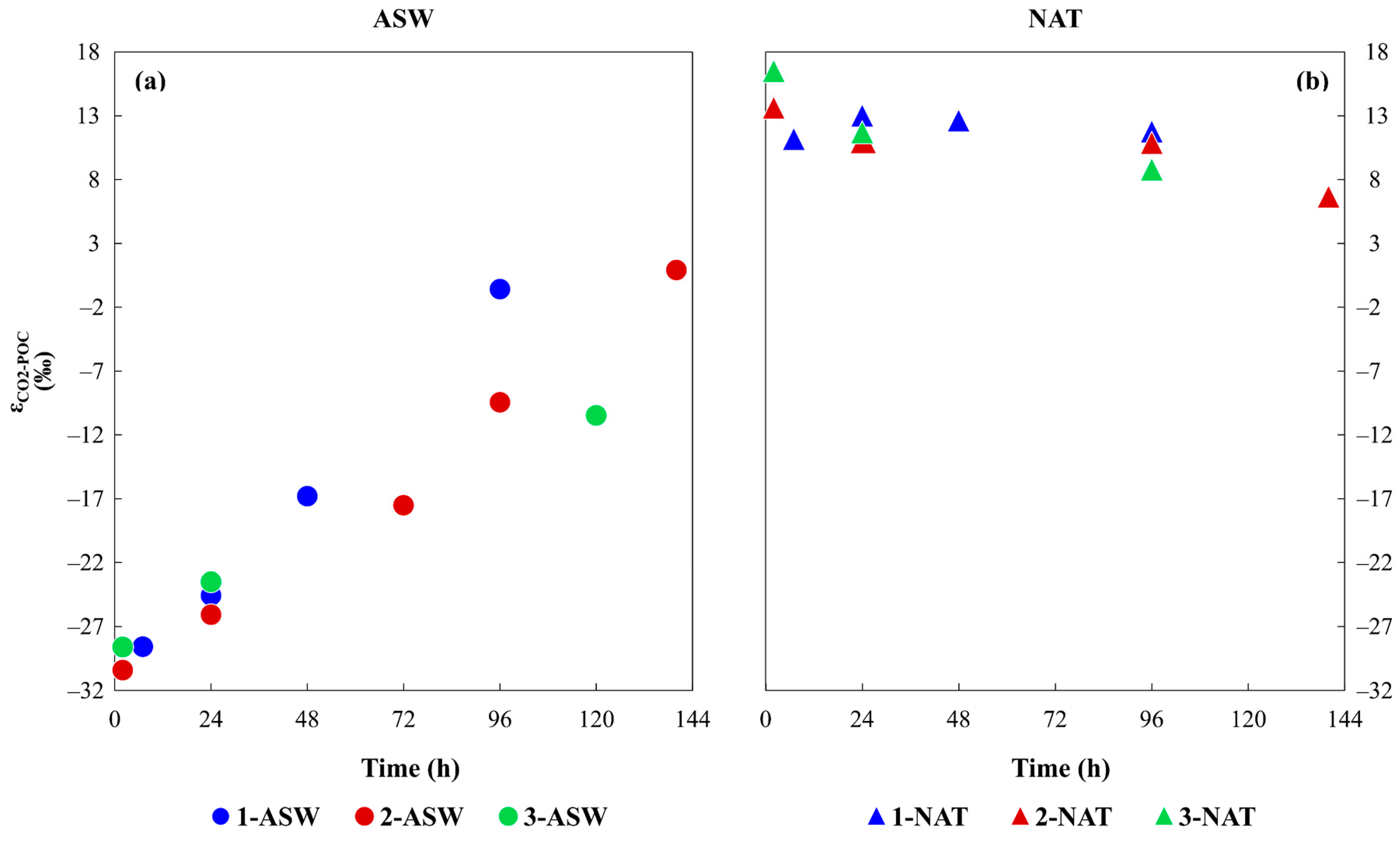
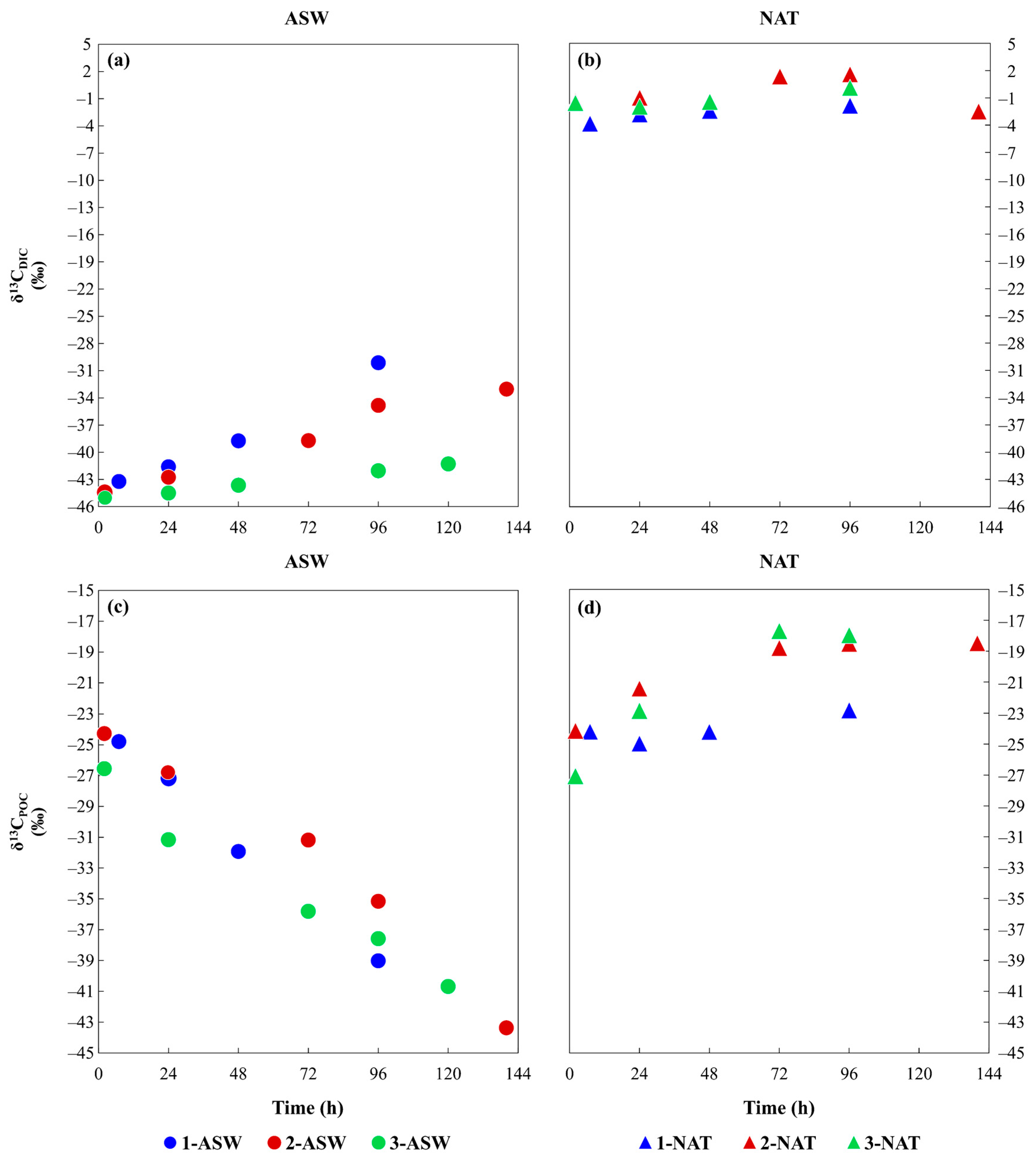
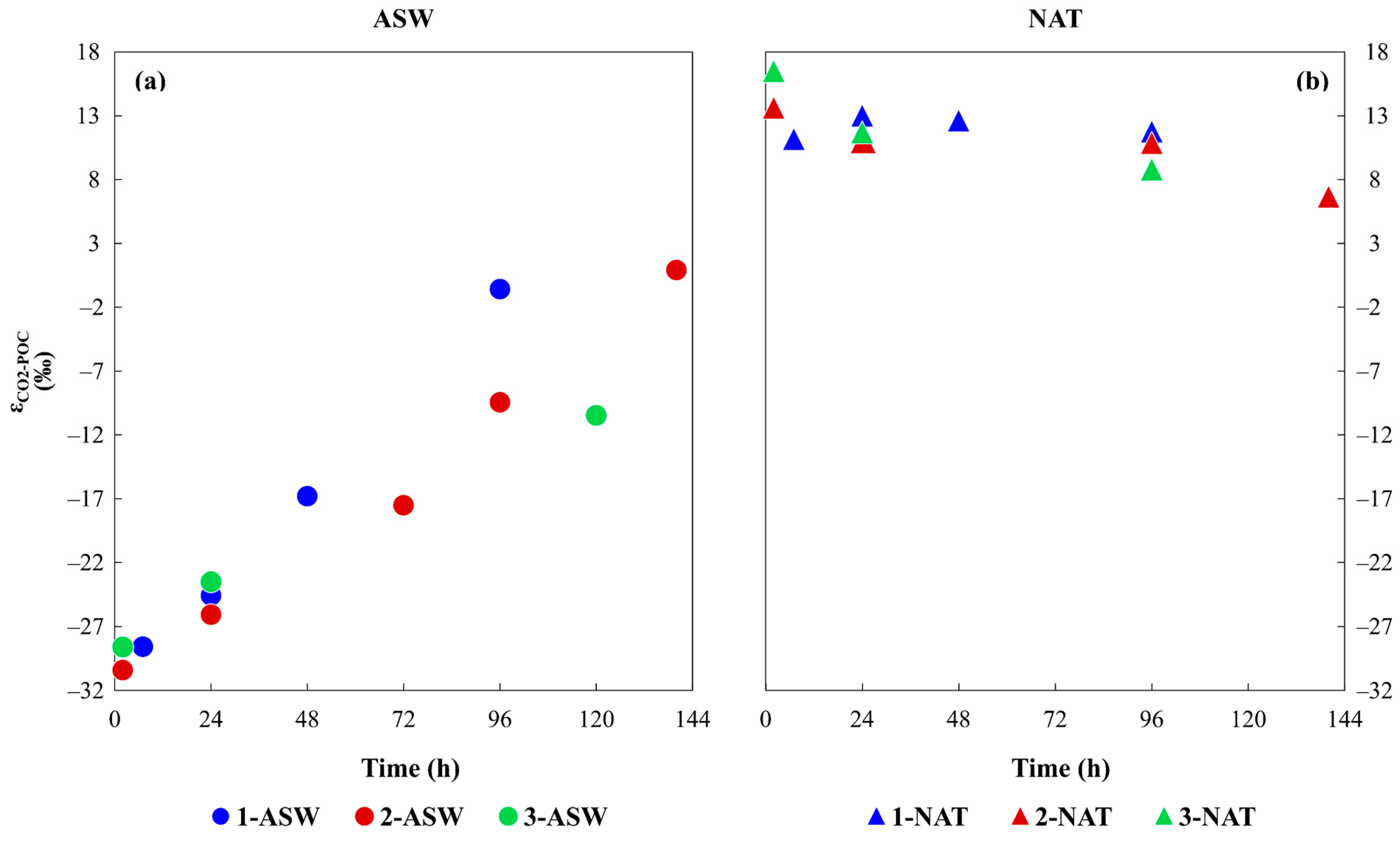
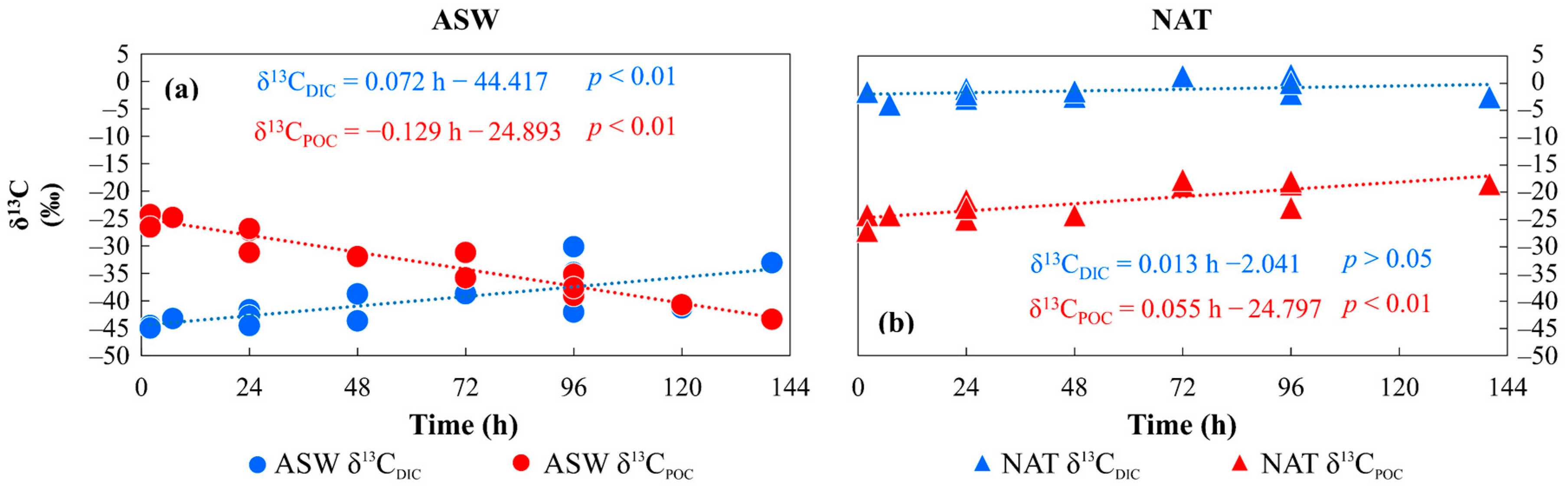
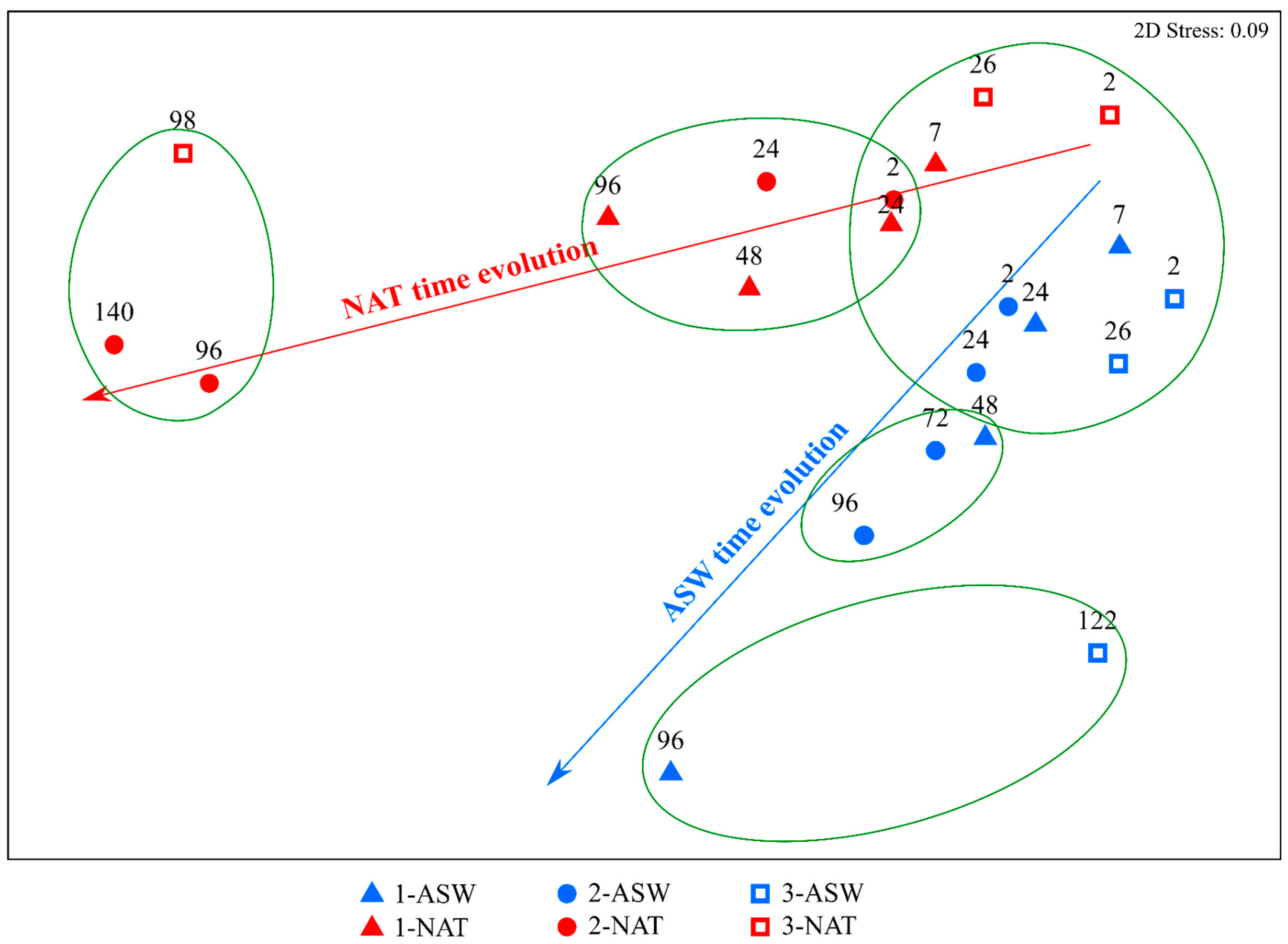
3.2. Isotopic Characterisation of the Media and Phytoplankton
4. Discussion
4.1. Validation of the Experimental Set-Up and Diatom Growth in the Natural and Modified Media
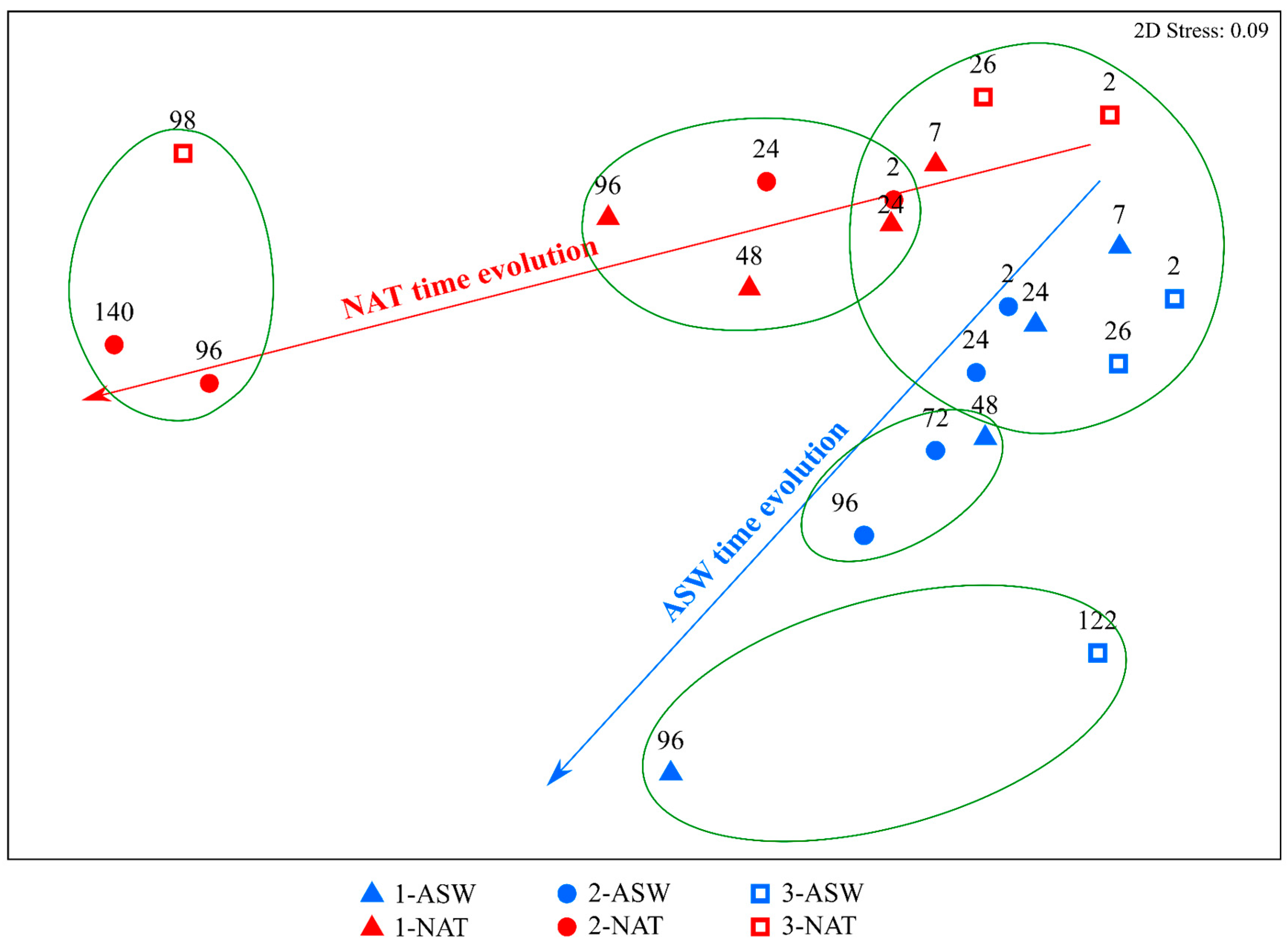
4.2. Time Variation of Phytoplankton Isotopic Composition as a Result of Different DIC Sources and the Inorganic Carbon Uptake
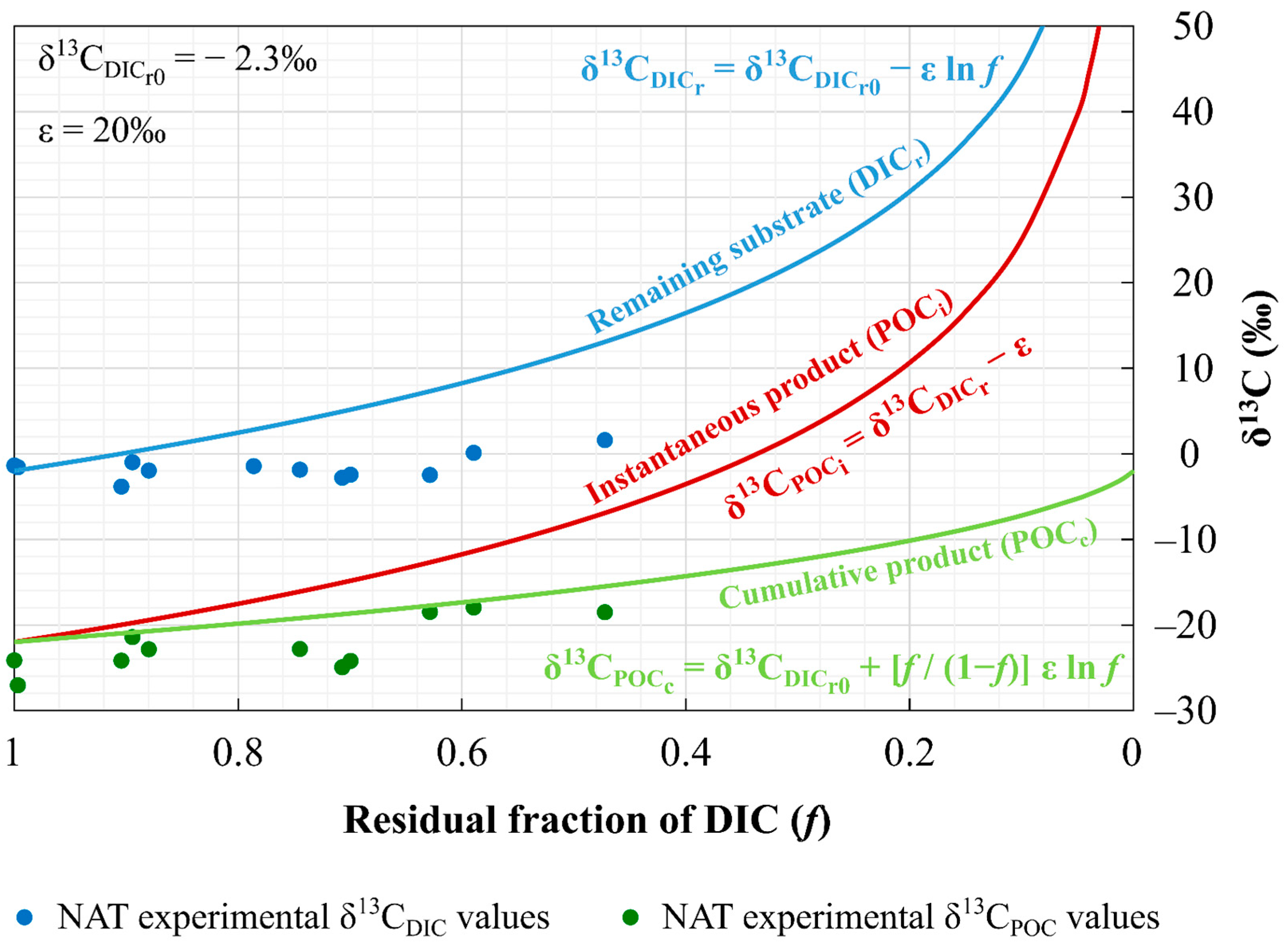
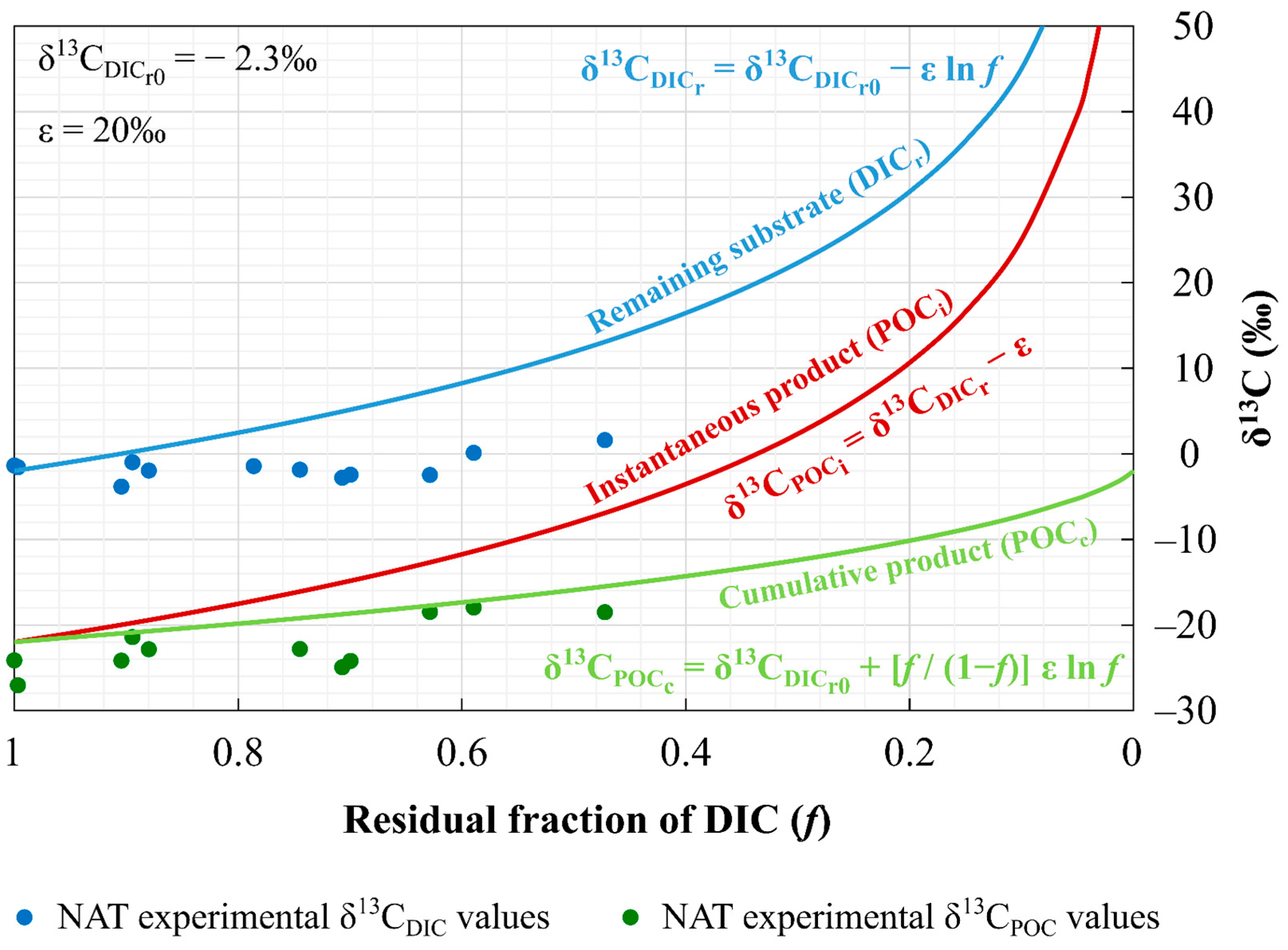
4.3. Phytoplankton Isotopic Fractionation
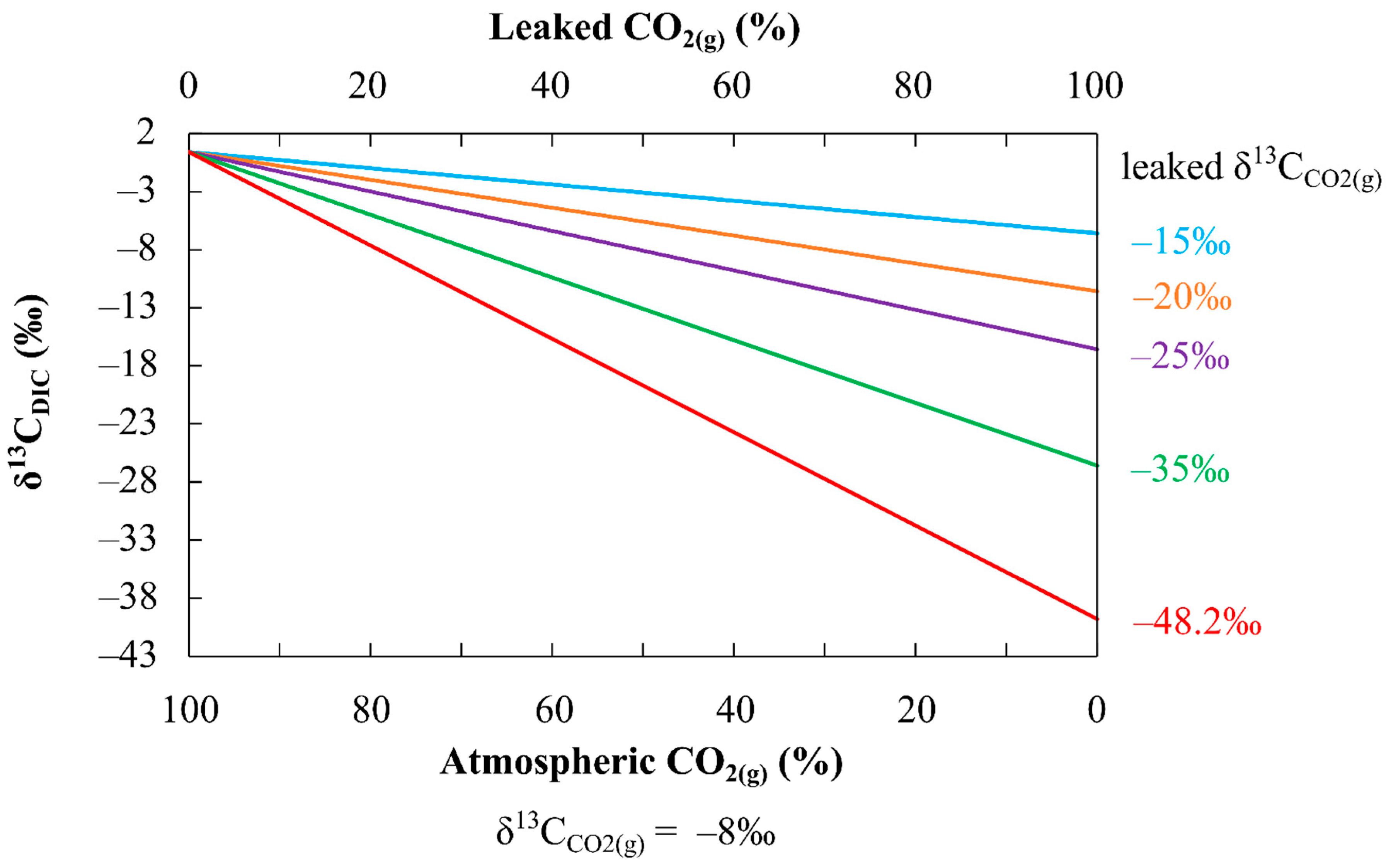
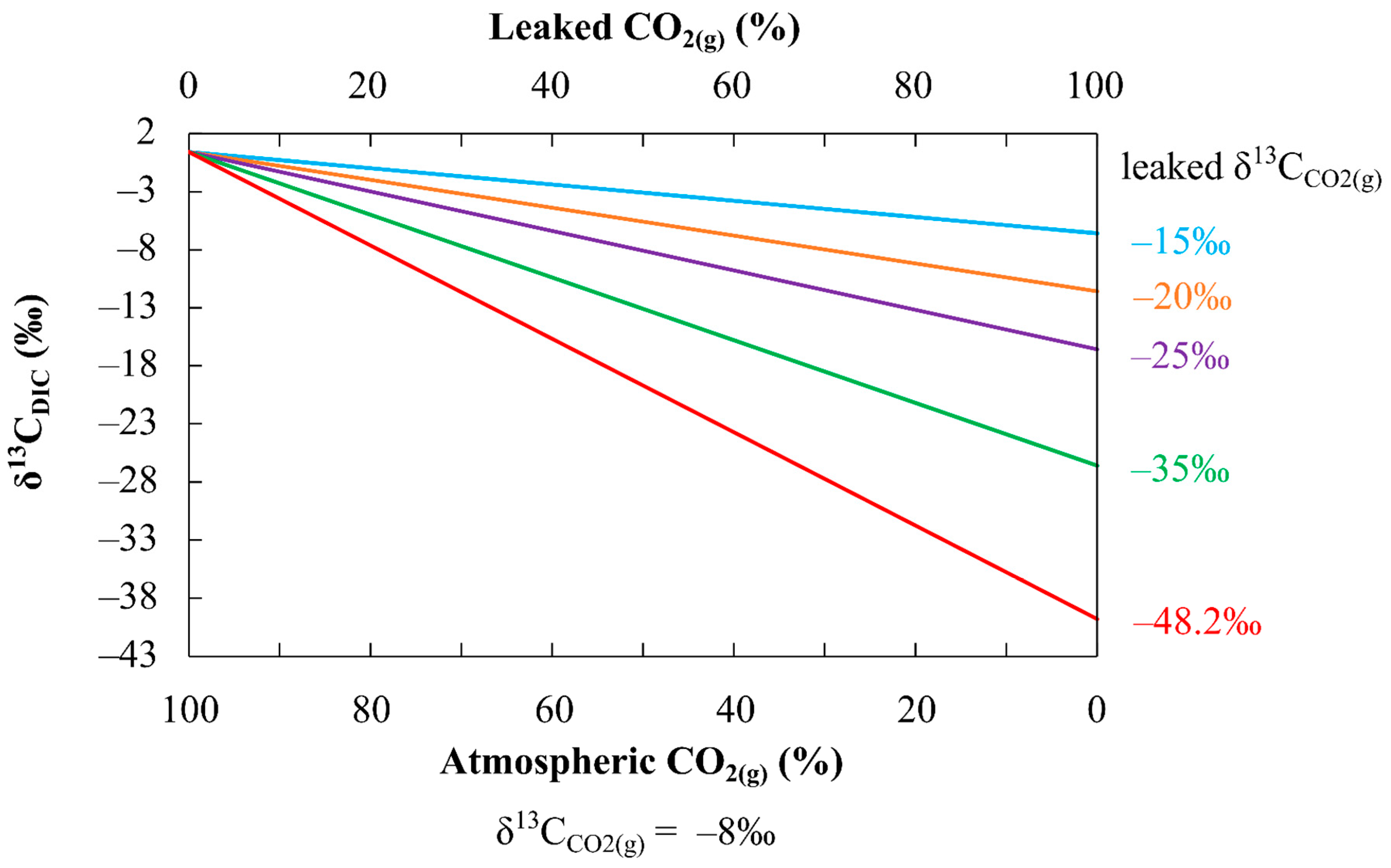
4.4. Application of the Method to CCS Sites: Limitations and Future Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- UNFCCC. Report of the Conference of the Parties on Its Twenty-First Session, Held in Paris from 30 November to 13 December 2015. Decision 1/CP.21. Report no.: FCCC/CP/2015/10 Add. 1. 2015. Available online: https://unfccc.int/resource/docs/2015/cop21/eng/10a01.pdf (accessed on 18 December 2020).
- Haszeldine, R.S.; Flude, S.; Johnson, G.; Scott, V. Negative emissions technologies and carbon capture and storage to achieve the Paris Agreement commitments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2018, 376, 20160447. [Google Scholar] [CrossRef] [Green Version]
- Blackford, J.C.; Alendal, G.; Avlesen, H.; Brereton, A.; Cazenave, P.W.; Chen, B.; Dewar, M.; Holt, J.; Phelps, J. Impact and detectability of hypothetical CCS offshore seep scenarios as an aid to storage assurance and risk assessment. Int. J. Greenh. Gas Control 2020, 95, 102949. [Google Scholar] [CrossRef]
- IPCC. IPCC special report on carbon dioxide capture and storage. In Working Group III of the Intergovernmental Panel on Climate Change; Metz, B., Davidson, O., de Coninck, H.C., Loos, M., Meyer, L.A., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2005. [Google Scholar]
- Flude, S.; Johnson, G.; Gilfillan, S.M.V.; Haszeldine, R.S. Inherent tracers for carbon capture and storage in sedimentary formations: Composition and applications. Environ. Sci. Technol. 2016, 50, 7939–7955. [Google Scholar] [CrossRef]
- IEAGHG. CCS Industry Build-out Rates—Comparison with Industry Analogues; Report No.: 2017-TR6; IEAGHG: Cheltenham, UK, 2017. [Google Scholar]
- Global CCS Institute. Global status of CCS; Global CCS Institute: Melbourne, VIC, Australia, 2019. [Google Scholar]
- IEAGHG. Review of Offshore Monitoring for CCS Projects; IEAGHG: Cheltenham, UK, 2015. [Google Scholar]
- Roberts, J.J.; Stalker, L. What have we learnt about CO2 leakage from CO2 release field experiments, and what are the gaps for the future? Earth-Sci. Rev. 2020, 209, 102939. [Google Scholar] [CrossRef]
- Blackford, J.C.; Stahl, H.; Bull, J.M.; Bergès, B.J.P.; Cevatoglu, M.; Lichtschlag, A.; Connelly, D.; James, R.H.; Kita, J.; Long, D.; et al. Detection and impacts of leakage from sub-seafloor deep geological carbon dioxide storage. Nat. Clim. Chang. 2014, 4, 1011–1016. [Google Scholar] [CrossRef]
- Jones, D.G.; Beaubien, S.E.; Blackford, J.C.; Foekema, E.M.; Lions, J.; De Vittor, C.; West, J.M.; Widdicombe, S.; Hauton, C.; Queirós, A.M. Developments since 2005 in understanding potential environmental impacts of CO2 leakage from geological storage. Int. J. Greenh. Gas Control 2015, 40, 350–377. [Google Scholar] [CrossRef] [Green Version]
- Noble, R.R.P.; Stalker, L.; Wakelin, S.A.; Pejcic, B.; Leybourne, M.I.; Hortle, A.L.; Michael, K. Biological monitoring for carbon capture and storage—a review and potential future developments. Int. J. Greenh. Gas Control 2012, 10, 520–535. [Google Scholar] [CrossRef]
- Shi, D.; Xu, Y.; Hopkinson, B.M.; Morel, F.M.M. Effect of ocean acidification on iron availability to marine phytoplankton. Science 2010, 327, 676–679. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, Y.H.; Kang, S.G.; Park, Y.G. Development of environmental impact monitoring protocol for offshore carbon capture and storage (CCS): A biological perspective. Environ. Impact Assess. Rev. 2016, 57, 139–150. [Google Scholar] [CrossRef]
- Gao, K.; Beardall, J.; Häder, D.P.; Hall-Spencer, J.M.; Gao, G.; Hutchins, D.A. Effects of ocean acidification on marine photosynthetic organisms under the concurrent influences of warming, UV radiation, and deoxygenation. Front. Mar. Sci. 2019, 6, 1–18. [Google Scholar] [CrossRef]
- Li, F.; Beardall, J.; Collins, S.; Gao, K. Decreased photosynthesis and growth with reduced respiration in the model diatom Phaeodactylum tricornutum grown under elevated CO2 over 1800 generations. Glob. Chang. Biol. 2017, 23, 127–137. [Google Scholar] [CrossRef]
- Gao, K.; Campbell, D.A. Photophysiological responses of marine diatoms to elevated CO2 and decreased pH: A review. Funct. Plant Biol. 2014, 41, 449–459. [Google Scholar] [CrossRef]
- Liu, N.; Beardall, J.; Gao, K. Elevated CO2 and associated seawater chemistry do not benefit a model diatom grown with increased availability of light. Aquat. Microb. Ecol. 2017, 79, 137–147. [Google Scholar] [CrossRef]
- Eberlein, T.; Wohlrab, S.; Rost, B.; John, U.; Bach, L.T.; Riebesell, U.; Van De Waal, D.B. Effects of ocean acidification on primary production in a coastal North Sea phytoplankton community. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tait, K.; Stahl, H.; Taylor, P.; Widdicombe, S. Rapid response of the active microbial community to CO2 exposure from a controlled sub-seabed CO2 leak in Ardmucknish Bay (Oban, Scotland). Int. J. Greenh. Gas Control 2015, 38, 171–181. [Google Scholar] [CrossRef]
- Cao, C.; Liu, H.; Hou, Z.; Mehmood, F.; Liao, J.; Feng, W. A review of CO2 storage in view of safety and cost-effectiveness. Energies 2020, 13, 600. [Google Scholar] [CrossRef] [Green Version]
- Blackford, J.C.; Bull, J.M.; Cevatoglu, M.; Connelly, D.; Hauton, C.; James, R.H.; Lichtschlag, A.; Stahl, H.; Widdicombe, S.; Wright, I.C. Marine baseline and monitoring strategies for carbon dioxide capture and storage (CCS). Int. J. Greenh. Gas Control 2015, 38, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, G.; Vizzini, S. Assessing anthropogenic pressures on coastal marine ecosystems using stable CNS isotopes: State of the art, knowledge gaps, and community-scale perspectives. Estuar. Coast. Shelf Sci. 2015, 156, 195–204. [Google Scholar] [CrossRef]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Mayer, B.; Humez, P.; Becker, V.; Dalkhaa, C.; Rock, L.; Myrttinen, A.; Barth, J.A.C. Assessing the usefulness of the isotopic composition of CO2 for leakage monitoring at CO2 storage sites: A review. Int. J. Greenh. Gas Control 2015, 37, 46–60. [Google Scholar] [CrossRef]
- Shin, W.J.; Ryu, J.S.; Choi, H.B.; Yun, S.T.; Lee, K.S. Monitoring the movement of artificially injected CO2 at a shallow experimental site in Korea using carbon isotopes. J. Environ. Manag. 2020, 258, 110030. [Google Scholar] [CrossRef]
- Flude, S.; Györe, D.; Stuart, F.M.; Zurakowska, M.; Boyce, A.J.; Haszeldine, R.S.; Chalaturnyk, R.; Gilfillan, S.M.V. The inherent tracer fingerprint of captured CO2. Int. J. Greenh. Gas Control 2017, 65, 40–54. [Google Scholar] [CrossRef]
- Warwick, P.D.; Ruppert, L.F. Carbon and oxygen isotopic composition of coal and carbon dioxide derived from laboratory coal combustion: A preliminary study. Int. J. Coal Geol. 2016, 166, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Hellevang, H.; Aagaard, P. Constraints on natural global atmospheric CO2 fluxes from 1860 to 2010 using a simplified explicit forward model. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Keeling, C.D. The Suess effect: 13Carbon-14Carbon interrelations. Environ. Int. 1979, 2, 229–300. [Google Scholar] [CrossRef]
- Zeebe, R.E.; Wolf-Gladrow, D.A. CO2 in Seawater: Equilibrium, Kinetics, Isotopes; Oceanography Series 65; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Magozzi, S.; Yool, A.; Vander Zanden, H.B.; Wunder, M.B.; Trueman, C.N. Using ocean models to predict spatial and temporal variation in marine carbon isotopes. Ecosphere 2017, 8, e01763. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.W.; Hamady, L.L.; Thorrold, S.R. A review of ecogeochemistry approaches to estimating movements of marine animals. Limnol. Oceanogr. 2013, 58, 697–714. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Normandeau, C.; Bowden, R.; Doucett, R.; Gallagher, B.; Gillikin, D.P.; Kumamoto, Y.; McKay, J.L.; Middlestead, P.; Ninnemann, U.; et al. An international intercomparison of stable carbon isotope composition measurements of dissolved inorganic carbon in seawater. Limnol. Oceanogr. Methods 2019, 17, 200–209. [Google Scholar] [CrossRef]
- Mook, W.G.; Bommerson, J.C.; Staverman, W.H. Carbon isotope fractionation between dissolved bicarbonate and gaseous carbon dioxide. Earth Planet. Sci. Lett. 1974, 22, 169–176. [Google Scholar] [CrossRef]
- Raven, J.A. Physiology of inorganic C acquisition and implications for resource use efficiency by marine phytoplankton: Relation to increased CO2 and temperature. Plant. Cell Environ. 1991, 14, 779–794. [Google Scholar] [CrossRef]
- Tcherkez, G.G.B.; Farquhar, G.D.; Andrews, T.J. Despite slow catalysis and confused substrate specificity, all ribulose bisphosphate carboxylases may be nearly perfectly optimized. Proc. Natl. Acad. Sci. USA 2006, 103, 7246–7251. [Google Scholar] [CrossRef] [Green Version]
- Baird, M.E. Using a phytoplankton growth model to predict the fractionation of stable carbon isotopes. J. Plankton Res. 2001, 23, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Laws, E.A.; Popp, B.N.; Bidigare, R.R.; Kennicutt, M.C.; Macko, S.A. Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2]aq: Theoretical considerations and experimental results. Geochim. Cosmochim. Acta 1995, 59, 1131–1138. [Google Scholar] [CrossRef]
- Popp, B.N.; Laws, E.A.; Bidigare, R.R.; Dore, J.E.; Hanson, K.L.; Wakeham, S.G. Effect of phytoplankton cell geometry on carbon isotopic fractionation. Geochim. Cosmochim. Acta 1998, 62, 69–77. [Google Scholar] [CrossRef]
- Bidigare, R.R.; Fluegge, A.; Freeman, K.H.; Hanson, K.L.; Hayes, J.M.; Hollander, D.; Jasper, J.P.; King, L.L.; Laws, E.A.; Milder, J.; et al. Consistent fractionation of 13C in nature and in the laboratory: Growth-rate effects in some haptophyte algae. Glob. Biogeochem. Cycles 1997, 11, 279–292. [Google Scholar]
- Nimer, N.A.; Merrett, M.J. Calcification rate in Emiliania huxleyi Lohmann in response to light, nitrate and availability of inorganic carbon. New Phytol. 1993, 673–677. [Google Scholar] [CrossRef]
- Nimer, N.A.; Iglesias-Rodríguez, M.D.; Merrett, M.J. Bicarbonate utilization by marine phytoplankton species. J. Phycol. 1997, 33, 625–631. [Google Scholar] [CrossRef]
- Burkhardt, S.; Riebesell, U.; Zondervan, I. Effects of growth rate, CO2 concentration, and cell size on the stable carbon isotope fractionation in marine phytoplankton. Geochim. Cosmochim. Acta 1999, 63, 3729–3741. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, T. Phytoplanktonic carbon isotope fractionation: Equations accounting for CO2-concentrating mechanisms. J. Plankton Res. 1997, 19, 1455–1476. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Berry, J.A. Carbon isotope fractionation of algae as influenced by an inducible CO2 concentrating mechanism. In Inorganic Carbon Uptake by Aquatic Photosynthetic Organisms; Lucas, W.J., Berry, J.A., Eds.; The American Society of Plan Physiologists: Rockville, MD, USA, 1985; pp. 389–401. [Google Scholar]
- Falkowski, P.G. Species variability in the fractionation of 13C and 12C by marine phytoplankton. J. Plankton Res. 1991, 13, 21–28. [Google Scholar] [CrossRef]
- Ha, J.H.; Jeen, S.W.; Hwang, H.T.; Lee, K.K. Changes in geochemical and carbon isotopic compositions during reactions of CO2-saturated groundwater with aquifer materials. Int. J. Greenh. Gas Control 2020, 95, 102961. [Google Scholar] [CrossRef]
- Mayer, B.; Shevalier, M.; Nightingale, M.; Kwon, J.-S.; Johnson, G.; Raistrick, M.; Hutcheon, I.; Perkins, E. Tracing the movement and the fate of injected CO2 at the IEA GHG Weyburn-Midale CO2 monitoring and storage project (Saskatchewan, Canada) using carbon isotope ratios. Int. J. Greenh. Gas Control 2013, 16, S177–S184. [Google Scholar] [CrossRef]
- Jeandel, E.; Battani, A.; Sarda, P. Lessons learned from natural and industrial analogues for storage of carbon dioxide. Int. J. Greenh. Gas Control 2010, 4, 890–909. [Google Scholar] [CrossRef]
- Shitashima, K.; Maeda, Y.; Sakamoto, A. Detection and monitoring of leaked CO2 through sediment, water column and atmosphere in a sub-seabed CCS experiment. Int. J. Greenh. Gas Control 2015, 38, 135–142. [Google Scholar] [CrossRef]
- Wirtz, K.; Smith, S.L. Vertical migration by bulk phytoplankton sustains biodiversity and nutrient input to the surface ocean. Sci. Rep. 2020, 10, 1142. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [Green Version]
- McClelland, H.L.O.; Bruggeman, J.; Hermoso, M.; Rickaby, R.E.M. The origin of carbon isotope vital effects in coccolith calcite. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Throndsen, J. Preservation and storage. In Phytoplankton Manual. Monographs on Oceanographic Methodology 6; Sournia, A., Ed.; UNESCO: Paris, France, 1978; pp. 69–74. [Google Scholar]
- Gattuso, J.P.; Gao, K.; Lee, K.; Rost, B.; Schulz, K.G. Approaches and tools to manipulate the carbonate chemistry. In Guide for Best Practices in Ocean Acidification Research and Data Reporting; Riebesell, U., Fabry, V.J., Lina, H., Gattuso, J.P., Eds.; Office for Official Publications of the European Union: Luxemburg, 2010. [Google Scholar]
- Dickson, A.G.; Sabine, C.; Christian, J.R. Guide to Best Practices for Ocean CO2 measurements. Standard operating Procedure (SOP) 3b, Determination of Total Alkalinity in Sea Water Using an Open-Cell Titration; PICES Special Publication 3; IOCCP Report 8; North Pacific Marine Science Organization: Sidney, BC, Canada, 2007; 191p. [Google Scholar]
- Ingrosso, G.; Giani, M.; Cibic, T.; Karuza, A.; Kralj, M.; Del Negro, P. Carbonate chemistry dynamics and biological processes along a river-sea gradient (Gulf of Trieste, northern Adriatic Sea). J. Mar. Syst. 2016, 155, 35–49. [Google Scholar] [CrossRef]
- Hansen, H.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; Wiley-VCH: Weinheim, Germany, 1999; pp. 159–228. [Google Scholar]
- Pierrot, D.; Lewis, E.; Wallace, D. MS Excel Program Developed for CO2 System Calculations; ORNL/CDIAC-105a; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy: Oak Ridge, TN, USA, 2006.
- Mehrbach, C.; Culberson, C.; Hawley, J.; Pytkowics, R. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Res. Part A. Oceanogr. Res. Pap. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- Dickson, A.G. Standard potential of the reaction: AgCl(s) + 12H2(g) = Ag(s) + HCl(aq), and the standard acidity constant of the ion HSO4− in synthetic sea water from 273.15 to 318.15 K. J. Chem. 1990, 22, 113–127. [Google Scholar] [CrossRef]
- Uppström, L.R. The boron/chlorinity ratio of deep-sea water from the Pacific Ocean. Deep Sea Res. 1974, 21, 161–162. [Google Scholar] [CrossRef]
- Marie, D.; Brussaard, C.P.D.; Thyrhaug, R.; Bratbak, G.; Vaulot, D. Enumeration of marine viruses in culture and natural samples by flow cytometry. Appl. Environ. Microbiol. 1999, 65, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Celussi, M.; Malfatti, F.; Annalisa, F.; Gazeau, F.; Giannakourou, A.; Pitta, P.; Tsiola, A.; Del Negro, P. Ocean acidification effect on prokaryotic metabolism tested in two diverse trophic regimes in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2017, 186, 125–138. [Google Scholar] [CrossRef]
- Hasle, G.; Syvertson, E. Marine diatoms. In Identifying Marine Phytoplankton; Tomas, C., Ed.; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Rau, G.H.; Riebesell, U.; Wolf-Gladrow, D.A. A model of photosynthetic 13C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake. Mar. Ecol. Prog. Ser. 1996, 133, 275–285. [Google Scholar] [CrossRef]
- Clark, I.D.; Fritz, P. (Eds.) Environmental Isotopes in Hydrogeology, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 1999; ISBN 1566702496. [Google Scholar]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Freeman, K.H.; Hayes, J.M. Fractionation of carbon isotopes by phytoplankton and estimates of ancient CO2 levels. Glob. Biogeochem. Cycles 1992, 6, 185–198. [Google Scholar] [CrossRef]
- Gat, J.R.; Gonfiantini, R. Stable Isotope Hydrology. Deuterium and Oxygen-18 in the Water Cycle; Technical Report 210; International Atomic Energy Agency: Vienna, Austria, 1981. [Google Scholar]
- Laws, E.A.; Thompson, P.A.; Popp, B.N.; Bidigare, R.R. Sources of inorganic carbon for marine microalgal photosynthesis: A reassessment of δ13C data from batch culture studies of Thalassiosira pseudonana and Emiliania huxleyi. Limnol. Oceanogr. 1998, 43, 136–142. [Google Scholar] [CrossRef]
- Zhang, J.; Quay, P.D.; Wilbur, D.O. Carbon isotope fractionation during gas-water exchange and dissolution of CO2. Geochim. Cosmochim. Acta 1995, 59, 107–114. [Google Scholar] [CrossRef]
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Ingrosso, G.; Bensi, M.; Cardin, V.; Giani, M. Anthropogenic CO2 in a dense water formation area of the Mediterranean Sea. Deep. Res. Part I Oceanogr. Res. Pap. 2017, 123, 118–128. [Google Scholar] [CrossRef]
- Tamše, S.; Ogrinc, N.; Walter, L.M.; Turk, D.; Faganeli, J. River sources of dissolved inorganic carbon in the Gulf of Trieste (N Adriatic): Stable carbon isotope evidence. Estuaries Coasts 2015, 38, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Bade, D.L.; Carpenter, S.R.; Cole, J.J.; Hanson, P.C.; Hesslein, R.H. Controls of δ13C-DIC in lakes: Geochemistry, lake metabolism, and morphometry. Limnol. Oceanogr. 2004, 49, 1160–1172. [Google Scholar] [CrossRef]
- Hinga, K.R. Effects of pH on coastal marine phytoplankton. Mar. Ecol. Prog. Ser. 2002, 238, 281–300. [Google Scholar] [CrossRef]
- Clark, D.R.; Flynn, K.J. The relationship between the dissolved inorganic carbon concentration and growth rate in marine phytoplankton. Proc. R. Soc. B Biol. Sci. 2000, 267, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.R.; Flynn, K.J.; Fabian, H. Variation in elemental stoichiometry of marine diatom Thalassiosira weissflogii (Bacillariophyceae) in response to combined nutrient stress and changes in carbonate chemistry. J. Phycol. 2014, 50, 640–651. [Google Scholar] [CrossRef]
- Maugendre, L.; Gattuso, J.P.; Poulton, A.J.; Dellisanti, W.; Gaubert, M.; Guieu, C.; Gazeau, F. No detectable effect of ocean acidification on plankton metabolism in the NW oligotrophic Mediterranean Sea: Results from two mesocosm studies. Estuar. Coast. Shelf Sci. 2017, 186, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Esposito, M.; Achterberg, E.P.; Bach, L.T.; Connelly, D.P.; Riebesell, U.; Taucher, J. Application of stable carbon isotopes in a subtropical North Atlantic mesocosm study: A new approach to assess CO2 effects in the marine carbon cycle. Front. Mar. Sci. 2019, 6, 616. [Google Scholar] [CrossRef]
- Hurd, C.L.; Hepburn, C.D.; Currie, K.I.; Raven, J.A.; Hunter, K.A. Testing the effects of ocean acidification on algal metabolism: Consideration for experimental designs. J. Phycol. 2009, 45, 1236–1251. [Google Scholar] [CrossRef]
- Li, F.; Fan, J.; Hu, L.; Beardall, J.; Xu, J.; Fields, D. Physiological and biochemical responses of Thalassiosira weissflogii (diatom) to seawater acidification and alkalization. ICES J. Mar. Sci. 2019, 76, 1850–1859. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Dupont, C.L.; Hopkinson, B.M. The diversity of CO2-concentrating mechanisms in marine diatoms as inferred from their genetic content. J. Exp. Bot. 2017, 68, 3937–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colman, B.; Huertas, I.E.; Bhatti, S.; Dason, J.S. The diversity of inorganic carbon acquisition mechanisms in eukaryotic microalgae. Funct. Plant Biol. 2002, 29, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, S.; Amoroso, G.; Riebesell, U.; Sültemeyer, D. CO2 and HCO3- uptake in marine diatoms acclimated to different CO2 concentrations. Limnol. Oceanogr. 2001, 46, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Cozzi, S.; Falconi, C.; Comici, C.; Čermelj, B.; Kovac, N.; Turk, V.; Giani, M. Recent evolution of river discharges in the Gulf of Trieste and their potential response to climate changes and anthropogenic pressure. Estuar. Coast. Shelf Sci. 2012, 115, 14–24. [Google Scholar] [CrossRef]
- Paul, A.J.; Bach, L.T.; Schulz, K.G.; Boxhammer, T.; Czerny, J.; Achterberg, E.P.; Hellemann, D.; Trense, Y.; Nausch, M.; Sswat, M.; et al. Effect of elevated CO2 on organic matter pools and fluxes in a summer Baltic Sea plankton community. Biogeosciences 2015, 12, 6181–6203. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wu, Y.; Hutchins, D.A.; Fu, F.; Gao, K. Physiological responses of coastal and oceanic diatoms to diurnal fluctuations in seawater carbonate chemistry under two CO2 concentrations. Biogeosciences 2016, 13, 6247–6259. [Google Scholar] [CrossRef] [Green Version]
- Tamše, S.; Mozetič, P.; Francé, J.; Ogrinc, N. Stable isotopes as a tool for nitrogen source identification and cycling in the Gulf of Trieste (Northern Adriatic). Cont. Shelf Res. 2014, 91, 145–157. [Google Scholar] [CrossRef]
- Faganeli, J.; Ogrinc, N.; Kovac, N.; Kukovec, K.; Falnoga, I.; Mozeticč, P.; Bajt, O. Carbon and nitrogen isotope composition of particulate organic matter in relation to mucilage formation in the northern Adriatic Sea. Mar. Chem. 2009, 114, 102–109. [Google Scholar] [CrossRef]
- Giani, M.; Berto, D.; Rampazzo, F.; Savelli, F.; Alvisi, F.; Giordano, P.; Ravaioli, M.; Frascari, F. Origin of sedimentary organic matter in the north-western Adriatic Sea. Estuar. Coast. Shelf Sci. 2009, 84, 573–583. [Google Scholar] [CrossRef]
- Poulain, C.; Lorrain, A.; Mas, R.; Gillikin, D.P.; Dehairs, F.; Robert, R.; Paulet, Y.M. Experimental shift of diet and DIC stable carbon isotopes: Influence on shell δ13C values in the Manila clam Ruditapes philippinarum. Chem. Geol. 2010, 272, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Tuerena, R.E.; Ganeshram, R.S.; Humphreys, M.P.; Browning, T.J.; Bouman, H.; Piotrowski, A.P. Isotopic fractionation of carbon during uptake by phytoplankton across the South Atlantic subtropical convergence. Biogeosciences 2019, 16, 3621–3635. [Google Scholar] [CrossRef] [Green Version]
- Henley, S.F.; Annett, A.L.; Ganeshram, R.S.; Carson, D.S.; Weston, K.; Crosta, X.; Tait, A.; Dougans, J.; Fallick, A.E.; Clarke, A. Factors influencing the stable carbon isotopic composition of suspended and sinking organic matter in the coastal Antarctic sea ice environment. Biogeosciences 2012, 9, 1137–1157. [Google Scholar] [CrossRef] [Green Version]
- Coffin, R.B.; Fry, B.; Peterson, B.J.; Wright, R.T. Carbon isotopic compositions of estuarine bacteria. Limnol. Oceanogr. 1989, 34, 1305–1310. [Google Scholar] [CrossRef]
- Hullar, M.; Fry, B.; Peterson, B.J.; Wright, R.T. Microbial utilization of estuarine dissolved organic carbon: A stable isotope tracer approach tested by mass balance. Appl. Environ. Microbiol. 1996, 62, 2489–2493. [Google Scholar] [CrossRef] [Green Version]
- Lammers, J.M.; Reichart, G.J.; Middelburg, J.J. Seasonal variability in phytoplankton stable carbon isotope ratios and bacterial carbon sources in a shallow Dutch lake. Limnol. Oceanogr. 2017, 62, 2773–2787. [Google Scholar] [CrossRef] [Green Version]
- De Kluijver, A.; Soetaert, K.; Schulz, K.G.; Riebesell, U.; Bellerby, R.G.J.; Middelburg, J.J. Phytoplankton-bacteria coupling under elevated CO2 levels: A stable isotope labelling study. Biogeosciences 2010, 7, 3783–3797. [Google Scholar] [CrossRef] [Green Version]
- Morales-Williams, A.M.; Wanamaker, A.D.; Downing, J.A. Cyanobacterial carbon concentrating mechanisms facilitate sustained CO2 depletion in eutrophic lakes. Biogeosciences 2017, 14, 2865–2875. [Google Scholar] [CrossRef] [Green Version]
- Boller, A.J.; Thomas, P.J.; Cavanaugh, C.M.; Scott, K.M. Isotopic discrimination and kinetic parameters of RubisCO from the marine bloom-forming diatom, Skeletonema costatum. Geobiology 2015, 13, 33–43. [Google Scholar] [CrossRef]
- Young, J.N.; Bruggeman, J.; Rickaby, R.E.M.; Erez, J.; Conte, M. Evidence for changes in carbon isotopic fractionation by phytoplankton between 1960 and 2010. Glob. Biogeochem. Cycles 2013, 27, 505–515. [Google Scholar] [CrossRef]
- Beardall, J.; Raven, J.A. The potential effects of global climate change on microalgal photosynthesis, growth and ecology. Phycologia 2004, 43, 26–40. [Google Scholar] [CrossRef]
- Rayleigh, L. LIX. On the distillation of binary mixtures. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1902, 4, 521–537. [Google Scholar] [CrossRef] [Green Version]
- Ohkouchi, N.; Ogawa, N.O.; Chikaraishi, Y.; Tanaka, H.; Wada, E. Biochemical and physiological bases for the use of carbon and nitrogen isotopes in environmental and ecological studies. Prog. Earth Planet. Sci. 2015, 2, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Riebesell, U.; Wolf-Gladrow, D.A. Growth limits on phytoplankton. Nature 1995, 373, 28. [Google Scholar] [CrossRef]
- Wang, S.; Yeager, K.M.; Lu, W. Carbon isotope fractionation in phytoplankton as a potential proxy for pH rather than for [CO2(aq)]: Observations from a carbonate lake. Limnol. Oceanogr. 2016, 61, 1259–1270. [Google Scholar] [CrossRef]
- Pham, L.H.H.P.; Rusli, R.; Shariff, A.M.; Khan, F. Dispersion of carbon dioxide bubble release from shallow subsea carbon dioxide storage to seawater. Cont. Shelf Res. 2020, 196, 104075. [Google Scholar] [CrossRef]
- Cotovicz, L.C.; Knoppers, B.A.; Deirmendjian, L.; Abril, G. Sources and sinks of dissolved inorganic carbon in an urban tropical coastal bay revealed by δ13C-DIC signals. Estuar. Coast. Shelf Sci. 2019, 220, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Alverson, A.J. Molecular systematics and the diatom species. Protist 2008, 159, 339–353. [Google Scholar] [CrossRef] [Green Version]
- MacGillivary, M.L.; Kaczmarska, I. Survey of the efficacy of a short fragment of the rbcL gene as a supplemental DNA barcode for diatoms. J. Eukaryot. Microbiol. 2011, 58, 529–536. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Altschul, S. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Medium Tested | Experiment Code | Initial Cell Abundance (Cells mL−1) | Initial DIC Concentration (µmol L−1) | Initial pH (pHNBS) | Experiment Duration (h) |
|---|---|---|---|---|---|---|
| EXP-1 | ASW | 1-ASW | 250 | 1952 | 8.14 | 96 |
| NAT | 1-NAT | 250 | 2222 | 8.32 | 96 | |
| EXP-2 | ASW | 2-ASW | 1000 | 893 | 8.19 | 140 |
| NAT | 2-NAT | 1000 | 2243 | 8.37 | 140 | |
| EXP-3 | ASW | 3-ASW | 1000 | 5098 | 8.19 | 120 |
| NAT | 3-NAT | 1000 | 2350 | 8.32 | 96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relitti, F.; Ogrinc, N.; Giani, M.; Cerino, F.; Smodlaka Tankovic, M.; Baricevic, A.; Urbini, L.; Krajnc, B.; Del Negro, P.; De Vittor, C. Stable Carbon Isotopes of Phytoplankton as a Tool to Monitor Anthropogenic CO2 Submarine Leakages. Water 2020, 12, 3573. https://doi.org/10.3390/w12123573
Relitti F, Ogrinc N, Giani M, Cerino F, Smodlaka Tankovic M, Baricevic A, Urbini L, Krajnc B, Del Negro P, De Vittor C. Stable Carbon Isotopes of Phytoplankton as a Tool to Monitor Anthropogenic CO2 Submarine Leakages. Water. 2020; 12(12):3573. https://doi.org/10.3390/w12123573
Chicago/Turabian StyleRelitti, Federica, Nives Ogrinc, Michele Giani, Federica Cerino, Mirta Smodlaka Tankovic, Ana Baricevic, Lidia Urbini, Bor Krajnc, Paola Del Negro, and Cinzia De Vittor. 2020. "Stable Carbon Isotopes of Phytoplankton as a Tool to Monitor Anthropogenic CO2 Submarine Leakages" Water 12, no. 12: 3573. https://doi.org/10.3390/w12123573
APA StyleRelitti, F., Ogrinc, N., Giani, M., Cerino, F., Smodlaka Tankovic, M., Baricevic, A., Urbini, L., Krajnc, B., Del Negro, P., & De Vittor, C. (2020). Stable Carbon Isotopes of Phytoplankton as a Tool to Monitor Anthropogenic CO2 Submarine Leakages. Water, 12(12), 3573. https://doi.org/10.3390/w12123573