Ecological Connectivity in Two Ancient Lakes: Impact Upon Planktonic Cyanobacteria and Water Quality


 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Work—Microscopy Analysis
2.3. Data Analysis
2.4. Water Quality Assessment
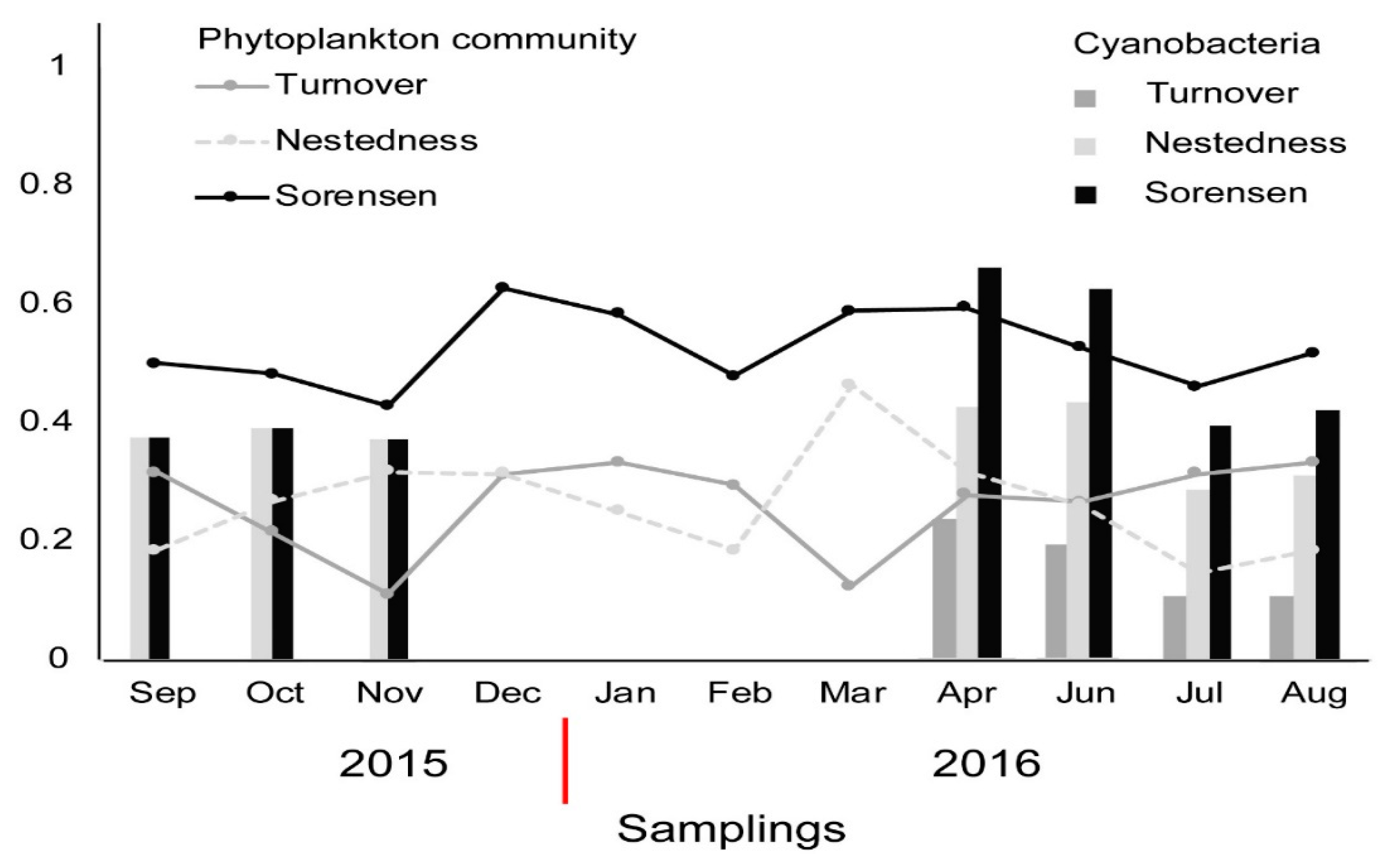
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lampert, W.; Sommer, U. The Ecology of Lakes and Streams, 2nd ed.; Oxford University Press: Oxford, UK, 2007; p. 32. [Google Scholar]
- Chrisostomou, A.; Moustaka-Gouni, M.; Sgardelis, S.; Lanaras, T. Air-dispersed phytoplankton in a Mediterranean river-reservoir system (Aliakmon-Polyphytos, Greece). J. Plankton Res. 2009, 31, 877–884. [Google Scholar] [CrossRef]
- Monchamp, M.-E.; Spaak, P.; Pomati, F. High dispersal levels and lake warming are emergent drivers of cyanobacterial community assembly in peri-Alpine lakes. Sci. Rep. 2019, 9, 7366. [Google Scholar] [CrossRef] [PubMed]
- De Meester, L.; Gómez, A.; Okamura, B.; Schwenk, K. The Monopolization Hypothesis and the dispersal-gene flow paradox in aquatic organisms. Acta Oecol. 2002, 23, 121–135. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef]
- Sukenik, A.; Quesada, A.; Salmaso, N. Global expansion of toxic and non-toxic cyanobacteria: Effect on ecosystem functioning. Biodivers. Conserv. 2015, 24, 889–908. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Sommer, U.; Katsiapi, M.; Vardaka, E. Monitoring of Cyanobacteria for Water Quality: Doing the Necessary Right or Wrong? Available online: http://www.publish.csiro.au/mf/MF18381 (accessed on 18 December 2019).
- Muhl, R.M.W.; Roelke, D.L.; Zohary, T.; Moustaka-Gouni, M.; Sommer, U.; Borics, G.; Gaedke, U.; Withrow, F.G.; Bhattacharyya, J. Resisting annihilation: Relationships between functional trait dissimilarity, assemblage competitive power and allelopathy. Ecol. Lett. 2018, 21, 1390–1400. [Google Scholar] [CrossRef]
- Maloufi, S.; Catherine, A.; Mouillot, D.; Louvard, C.; Couté, A.; Bernard, C.; Troussellier, M. Environmental heterogeneity among lakes promotes hyper β-diversity across phytoplankton communities. Freshw. Biol. 2016, 61, 633–645. [Google Scholar] [CrossRef]
- Leng, M.J.; Wagner, B.; Boehm, A.; Panagiotopoulos, K.; Vane, C.H.; Snelling, A.; Haidon, C.; Woodley, E.; Vogel, H.; Zanchetta, G.; et al. Understanding past climatic and hydrological variability in the Mediterranean from Lake Prespa sediment isotope and geochemical record over the last glacial cycle. Quat. Sci. Rev. 2013, 66, 123–136. [Google Scholar] [CrossRef]
- Hadjisolomou, E.; Stefanidis, K.; Papatheodorou, G.; Papastergiadou, E. Assessment of the eutrophication-related environmental parameters in two Mediterranean lakes by integrating statistical techniques and self-organizing maps. Int. J. Environ. Res. Public Health 2018, 15, 547. [Google Scholar] [CrossRef] [PubMed]
- Hollis, G.E.; Stevenson, A.C. The physical basis of the Lake Mikri Prespa systems: Geology, climate, hydrology and water quality. Hydrobiologia 1997, 351, 1–19. [Google Scholar] [CrossRef]
- Tryfon, E.; Moustaka-Gouni, M.; Nikolaidis, G.; Tsekos, I. Phytoplankton and physical-chemical features of the shallow Lake Mikri Prespa, Macedonia, Greece. Arch. Hydrobiol. 1994, 131, 477–494. [Google Scholar]
- Vardaka, E.; Moustaka-Gouni, M.; Cook, C.M.; Lanaras, T. Cyanobacterial blooms and water quality in Greek waterbodies. J. Appl. Phycol. 2005, 17, 391–401. [Google Scholar] [CrossRef]
- Christophoridis, C.; Zervou, S.-K.; Manolidi, K.; Katsiapi, M.; Moustaka-Gouni, M.; Kaloudis, T.; Triantis, T.M.; Hiskia, A. Occurrence and diversity of cyanotoxins in Greek lakes. Sci. Rep. 2018, 8, 17877. [Google Scholar] [CrossRef] [PubMed]
- Stamou, G.; Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E. Grazing potential—A functional plankton food web metric for ecological water quality assessment in Mediterranean lakes. Water 2019, 11, 1274. [Google Scholar] [CrossRef]
- Hampton, S.E.; McGowan, S.; Ozersky, T.; Virdis, S.G.P.; Vu, T.T.; Spanbauer, T.L.; Kraemer, B.M.; Swann, G.; Mackay, A.W.; Powers, S.M.; et al. Recent ecological change in ancient lakes. Limnol. Oceanogr. 2018, 63, 2277–2304. [Google Scholar] [CrossRef]
- European Environment Agency. Biogeographical Regions. Available online: http://www.eea.europa.eu/data-and-maps/figures/biogeographical-regions-in-europe-1 (accessed on 5 November 2019).
- Tryfon, E.; Moustaka-Gouni, M. Species composition and seasonal cycles of phytoplankton with special reference to the nanoplankton of Lake Mikri Prespa. Hydrobiologia 1997, 351, 61–75. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Sommer, U.; Economou-Amilli, A.; Arhonditsis, G.B.; Katsiapi, M.; Papastergiadou, E.; Kormas, K.A.; Vardaka, E.; Karayanni, H.; Papadimitriou, T. Implementation of the Water Framework Directive: Lessons learned and future perspectives for an ecologically meaningful classification based on phytoplankton of the status of Greek lakes, Mediterranean region. Environ. Manag. 2019, 64, 675–688. [Google Scholar] [CrossRef]
- Katsiapi, M.; Michaloudi, E.; Moustaka-Gouni, M.; Pahissa López, J. First ecological evaluation of the ancient Balkan Lake Megali Prespa based on plankton. J. Biol. Res. 2012, 17, 51–56. [Google Scholar]
- Parisopoulos, G.; Sapountzakis, G.; Georgiou, P.; Giamouri, M.; Malakou, M. Development of rule curves for sluice gate operation and water level management in lake Micro Prespa. In Proceedings of the 4th Conference on Water Observation and Information System for Decision Support (BALWOIS), Ohrid, Republic of Macedonia, 25–29 May 2010. [Google Scholar]
- Parisopoulos, G. Review of the Sluice Gate Operation and Water Level Management in Lake Micro Prespa; Technical Report; Society for the Protection of Prespa: Agios Germanos, Greece, 2016. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Chroococcales. In ‘Süsswasserflora von Mitteleuropa; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg, Germany, 1998; pp. 1–548. [Google Scholar]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Baselga, A. Partitioning abundance-based multiple-site dissimilarity into components: Balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef]
- Baselga, A. Multiplicative partition of true diversity yields independent alpha and beta components; additive partition does not. Ecology 2010, 91, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- Reshef, D.N.; Reshef, Y.A.; Finucane, H.K.; Grossman, S.R.; McVean, G.; Turnbaugh, P.J.; Lander, E.S.; Mitzenmacher, M.; Sabeti, P.C. Detecting novel associations in large data sets. Science 2011, 334, 1518–1524. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]
- Hernández-Ruiz, M.; Barber-Lluch, E.; Prieto, A.; Alvarez-Salgado, X.A.; Logares, R.; Teira, E. Seasonal succession of small planktonic eukaryotes inhabiting surface waters of a coastal upwelling system. Environ. Microbiol. 2018, 20, 2955–2973. [Google Scholar] [CrossRef]
- Monchamp, M.-E.; Spaak, P.; Domaizon, I.; Dubois, N.; Bouffard, D.; Pomati, F. Homogenization of lake cyanobacterial communities over a century of climate change and eutrophication. Nat. Ecol. Evol. 2018, 2, 317–324. [Google Scholar] [CrossRef]
- Stomp, M.; Huisman, J.; Mittelbach, G.G.; Litchman, E.; Klausmeier, C.A. Large-scale biodiversity patterns in freshwater phytoplankton. Ecology 2011, 92, 2096–2107. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Hiskia, A.; Genitsaris, S.; Katsiapi, M.; Manolidi, K.; Zervou, S.-K.; Christophoridis, C.; Triantis, T.M.; Kaloudis, T.; Orfanidis, S. First report of Aphanizomenon favaloroi occurrence in Europe associated with saxitoxins and a massive fish kill in Lake Vistonis, Greece. Mar. Freshw. Res. 2017, 68, 793–800. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velásquez, D.; Casero, M.C. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microb. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Lepistö, L.; Kauppila, P.; Rapala, J.; Pekkarinen, M.; Sammalkorpi, I.; Villa, L. Estimation of reference conditions for phytoplankton in a naturally eutrophic shallow lake. Hydrobiologia 2006, 568, 55–66. [Google Scholar] [CrossRef]
- Salmaso, N.; Capelli, C.; Shams, S.; Cerasino, L. Expansion of bloom-forming Dolichospermum lemmermannii (Nostocales, Cyanobacteria) to the deep lakes south of the Alps: Colonization patterns, driving forces and implications for water use. Harmful Algae 2015, 50, 76–87. [Google Scholar] [CrossRef]
- Finlay, B.J. Global dispersal of free-living microbial eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Vardaka, E.; Tryfon, E. Phytoplankton species succession in a shallow Mediterranean lake (L. Kastoria, Greece): Steady-state dominance of Limnothrix redekei, Microcystis aeruginosa and Cylindrospermopsis raciborskii. Hydrobiologia 2007, 575, 129–140. [Google Scholar] [CrossRef]
- Pujoni, D.G.F.; Barros, C.F.D.A.; Dos Santos, J.B.O.; Maia-Barbosa, P.M.; Barbosa, F.A.R. Dispersal ability and niche breadth act synergistically to determine zooplankton but nor phytoplankton metacommunity structure. J. Plankton Res. 2019, 41, 479–490. [Google Scholar] [CrossRef]
- Xie, L.; Hagar, J.; Rediske, R.R.; O’Keefe, J.; Dyble, J.; Hong, Y.; Steinman, A.D. The influence of environmental conditions and hydrologic connectivity on cyanobacteria assemblages in two drowned river mouth lakes. J. Gt. Lakes Res. 2011, 37, 470–479. [Google Scholar] [CrossRef]
- Soininen, J.; Köngäs, P. Analysis of nestedness in freshwater assemblages-patterns across species and trophic levels. Freshw. Sci. 2012, 31, 1145–1155. [Google Scholar] [CrossRef]
- Moritz, C.; Meynard, C.N.; Devictor, V.; Guizien, K.; Labrune, C.; Guarini, J.-M. Disentangling the role of connectivity, environmental filtering, and spatial structure on metacommunity dynamics. Oikos 2013, 122, 1401–1410. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Jähnig, S.C.; Haase, P. Contrasting metacommunity structure and beta diversity in an aquatic-floodplain system. Oikos 2016, 125, 686–697. [Google Scholar] [CrossRef]
- Wojciechowski, J.; Heino, J.; Bini, L.M.; Padial, A.A. Temporal variation in phytoplankton beta diversity patterns and metacommunity structures across subtropical reservoirs. Freshwater Biol. 2017, 62, 751–766. [Google Scholar] [CrossRef]
- Soininen, J.; Heino, J.; Wang, J. A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosystems. Glob. Ecol. Biogeogr. 2018, 27, 96–109. [Google Scholar] [CrossRef]
- Kraus, C.N.; Bonnet, M.-P.; Miranda, C.A.; de Souza Nogueira, I.; Garnier, J.; Vieira, L.C.G. Interannual hydrological variations and ecological phytoplankton patterns in Amazonian floodplain lakes. Hydrobiologia 2019, 830, 135–149. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | TB (mg L−1) | CB (mg L−1) | SR-ph | SR-cy | SI-ph | SI-cy | PhyCoIGP | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| September 2015 | 3.07 | 1.18 | 2.47 | 0.29 | 64 | 36 | 26 | 12 | 0.80 | 0.69 | 0.70 | 0.73 | 2.7 | 3.1 |
| October 2015 | 2.37 | 1.72 | 1.93 | 0.07 | 57 | 28 | 19 | 9 | 0.84 | 0.65 | 0.77 | 0.55 | 2.7 | 2.7 |
| November 2015 | 1.18 | 1.46 | 0.86 | 0.03 | 56 | 28 | 20 | 9 | 0.81 | 0.54 | 0.67 | 0.26 | 3.0 | 3.1 |
| December 2015 | 0.92 | 0.89 | 0.67 | 0.003 | 43 | 19 | 15 | 2 | 0.78 | 0.18 | - | - | 3.3 | 3.6 |
| January 2016 | 0.23 | 0.10 | 0.08 | 0.001 | 36 | 17 | 11 | 3 | 0.90 | 0.69 | - | - | 3.5 | 4.2 |
| February 2016 | 0.72 | 0.12 | 0.17 | 0.001 | 33 | 19 | 9 | 2 | 0.57 | 0.45 | - | - | 3.5 | 3.8 |
| March 2016 | 0.70 | 0.58 | 0.11 | 0.002 | 51 | 18 | 17 | 3 | 0.78 | 0.20 | - | - | 3.6 | 4.3 |
| April 2016 | 1.54 | 0.41 | 0.38 | 0.05 | 46 | 18 | 17 | 4 | 0.63 | 0.75 | 0.72 | 0.19 | 3.3 | 3.9 |
| June 2016 | 0.87 | 4.10 | 0.54 | 2.63 | 66 | 27 | 22 | 5 | 0.86 | 0.53 | 0.71 | 0.004 | 3.7 | 2.6 |
| July 2016 | 1.07 | 0.72 | 0.43 | 0.10 | 54 | 15 | 19 | 6 | 0.85 | 0.72 | 0.77 | 0.43 | 3.5 | 2.9 |
| August 2016 | 1.93 | 0.76 | 0.91 | 0.23 | 54 | 20 | 19 | 8 | 0.81 | 0.75 | 0.57 | 0.43 | 3.4 | 3.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsiapi, M.; Genitsaris, S.; Stefanidou, N.; Tsavdaridou, A.; Giannopoulou, I.; Stamou, G.; Michaloudi, E.; Mazaris, A.D.; Moustaka-Gouni, M. Ecological Connectivity in Two Ancient Lakes: Impact Upon Planktonic Cyanobacteria and Water Quality. Water 2020, 12, 18. https://doi.org/10.3390/w12010018
Katsiapi M, Genitsaris S, Stefanidou N, Tsavdaridou A, Giannopoulou I, Stamou G, Michaloudi E, Mazaris AD, Moustaka-Gouni M. Ecological Connectivity in Two Ancient Lakes: Impact Upon Planktonic Cyanobacteria and Water Quality. Water. 2020; 12(1):18. https://doi.org/10.3390/w12010018
Chicago/Turabian StyleKatsiapi, Matina, Savvas Genitsaris, Natassa Stefanidou, Anastasia Tsavdaridou, Irakleia Giannopoulou, Georgia Stamou, Evangelia Michaloudi, Antonios D. Mazaris, and Maria Moustaka-Gouni. 2020. "Ecological Connectivity in Two Ancient Lakes: Impact Upon Planktonic Cyanobacteria and Water Quality" Water 12, no. 1: 18. https://doi.org/10.3390/w12010018
APA StyleKatsiapi, M., Genitsaris, S., Stefanidou, N., Tsavdaridou, A., Giannopoulou, I., Stamou, G., Michaloudi, E., Mazaris, A. D., & Moustaka-Gouni, M. (2020). Ecological Connectivity in Two Ancient Lakes: Impact Upon Planktonic Cyanobacteria and Water Quality. Water, 12(1), 18. https://doi.org/10.3390/w12010018