Fish Community Responses to Human-Induced Stresses in the Lower Mekong Basin


Abstract
:1. Introduction
2. Materials and Methods
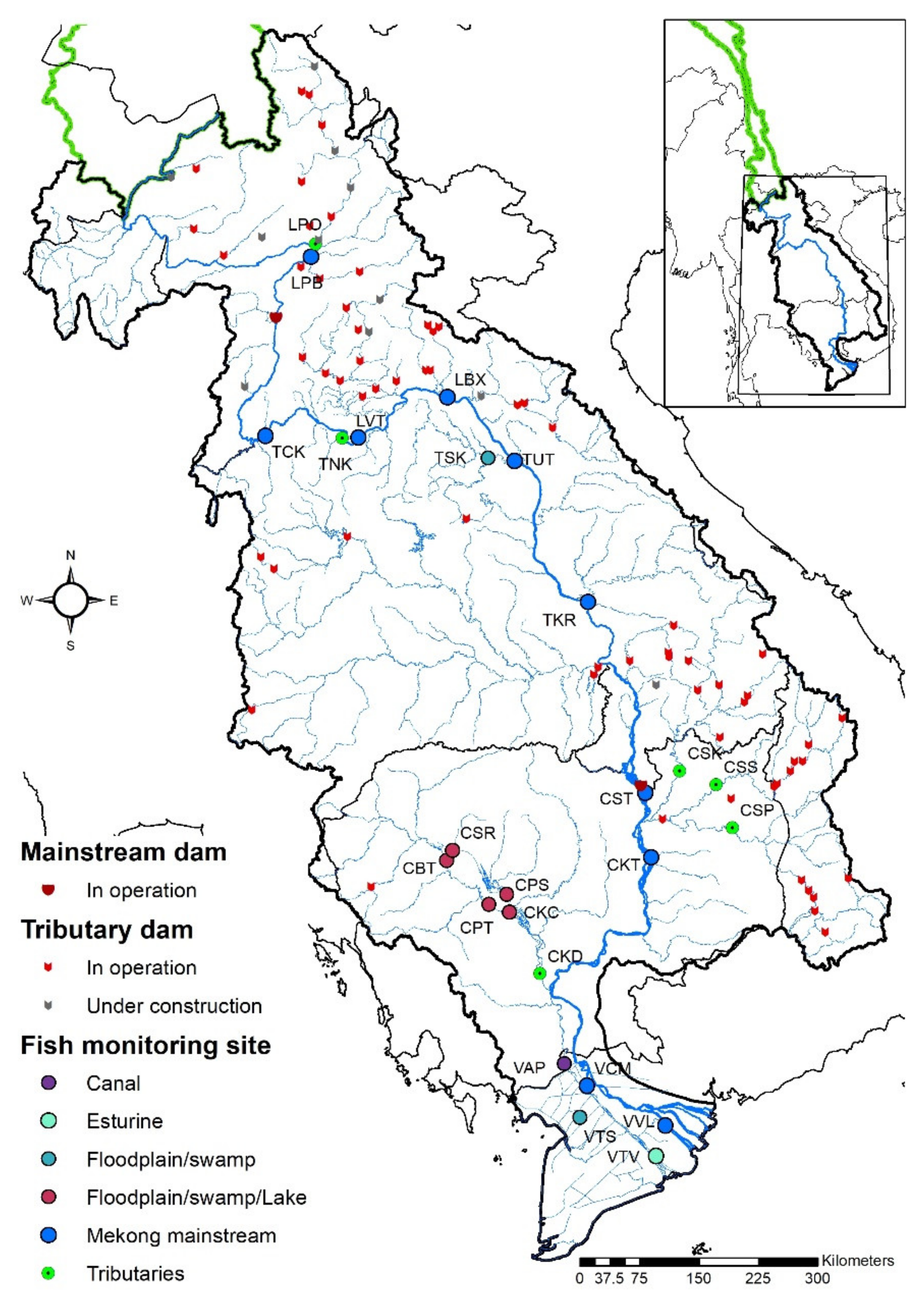
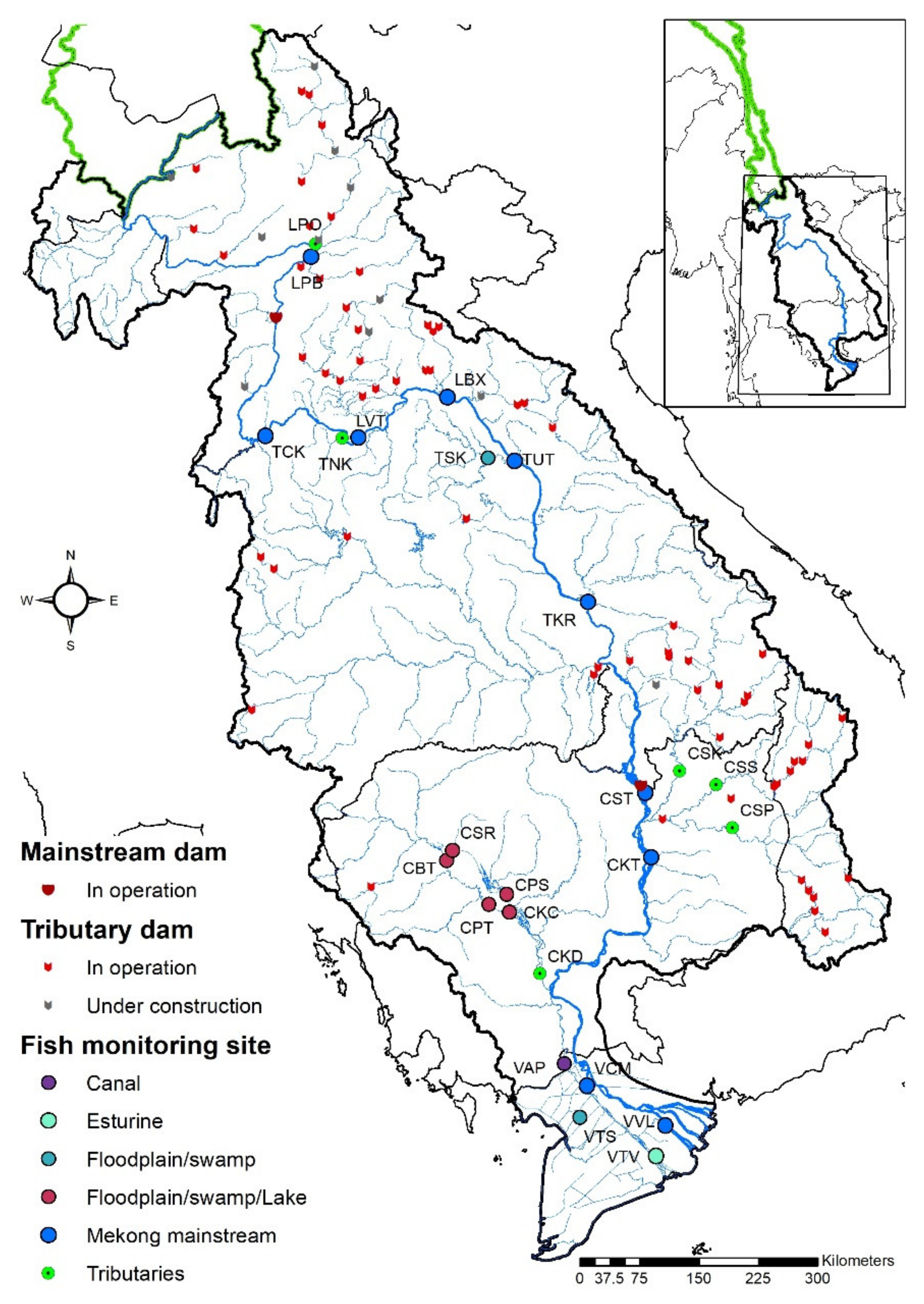
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analysis
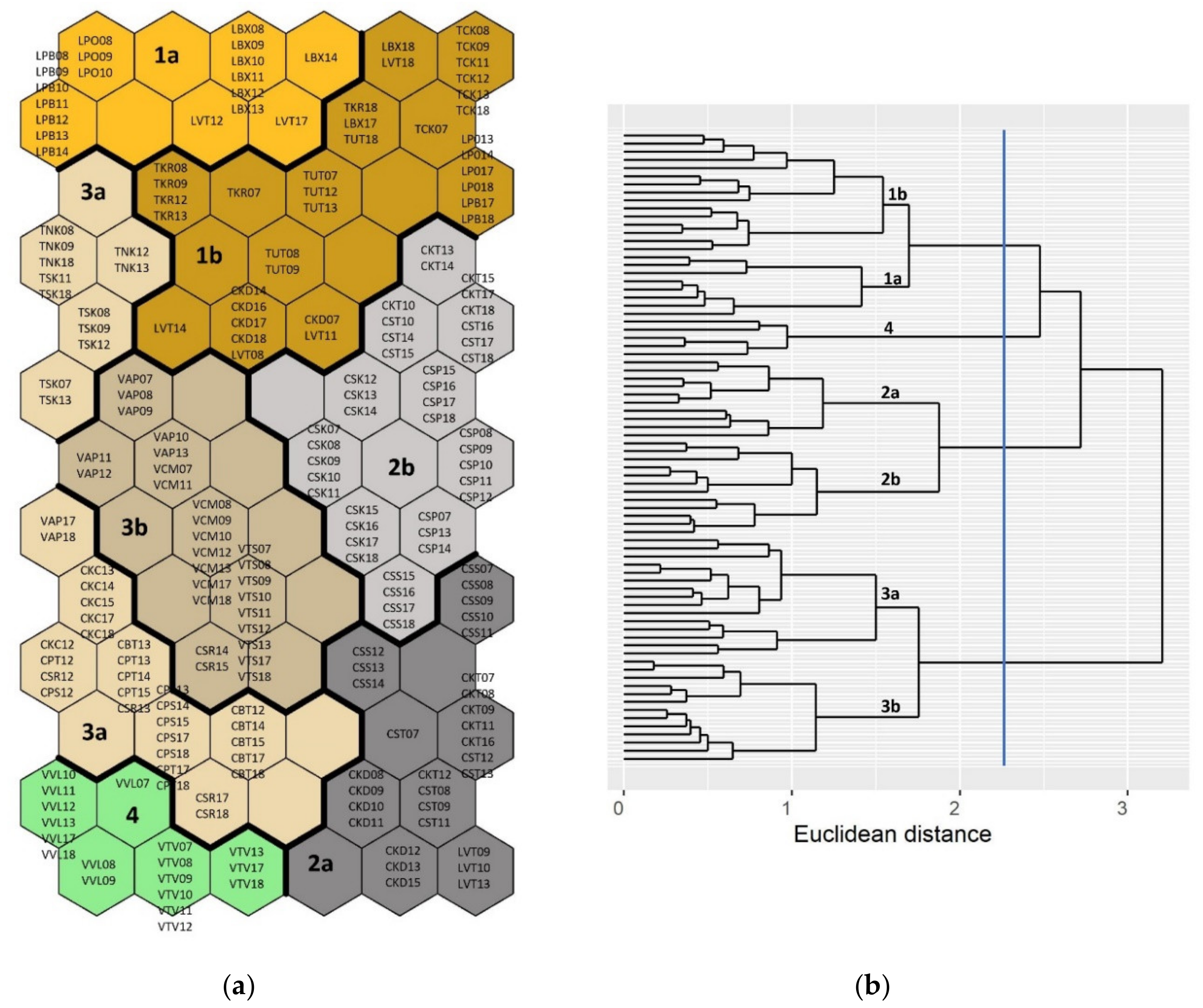
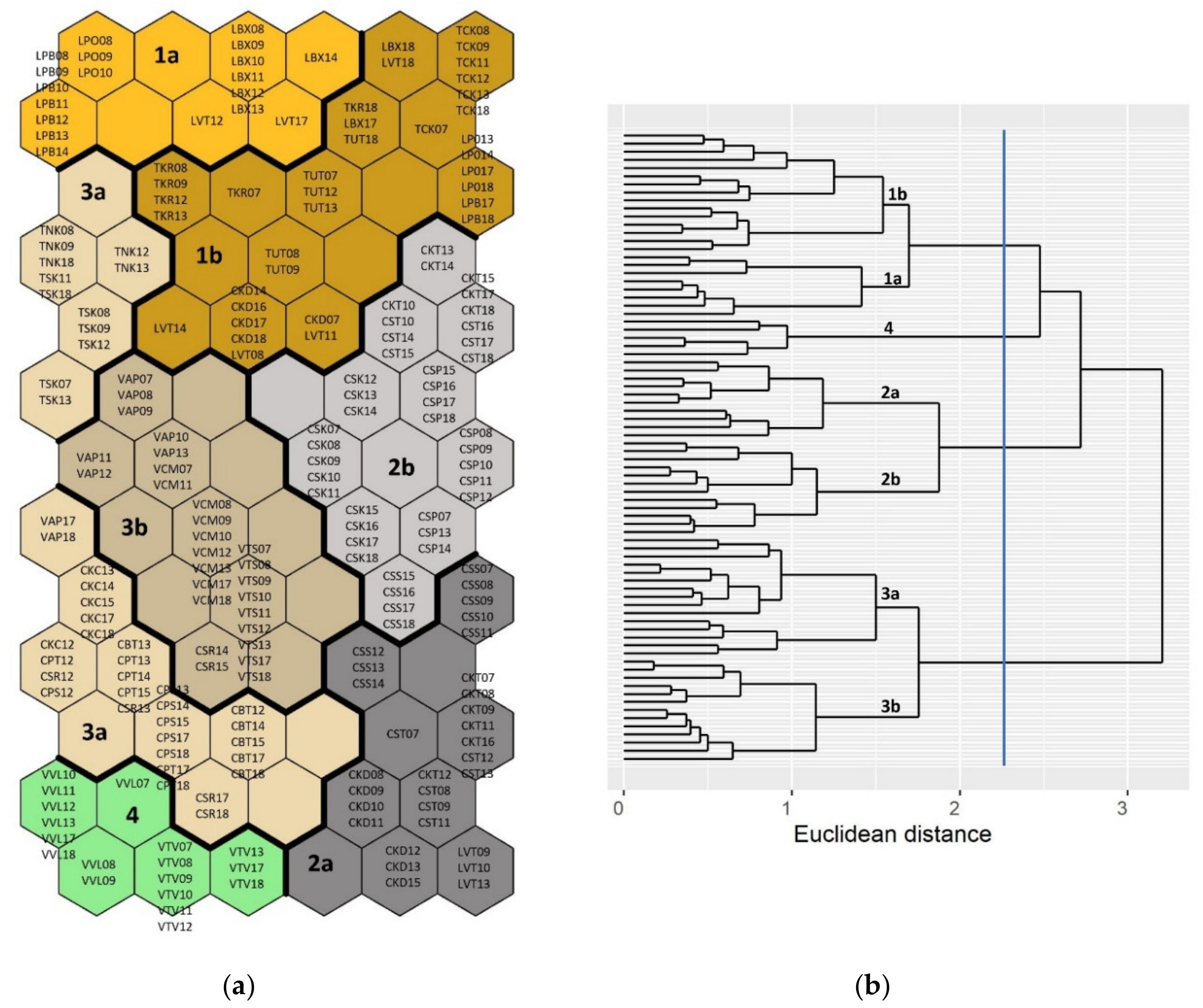
2.3.1. Fish Community Structures and Spatiotemporal Variations
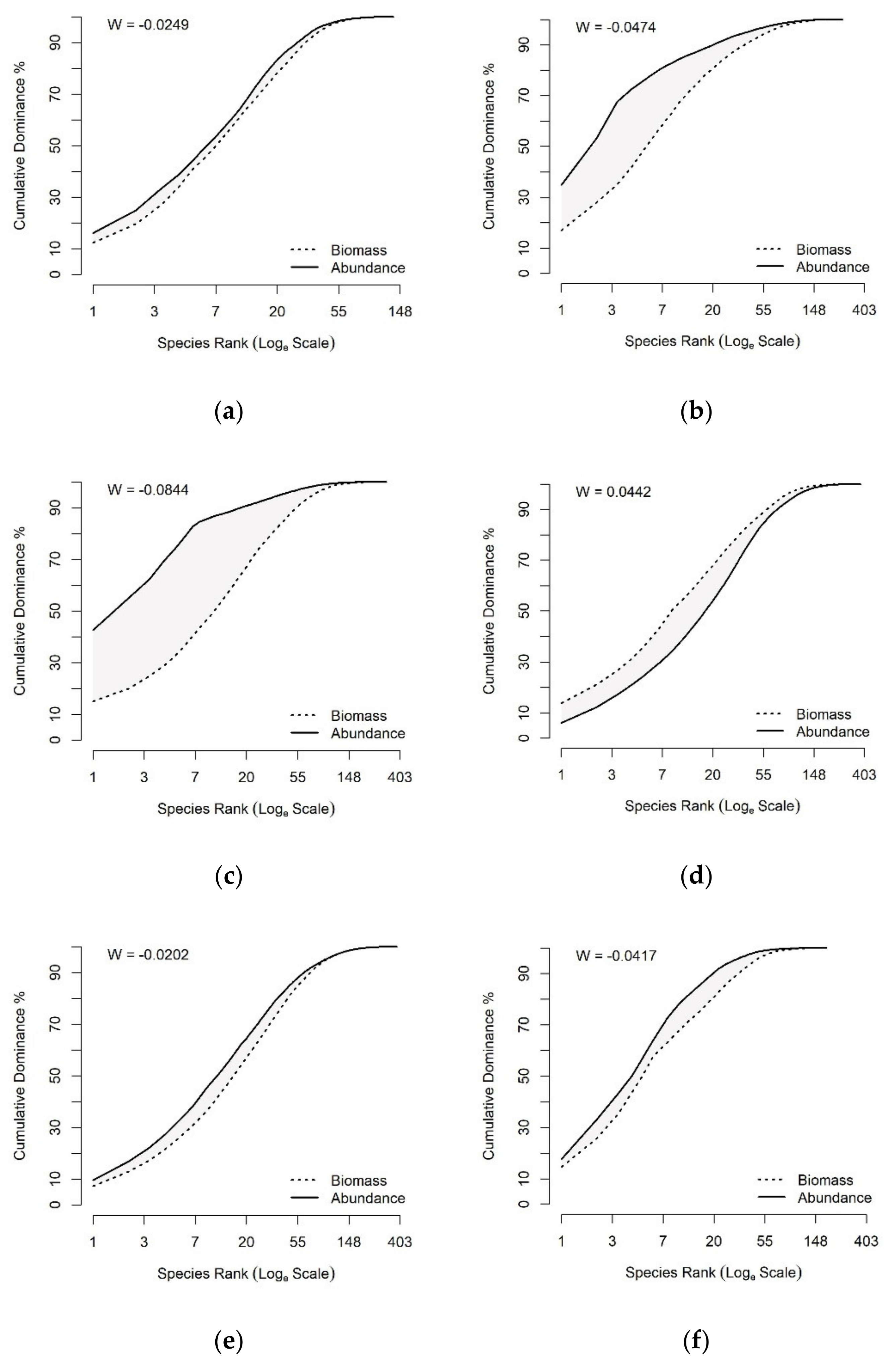
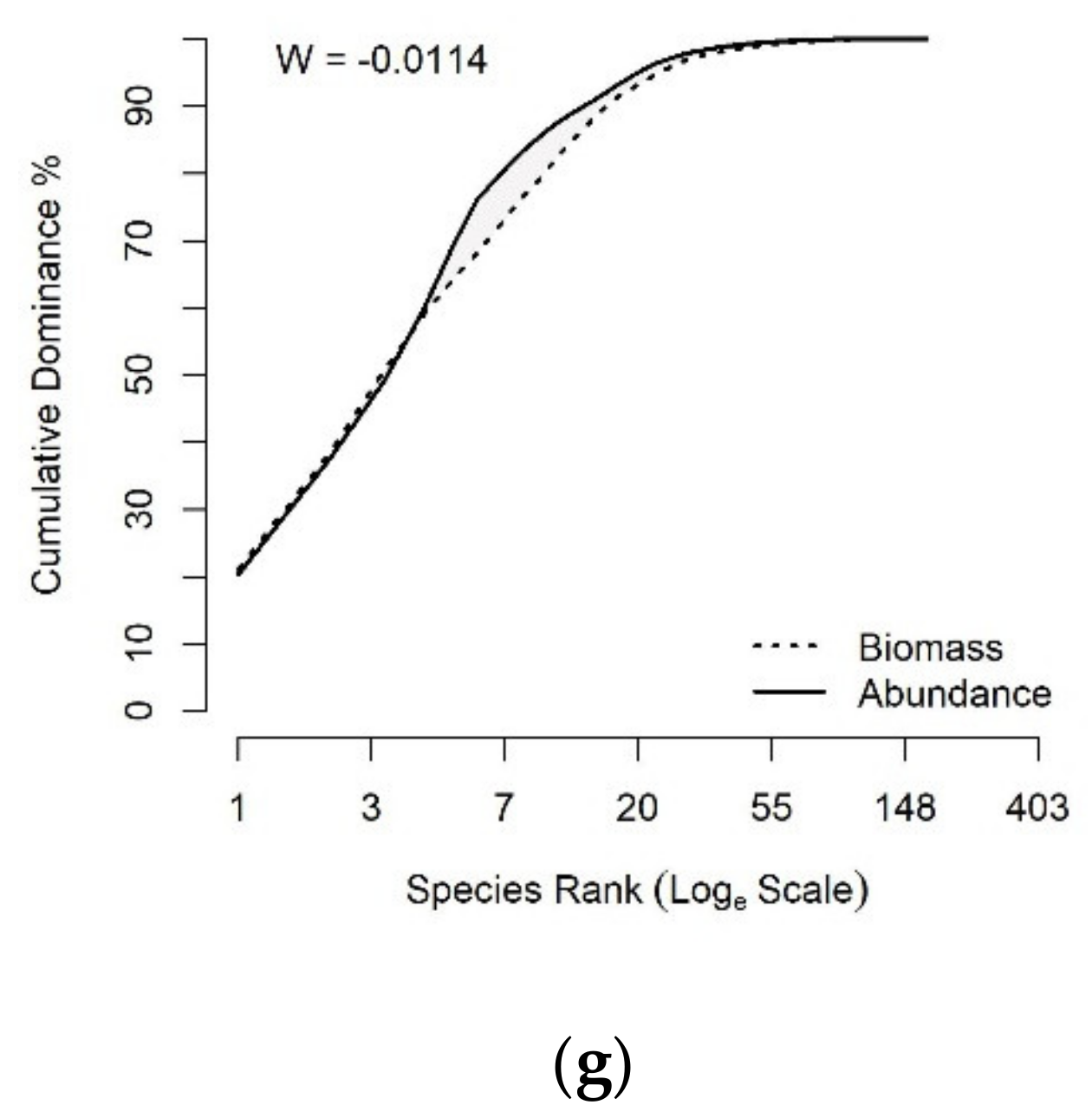
2.3.2. Status of Fish Communities in the LMB
2.3.3. Classification of Fish-Bodied Size
3. Results
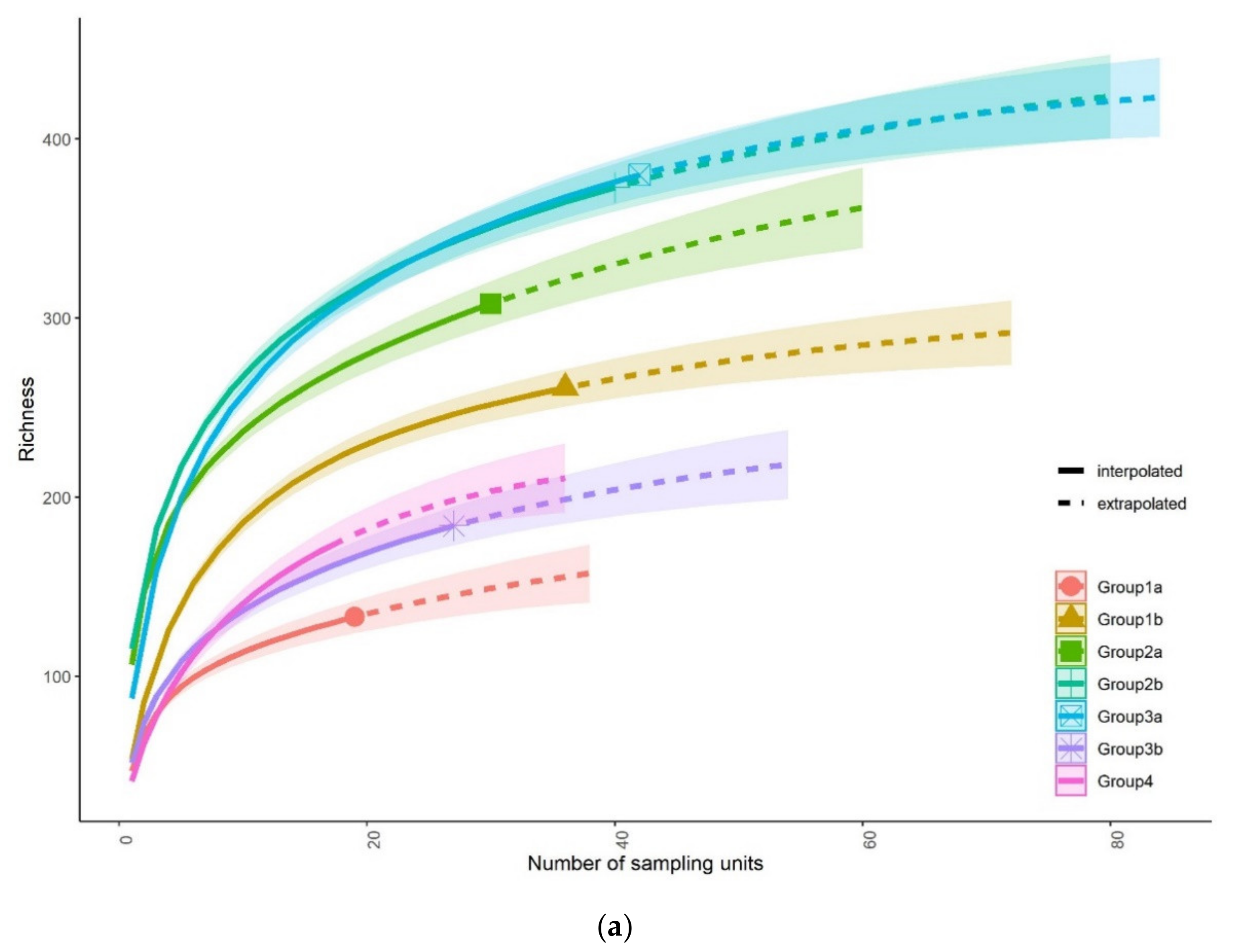
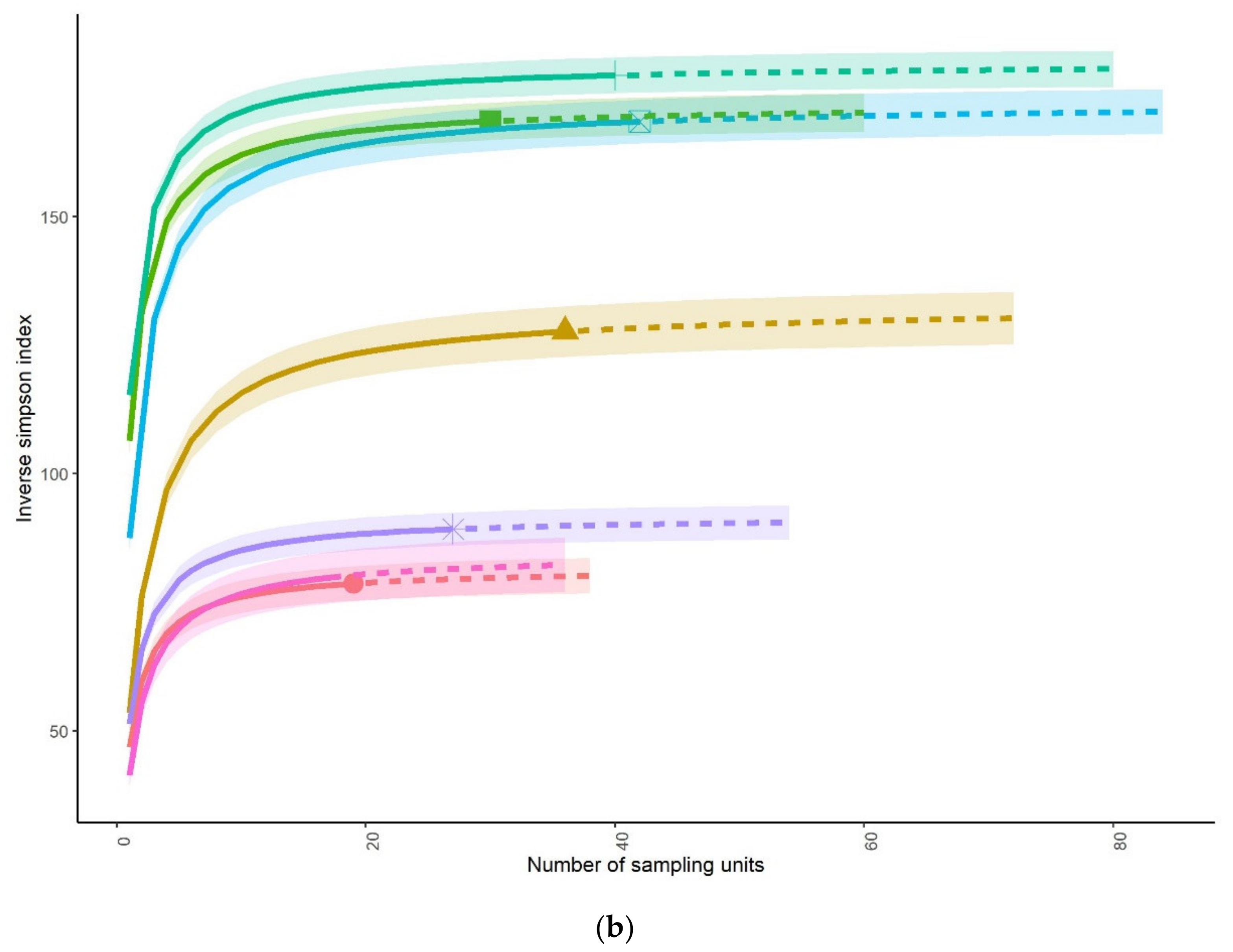
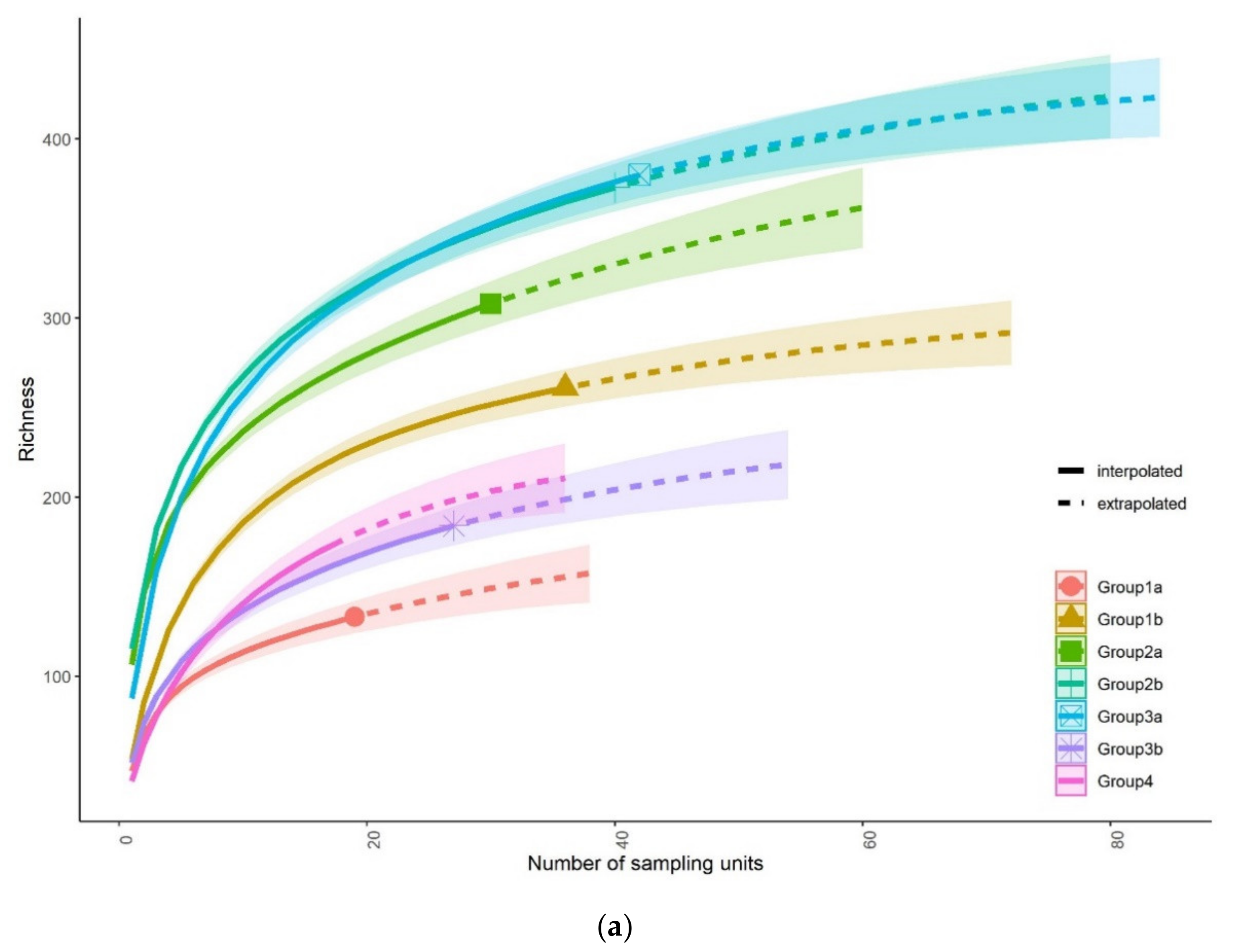
3.1. Fish Community Structures
3.2. Indicator Species by Groups
3.3. Status of Fish Communities in the LMB
4. Discussion
4.1. Fish Diversity in the LMB
4.2. Fish Community Structure and Its Ecological Status
5. Conclusions and Recommendation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benejam, L.; Saura-Mas, S.; Bardina, M.; Solà, C.; Munné, A.; García-Berthou, E. Ecological impacts of small hydropower plants on headwater stream fish: From individual to community effects. Ecol. Freshw. Fish 2016, 25, 295–306. [Google Scholar] [CrossRef]
- Gomes-Silva, G.; Cyubahiro, E.; Wronski, T.; Riesch, R.; Apio, A.; Plath, M. Water pollution affects fish community structure and alters evolutionary trajectories of invasive guppies (Poecilia reticulata). Sci. Total Environ. 2020, 730, 138912. [Google Scholar] [CrossRef] [PubMed]
- McKay, S.F.; King, A.J. Potential ecological effects of water extraction in small, unregulated streams. River Res. Appl. 2006, 22, 1023–1037. [Google Scholar] [CrossRef]
- Ngor, P.B.; McCann, K.S.; Grenouillet, G.; So, N.; McMeans, B.C.; Fraser, E.; Lek, S. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, K.S.; Gellner, G.; McMeans, B.C.; Deenik, T.; Holtgrieve, G.; Rooney, N.; Hannah, L.; Cooperman, M.; Nam, S. Food webs and the sustainability of indiscriminate fisheries. Can. J. Fish. Aquat. Sci. 2016, 73, 656–665. [Google Scholar] [CrossRef]
- Jellyman, P.G.; Harding, J.S. The role of dams in altering freshwater fish communities in New Zealand. N. Zeal. J. Mar. Freshw. Res. 2012, 46, 475–489. [Google Scholar] [CrossRef] [Green Version]
- Oeurng, C.; Sok, T. Assessing changes in flow and water quality emerging from hydropower development and operation in the Sesan River Basin of the Lower Mekong Region. Sustain. Water Resour. Manag. 2020, 6, 1–12. [Google Scholar] [CrossRef]
- Mekong River Commission. The Council Study: The Study on Sustainable Management and Development of the Mekong River, Including Impacts of Mainstream Hydropower Projects. Biological Resource Assessment Final Technical Report Series. Volume 1: Specialists’ Report; Mekong River Commission Secretariat: Vientiane, Laos, 2017; Volume 1. [Google Scholar]
- Penczak, T.; Kruk, A. Applicability of the abundance/biomass comparison method for detecting human impacts on fish populations in the Pilica River, Poland. Fish. Res. 1999, 39, 229–240. [Google Scholar] [CrossRef]
- Hu, M.; Wang, C.; Liu, Y.; Zhang, X.; Jian, S. Fish species composition, distribution and community structure in the lower reaches of Ganjiang River, Jiangxi, China. Sci. Rep. 2019, 9, 10100. [Google Scholar] [CrossRef] [Green Version]
- Kundu, G.K.; Islam, M.M.; Hasan, M.F.; Saha, S.; Mondal, G.; Paul, B.; Mustafa, M.G. Patterns of fish community composition and biodiversity in riverine fish sanctuaries in Bangladesh: Implications for hilsa shad conservation. Ecol. Freshw. Fish 2020, 29, 364–376. [Google Scholar] [CrossRef]
- Stojković, M.; Milošević, D.; Simić, S.; Simić, V. Using a Fish-Based Model to Assess the Ecological Status of Lotic Systems in Serbia. Water Resour. Manag. 2014, 28, 4615–4629. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Wadable Streams and Rivers: Periphyton, Benthic Macroinvertebrates, and Fish, 2nd ed.; U.S. Environmental Protection Agency: Washington, DC, USA, 1999.
- Huang, J.; Huang, L.; Wu, Z.; Mo, Y.; Zou, Q.; Wu, N.; Chen, Z. Correlation of fish assemblages with habitat and environmental variables in a headwater stream section of Lijiang River, China. Sustainability 2019, 11, 1135. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Chakrabarty, D. The use of fish community structure as a measure of ecological degradation: A case study in two tropical rivers of India. BioSystems 2007, 90, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.C. Introduction. In The Mekong Biophysical Environment of an International River Basin; Campbell, I.C., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2009; ISBN 978-0-12-374026-7. [Google Scholar]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots; Zachos, F., Habel, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. ISBN 978-3-642-20991-8. [Google Scholar]
- Ng, T.H.; Jeratthitikul, E.; Sutcharit, C.; Chhuoy, S.; Pin, K.; Pholyotha, A.; Siriwut, W.; Srisonchai, R.; Hogan, Z.S.; Ngor, P.B. Annotated checklist of freshwater molluscs from the largest freshwater lake in Southeast Asia. Zookeys 2020, 2020, 107–141. [Google Scholar] [CrossRef]
- Sor, R.; Ngor, P.B.; Boets, P.; Goethals, P.L.M.; Lek, S.; Hogan, Z.S.; Park, Y. Patterns of Mekong Mollusc Biodiversity: Identification of Emerging Threats and Importance to Management and Livelihoods in a Region of Globally Significant Biodiversity and Endemism. Water 2020, 12, 2619. [Google Scholar] [CrossRef]
- Ngor, P.B.; Sor, R.; Prak, L.H.; So, N.; Hogan, Z.S.; Lek, S. Mollusc fisheries and length-weight relationship in Tonle Sap flood pulse system, Cambodia. Ann. Limnol. 2018, 54. [Google Scholar] [CrossRef]
- Mekong River Commission. State of the Basin Report 2018; Mekong River Commission Secretariat: Vientiane, Laos, 2018. [Google Scholar]
- Jézéquel, C.; Tedesco, P.A.; Bigorne, R.; Maldonado-Ocampo, J.A.; Ortega, H.; Hidalgo, M.; Martens, K.; Torrente-Vilara, G.; Zuanon, J.; Acosta, A.; et al. A database of freshwater fish species of the Amazon Basin. Sci. Data 2020, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Harrison, I.J.; Brummett, R.; Stiassny, M.L.J. Congo River Basin. In The Wetland Book; Finlayson, C., Milton, G., Prentice, R., Davidson, N., Eds.; Springer: Dordrecht, The Netherlands, 2018. [Google Scholar]
- Mekong River Commission. Report on the 2015 Biomonitoring Survey of the Lower Mekong River and Selected Tributaries; Mekong River Commission Secretariat: Vientiane, Laos, 2019. [Google Scholar]
- Bravard, J.-P.; Goichot, M.; Gaillot, S. Geography of Sand and Gravel Mining in the Lower Mekong River. EchoGéo 2013, 26. [Google Scholar] [CrossRef] [Green Version]
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of Inland Waters. Bioscience 2005, 55, 1041. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.E.; Holtgrieve, G.W.; Ngor, P.B.; Dang, T.D.; Piman, T. Maintaining perspective of ongoing environmental change in the Mekong floodplains. Curr. Opin. Environ. Sustain. 2019, 37, 1–7. [Google Scholar] [CrossRef]
- Xie, J.Y.; Tang, W.J.; Yang, Y.H. Fish assemblage changes over half a century in the Yellow River, China. Ecol. Evol. 2018, 8, 4173–4182. [Google Scholar] [CrossRef] [PubMed]
- Yemane, D.; Field, J.G.; Leslie, R.W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves. ICES J. Mar. Sci. 2005, 62, 374–379. [Google Scholar] [CrossRef] [Green Version]
- Chea, R.; Lek, S.; Ngor, P.; Grenouillet, G. Large-scale patterns of fish diversity and assemblage structure in the longest tropical river in Asia. Ecol. Freshw. Fish 2017, 26, 575–585. [Google Scholar] [CrossRef]
- Mekong River Commission. Overview of the Hydrology of the Mekong Basin; Mekong River Commission: Phnom Penh, Cambodia, 2005. [Google Scholar]
- Mekong River Commission. Monitoring Fish Abundance and Diversity in the Lower Mekong Basin: Methodological Guidelines; Mekong River Commission: Phnom Penh, Cambodia, 2007. [Google Scholar]
- Ngor, P.B.; Hortle, K.G.; So, N. Standard Sampling Procedures for Fish Abundance and Diversity Monitoring in the Lower Mekong Basin; Mekong River Commission Secretariat: Phnom Penh, Cambodia, 2016. [Google Scholar]
- Kohonen, T. MATLAB Implementations and Applications of the Self-Organizing Map; Unigrafia Oy: Helsinki, Finland, 2014; ISBN 9789526036786. [Google Scholar]
- Brosse, S.; Giraudel, J.L.; Lek, S. Utilisation of non-supervised neural networks and principal component analysis to study fish assemblages. Ecol. Modell. 2001, 146, 159–166. [Google Scholar] [CrossRef]
- Giraudel, J.L.; Lek, S. A comparison of self-organizing map algorithm and some conventional statistical methods for ecological community ordination. Ecol. Modell. 2001, 146, 329–339. [Google Scholar] [CrossRef]
- Park, S.H.; Hosoishi, S.; Ogata, K.; Kuboki, Y. Clustering of ant communities and indicator species analysis using self-organizing maps. Comptes Rendus Biol. 2014, 337, 545–552. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organazing Maps, 3rd ed.; Springer: Berlin, Germany, 2001; ISBN 9783540679219. [Google Scholar]
- Vesanto, J. SOM Implementation in SOM Toolbox. SOM Toolbox Online Help. Available online: http://www.cis.hut.fi/projects/somtoolbox/documentation/somalg.shtml (accessed on 1 January 2020).
- Wehrens, R.; Buydens, L.M.C. Self- and super-organizing maps in R: The kohonen package. J. Stat. Softw. 2007, 21, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, J.A.; Reynolds, J.F. Statistical Ecology, a Primer on Methods and Computing; John Wiley & Sons: Hoboken, NJ, USA, 1988. [Google Scholar]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Cox, K.D.; Black, M.J.; Filip, N.; Miller, M.R.; Mohns, K.; Mortimor, J.; Freitas, T.R.; Greiter Loerzer, R.; Gerwing, T.G.; Juanes, F.; et al. Community assessment techniques and the implications for rarefaction and extrapolation with Hill numbers. Ecol. Evol. 2017, 7, 11213–11226. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Bauman, D.; Blanchet, F.G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate multiscale spatial analysis. Cran. 2020. Available online: https://cran.r-project.org/web/packages/adespatial/index.html (accessed on 1 January 2020).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Warwick, R.M. A new method for detecting pollution effect on macrobenthic communitries. Mar. Biol. 1986, 92, 557–562. [Google Scholar] [CrossRef]
- Coeck, J.; Vandelannoote, A.; Yseboodt, R.; Verheyen, R.F. Use of the abundance/biomass method for comparison of fish communities in regulated and unregulated lowland rivers in Belgium. Regul. Rivers Res. Manag. 1993, 8, 73–82. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.; Sommerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Aluizio, R. Foraminifera and Community Ecology Analyses. 2015. Available online: https://rdrr.io/cran/forams/ (accessed on 1 January 2020).
- Ngor, P.B.; Oberdorff, T.; Phen, C.; Baehr, C.; Grenouillet, G.; Lek, S. Fish assemblage responses to flow seasonality and predictability in a tropical flood pulse system. Ecosphere 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- MFD. Mekong Fish Database: A Taxonomic Fish Database for the Mekong Basin; MFD: Phnom Penh, Cambodia, 2003. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 1 January 2020).
- Ngor, P.B.; Legendre, P.; Oberdorff, T.; Lek, S. Flow alterations by dams shaped fish assemblage dynamics in the complex Mekong-3S river system. Ecol. Indic. 2018, 88, 103–114. [Google Scholar] [CrossRef]
- Ngor, P.B.; Grenouillet, G.; Phem, S.; So, N.; Lek, S. Spatial and temporal variation in fish community structure and diversity in the largest tropical flood-pulse system of South-East Asia. Ecol. Freshw. Fish 2018, 27, 1087–1100. [Google Scholar] [CrossRef]
- Valbo-Jorgensen, J.; Coates, D.; Hortle, K.G. Fish diversity in the Mekong River Basin. In The Mekong Biophysical Environment of an International River Basin; Cambel, C.I., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2009; pp. 162–185. [Google Scholar]
- Welcomme, R.L. River Fisheries; Technical Paper No. 262; FAO: Rome, Italy, 1985. [Google Scholar]
- Oberdorff, T.; Dias, M.S.; Jézéquel, C.; Albert, J.S.; Arantes, C.C.; Bigorne, R.; Carvajal-Valleros, F.M.; De Wever, A.; Frederico, R.G.; Hidalgo, M.; et al. Unexpected fish diversity gradients in the Amazon basin. Sci. Adv. 2019, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenouillet, G.; Pont, D.; Hérissé, C. Within-basin fish assemblage structure: The relative influence of habitat versus stream spatial position on local species richness. Can. J. Fish. Aquat. Sci. 2004, 61, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Suvarnaraksha, A.; Lek, S.; Lek-Ang, S.; Jutagate, T. Fish diversity and assemblage patterns along the longitudinal gradient of a tropical river in the Indo-Burma hotspot region (Ping-Wang River Basin, Thailand). Hydrobiologia 2012, 694, 153–169. [Google Scholar] [CrossRef]
- Klein, J.P.; Vanderpoorten, A. Bryophytic vegetation in riparian forests: Their use in the ecological assessment of the connectivity between the Rhine and its floodplain (Alsace, France). Glob. Ecol. Biogeogr. Lett. 1997, 6, 257–265. [Google Scholar] [CrossRef]
- Reid, M.A.; Reid, M.C.; Thoms, M.C. Ecological significance of hydrological connectivity for wetland plant communities on a dryland floodplain river, MacIntyre River, Australia. Aquat. Sci. 2016, 78, 139–158. [Google Scholar] [CrossRef]
- Poulsen, A.F.; Ouch, P.; Sinthavong, V.; Ubolratana, S.; Nguyen, T.T. Fish Migrations of the Lower Mekong River Basin: Implications for Development, Planning and Environmental Management; Mekong River Commission Secretariat: Phnom Penh, Cambodia, 2002. [Google Scholar]
- International Finance Corporation. Nam Ou River Basin Profile; International Finance Corporation: Washington, DC, USA, 2017. [Google Scholar]
- Hogan, Z. A Mekong Giant: Current Status, Threats and Preliminary Conservation Measures for the Critically Endangered Mekong Giant Catfish; WWF-Great Mekong: Bangkok, Thailand, 2012. [Google Scholar]
- Kottelat, M. The fishes of the Nam Theun and Xe Bangfai drainages, Laos. Hydroecologie Appl. 2015, 19, 271–320. [Google Scholar] [CrossRef] [Green Version]
- Mekong River Commission. Modelling the Impacts of Climate Change and Development Infrastructure on Mekong Flow, Sediment Regimes and Water Quality; Mekong River Commission Secretariat: Vientiane, Laos, 2018; Volume 1. [Google Scholar]
- Baran, E.; Starr, P.; Kura, Y. Influence of Built Structures on Tonle Sap Fisheries; Cambodia National Mekong Committee and the WordFish Center: Phnom Penh, Cambodia, 2007. [Google Scholar]
- Van Binh, D.; Kantoush, S.; Sumi, T. Changes to long-term discharge and sediment loads in the Vietnamese Mekong Delta caused by upstream dams. Geomorphology 2020, 353, 107011. [Google Scholar] [CrossRef]
- Grumbine, R.E.; Xu, J. Mekong hydropower development. Science 2011, 332, 178–179. [Google Scholar] [CrossRef]
- Kummu, M.; Lu, X.X.; Wang, J.J.; Varis, O. Basin-wide sediment trapping efficiency of emerging reservoirs along the Mekong. Geomorphology 2010, 119, 181–197. [Google Scholar] [CrossRef]
- Kummu, M.; Varis, O. Sediment-related impacts due to upstream reservoir trapping, the Lower Mekong River. Geomorphology 2007, 85, 275–293. [Google Scholar] [CrossRef]
- Mekong River Commission. Thematic Report on the Positive and Negative Impacts of Hydropower Development on the Social, Environmental, and Economic Conditions of the Lower Mekong River Basin; Mekong River Commission Secretariat: Vientiane, Laos, 2017. [Google Scholar]
- Asian Development Bank. Environmental Monitoring Report for Second Quarter of 2018; Asian Development Bank: Mandaluyong, Philippines, 2018. [Google Scholar]
- Baran, E.; Samadee, S.; Jiau, T.S.; Tran, T.C. Fish and Fisheries in the Sesan, Sekong and Srepok River Basins (Mekong Watershed), with Special Reference to the Sesan River; ICEM–International Centre for Environmental Management: Hanoi, Vietnam, 2013. [Google Scholar]
- Benzuijen, M.R.; Timmins, R.; Seng, T. Biological Surveys of the Mekong River between Kratie and Stung Treng Towns, Northeast Cambodia, 2006–2007; WWF-Great Mekong-Cambodia Country Programme, Cambodia Fisheries Administration and Cambodia Forestry Administration: Phnom Penh, Cambodia, 2007. [Google Scholar]
- Montaña, C.G.; Ou, C.; Keppeler, F.W.; Winemiller, K.O. Functional and trophic diversity of fishes in the Mekong-3S river system: Comparison of morphological and isotopic patterns. Environ. Biol. Fishes 2020, 103, 185–200. [Google Scholar] [CrossRef]
- Oliveira, A.G.; Baumgartner, M.T.; Gomes, L.C.; Dias, R.M.; Agostinho, A.A. Long-term effects of flow regulation by dams simplify fish functional diversity. Freshw. Biol. 2018, 63, 293–305. [Google Scholar] [CrossRef]
- Mekong River Commission. Planning Atlas of the Lower Mekong River Basin; Mekong River Commission Secretariat: Vientiane, Laos, 2011. [Google Scholar]
- Anticamara, J.A.; Zeller, D.; Vincent, A.C.J. Spatial and temporal variation of abundance, biomass and diversity within marine reserves in the Philippines. Divers. Distrib. 2010, 16, 529–536. [Google Scholar] [CrossRef]
- Constable, D. Atlas of the 3S Basins; IUCN: Bangkok, Thailand, 2015. [Google Scholar]
- Lo, M.; Reed, J.; Castello, L.; Steel, E.A.; Frimpong, E.A.; Ickowitz, A. The Influence of Forests on Freshwater Fish in the Tropics: A Systematic Review. Bioscience 2020, 70, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres, F. Fishing down marine food webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef]
- Nuon, V.; Gallardob, W. Perceptions of the local community on the outcome of community fishery management in Krala Peah village, Cambodia. Int. J. Sustain. Dev. World Ecol. 2011, 18. [Google Scholar] [CrossRef]
- Ounboundisane, S.; Loury, E.; Chamsinh, B.; Guegan, F. Status Report on Fish Conservation Zones in Laos; FISHBIO: Vientiane, Laos, 2019. [Google Scholar]
- Mekong River Commission. Transboundary Fisheries Management Issues in the Mekong and Sekong Rivers of Cambodia and Laos; Mekong River Commission Secretariat: Vientiane, Laos, 2017. [Google Scholar]
- Pawaputanon, O.; Mahasarakarm, N. An introduction to the Mekong Fisheries of Thailand; Mekong River Commission Secretariat: Vientiane, Laos, 2007. [Google Scholar]
- Dieu, V.L. Electro fishing practices in the Srepok River Basin in central highlands, Vietnam. In Proceedings of the 10th Technical Symposium on the Mekong Fisheries, Pakse, Laos, 11–13 November 2009; Mekong River Commission Secretariat: Vientiane, Laos, 2009. [Google Scholar]
- Campbell, T.; Pin, K.; Ngor, P.B.; Hogan, Z. Conserving mekong megafishes: Current status and critical threats in Cambodia. Water 2020, 12. [Google Scholar] [CrossRef]
- Chan, B.; Ngor, P.B.; So, N.; Lek, S. Spatial and temporal changes in fish yields and fish communities in the largest tropical floodplain lake in Asia. Ann. Limnol. 2017, 53, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Rainboth, W. FAO Species Identification Field Guide for Fishery Purposes. Fishes of the Cambodia Mekong; FAO: Rome, Italy, 1996. [Google Scholar]
- Thai Ban Research Network. Fish Species in the “Pa Bung Pa Thamm”–Local Knowledge of Fishers in the Lower Songkhram River Basin; Sustainable Biodiversity Use and Conservation in the Mekong Basin Wetlands Project, Nakhon Phanom Conservation Association and the Lower Songkhram Basin Tai Ban Research Network: Bangkok, Thailand, 2005. [Google Scholar]
- An, V.V.; Tien, V.D.; Ngor, P.B.; Nguyen, H.S.; So, N. Exotic species in southern Viet Nam. Catch Cult. 2013, 19, 1–48. [Google Scholar]
- Young, A. Regional Irrigation Sector Review for Joint Basin Planning Process, Regional Irrigation Sector Review for Joint Basin Planning Process; Mekong River Commission Secretariat: Vientiane, Laos, 2009. [Google Scholar]
- Malherbe, W.; Wepener, V.; Van Vuren, J.H.J. The effect of a large-scale irrigation scheme on the fish community structure and integrity of a subtropical river system in South Africa. Ecol. Indic. 2016, 69, 533–539. [Google Scholar] [CrossRef]
- Olson, K.R.; Wright Morton, L. Polders, dikes, canals, rice, and aquaculture in the Mekong Delta. J. Soil Water Conserv. 2018, 73, 83A–89A. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, L.J. Population estimation methods to quantify temporal variation in fish accumulations downstream of a weir. Fish. Manag. Ecol. 2006, 13, 355–364. [Google Scholar] [CrossRef]
- Piman, T.; Shrestha, M. Case Study on Sediment in the Mekong River Basin: Current State and Future Trends; Sotckholm Environment Institute: Stockholm, Sweden, 2017. [Google Scholar]
- Jordan, C.; Tiede, J.; Lojek, O.; Visscher, J.; Apel, H.; Nguyen, H.Q.; Quang, C.N.X.; Schlurmann, T. Sand mining in the Mekong Delta revisited-current scales of local sediment deficits. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tonle Sap Authority. EIA on Mineral Resources around Tonle Sap Lake and Related Areas; Authority Tonle Sap: Phnom Penh, Cambodia, 2019. [Google Scholar]
- Hackney, C.R.; Darby, S.E.; Parsons, D.R.; Leyland, J.; Best, J.L.; Aalto, R.; Nicholas, A.P.; Houseago, R.C. River bank instability from unsustainable sand mining in the lower Mekong River. Nat. Sustain. 2020, 3, 217–225. [Google Scholar] [CrossRef]
- Pitchaiah, P.S. Impacts of Sand Mining on Environment—A Review. Int. J. Geoinformatics Geol. Sci. 2017, 4, 1–6. [Google Scholar] [CrossRef]
- Duan, X.; Wang, Z.; Tian, S. Effect of streambed substrate on macroinvertebrate biodiversity. Front. Environ. Sci. Eng. China 2008, 2, 122–128. [Google Scholar] [CrossRef]
- Boyd, S.E.; Limpenny, D.S.; Rees, H.L.; Cooper, K.M. The effects of marine sand and gravel extraction on the macrobenthos at a commercial dredging site (results 6 years post-dredging). ICES J. Mar. Sci. 2005, 62, 145–162. [Google Scholar] [CrossRef] [Green Version]
- Diehl, S.; Kornijów, R. Influence of Submerged Macrophytes on Trophic Interactions Among Fish and Macroinvertebrates. In The Structuring Role of Submerged Macrophytes in Lakes; Jeppesen, E., Sondergaard, M., Sondergaard, M., Christoffersen, K., Eds.; Springer: New York, NY, USA, 1998; pp. 24–46. [Google Scholar]
- Tonle Sap Authority. Technical Report on an Update of Fishing Gears in the Six Provinces around Tonle Sap Lake; Tonle Sap Authority: Phnom Penh, Cambodia, 2018. [Google Scholar]
- Sovannara, H. Annual Report on Fisher Catch Monitoring in the Provinces around Tonle Sap Lake, Cambodia; Tonle Sap Authority: Phnom Penh, Cambodia, 2020. [Google Scholar]
- Kc, K.B.; Bond, N.; Fraser, E.D.G.; Elliott, V.; Farrell, T.; McCann, K.; Rooney, N.; Bieg, C. Exploring tropical fisheries through fishers’ perceptions: Fishing down the food web in the Tonlé Sap, Cambodia. Fish. Manag. Ecol. 2017, 24, 452–459. [Google Scholar] [CrossRef]
- Johnstone, G.; Ranjtha, P.; Fabrice, D.; Kosal, M.; Oeur, I.; Sithirith, M.; Bunna, P.; Sophat, S.; Sokheng, C.; Sochanny, H.; et al. Tonle Sap Scoping Report; CGIAR Research Program on Aquatic Agricultural Systems: Penang, Malaysia, 2013. [Google Scholar]
- Torell, A.; Salamanca, A. Wetlands Management in Vietnam’s Mekong Delta: An Overview of Pressures and Responses. In Wetlands Management in Vietnam: Issues and Perspectives; Torell, M., Salamanca, A.M., Ratner, B.D., Eds.; WorldFish: Penang, Malaysia, 2003; pp. 1–19. [Google Scholar]
- Vu, A.; Baumgartner, L.J.; Howitt, J.; Limburg, K.; Reinhardt, J.; Mallen-Cooper, M.; Cowx, I. Connectivity of Mekong fishes between freshwater and marine habitats. Catch Cult. Environ. 2020, 26, 1–48. [Google Scholar]
- Hogan, Z.; Baird, I.G.; Radtke, R.; Vander Zanden, M.J. Long distance migration and marine habitation in the tropical Asian catfish, Pangasius krempfi. J. Fish Biol. 2007, 71, 818–832. [Google Scholar] [CrossRef]
- Tran, N.T.; Labonne, M.; Hoang, H.D.; Panfili, J. Changes in environmental salinity during the life of Pangasius krempfi in the Mekong Delta (Vietnam) estimated from otolith Sr: Ca ratios. Mar. Freshw. Res. 2019, 70, 1734–1746. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.T.C.; Avtar, R.; Fujii, M. Monitoring changes in land use and distribution of mangroves in the southeastern part of the Mekong River Delta, Vietnam. Trop. Ecol. 2019, 60, 552–565. [Google Scholar] [CrossRef]
- Laegdsgaard, P.; Johnson, C. Why do juvenile fish utilise mangrove habitats? J. Exp. Mar. Bio. Ecol. 2001, 257, 229–253. [Google Scholar] [CrossRef] [Green Version]
- Wenger, A.S.; Harvey, E.; Wilson, S.; Rawson, C.; Newman, S.J.; Clarke, D.; Saunders, B.J.; Browne, N.; Travers, M.J.; Mcilwain, J.L.; et al. A critical analysis of the direct effects of dredging on fish. Fish Fish. 2017, 18, 967–985. [Google Scholar] [CrossRef]
- Mingist, M.; Gebremedhin, S. Could sand mining be a major threat for the declining endemic Labeobarbus species of Lake Tana, Ethiopia? Singap. J. Trop. Geogr. 2016, 37, 195–208. [Google Scholar] [CrossRef]
- Koehnken, L.; Rintoul, M.S.; Goichot, M.; Tickner, D.; Loftus, A.C.; Acreman, M.C. Impacts of riverine sand mining on freshwater ecosystems: A review of the scientific evidence and guidance for future research. River Res. Appl. 2020, 36, 362–370. [Google Scholar] [CrossRef]
- Shao, X.; Fang, Y.; Jawitz, J.W.; Yan, J.; Cui, B. River network connectivity and fish diversity. Sci. Total Environ. 2019, 689, 21–30. [Google Scholar] [CrossRef]
- Wilkinson, C.L.; Yeo, D.C.J.; Tan, H.H.; Fikri, A.H.; Ewers, R.M. Land-use change is associated with a significant loss of freshwater fish species and functional richness in Sabah, Malaysia. Biol. Conserv. 2018, 222, 164–171. [Google Scholar] [CrossRef]
- Jones, E.B.D.; Helfman, G.S.; Harper, J.O.; Bolstad, P.V. Effects of riparian forest removal on fish assemblages in southern Appalachian streams. Conserv. Biol. 1999, 13, 1454–1465. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Scientific Name | Category |
|---|---|---|---|
| Cypriniformes | Cyprinidae | Aaptosyax grypus | CR |
| Cypriniformes | Cyprinidae | Catlocarpio siamensis | CR |
| Siluriformes | Pangasiidae | Pangasianodon gigas | CR |
| Siluriformes | Pangasiidae | Pangasius sanitwongsei | CR |
| Cypriniformes | Cyprinidae | Poropuntius deauratus | EN |
| Siluriformes | Pangasiidae | Pangasianodon hypophthalmus | EN |
| Cypriniformes | Cyprinidae | Probarbus jullieni | EN |
| Cypriniformes | Cyprinidae | Probarbus labeamajor | EN |
| Osteoglossiformes | Osteoglossidae | Scleropages formosus | EN |
| Cypriniformes | Cyprinidae | Luciocyprinus striolatus | EN |
| Cypriniformes | Cyprinidae | Poropuntius solitus | EN |
| Myliobatiformes | Dasyatidae | Dasyatis laosensis | EN |
| Cypriniformes | Cyprinidae | Poropuntius bolovenensis | EN |
| Myliobatiformes | Dasyatidae | Himantura oxyrhynchus | EN |
| Cypriniformes | Cyprinidae | Poropuntius consternans | EN |
| Myliobatiformes | Dasyatidae | Himantura signifer | EN |
| Cypriniformes | Cyprinidae | Hypsibarbus lagleri | VU |
| Siluriformes | Pangasiidae | Pangasius krempfi | VU |
| Cypriniformes | Cyprinidae | Cirrhinus microlepis | VU |
| Cypriniformes | Cyprinidae | Scaphognathops bandanensis | VU |
| Perciformes | Osphronemidae | Osphronemus exodon | VU |
| Perciformes | Datnioididae | Datnioides undecimradiatus | VU |
| Cypriniformes | Cyprinidae | Labeo pierrei | VU |
| Cypriniformes | Cyprinidae | Bangana behri | VU |
| Clupeiformes | Clupeidae | Tenualosa thibaudeaui | VU |
| Cypriniformes | Cyprinidae | Epalzeorhynchos munense | VU |
| Cypriniformes | Cyprinidae | Poropuntius speleops | VU |
| Cypriniformes | Cobitidae | Botia nigrolineata | VU |
| Cypriniformes | Cobitidae | Serpenticobitis cingulata | VU |
| Cypriniformes | Cyprinidae | Tor ater | VU |
| Cypriniformes | Cyprinidae | Mystacoleucus lepturus | VU |
| Cypriniformes | Balitoridae | Hemimyzon confluens | VU |
| Cypriniformes | Cyprinidae | Oxygaster pointoni | VU |
| Cypriniformes | Cyprinidae | Troglocyclocheilus khammouanensis | VU |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuon, V.; Lek, S.; Ngor, P.B.; So, N.; Grenouillet, G. Fish Community Responses to Human-Induced Stresses in the Lower Mekong Basin. Water 2020, 12, 3522. https://doi.org/10.3390/w12123522
Nuon V, Lek S, Ngor PB, So N, Grenouillet G. Fish Community Responses to Human-Induced Stresses in the Lower Mekong Basin. Water. 2020; 12(12):3522. https://doi.org/10.3390/w12123522
Chicago/Turabian StyleNuon, Vanna, Sovan Lek, Peng Bun Ngor, Nam So, and Gaël Grenouillet. 2020. "Fish Community Responses to Human-Induced Stresses in the Lower Mekong Basin" Water 12, no. 12: 3522. https://doi.org/10.3390/w12123522
APA StyleNuon, V., Lek, S., Ngor, P. B., So, N., & Grenouillet, G. (2020). Fish Community Responses to Human-Induced Stresses in the Lower Mekong Basin. Water, 12(12), 3522. https://doi.org/10.3390/w12123522