Response of Physiological Indicators to Environmental Factors under Water Level Regulation of Paddy Fields in Southern China

Abstract
:1. Introduction
2. Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Method
3. Results and Discussion
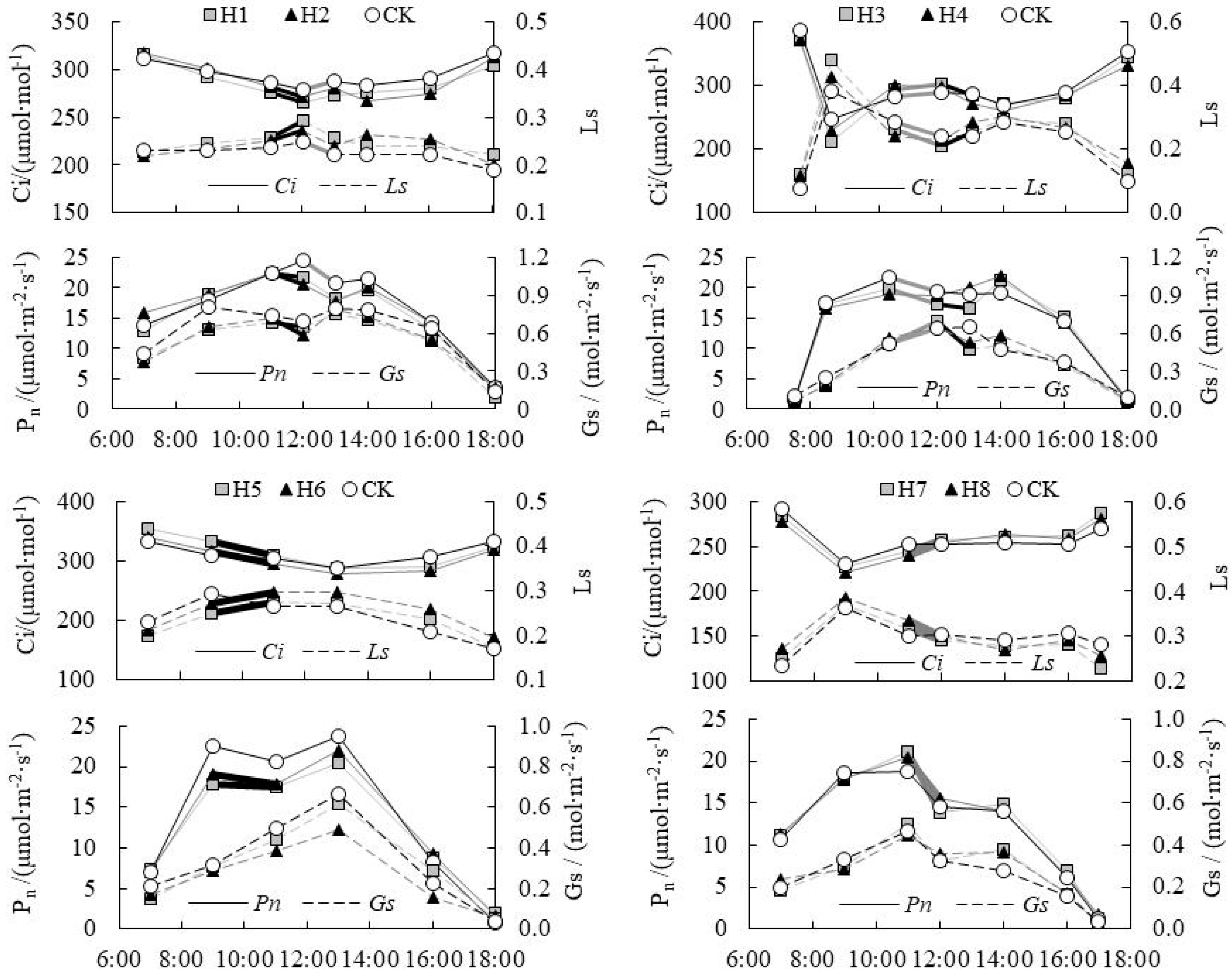
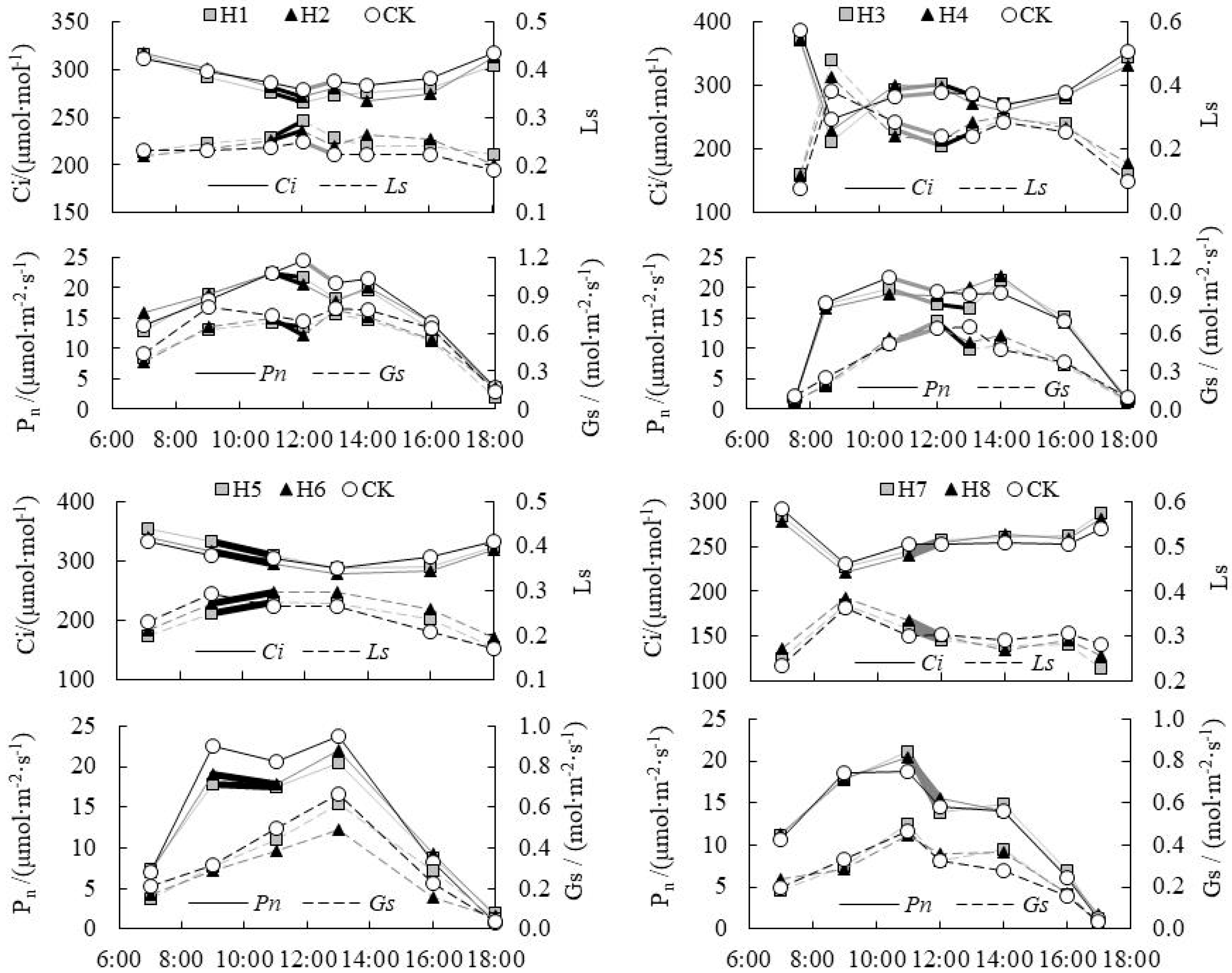
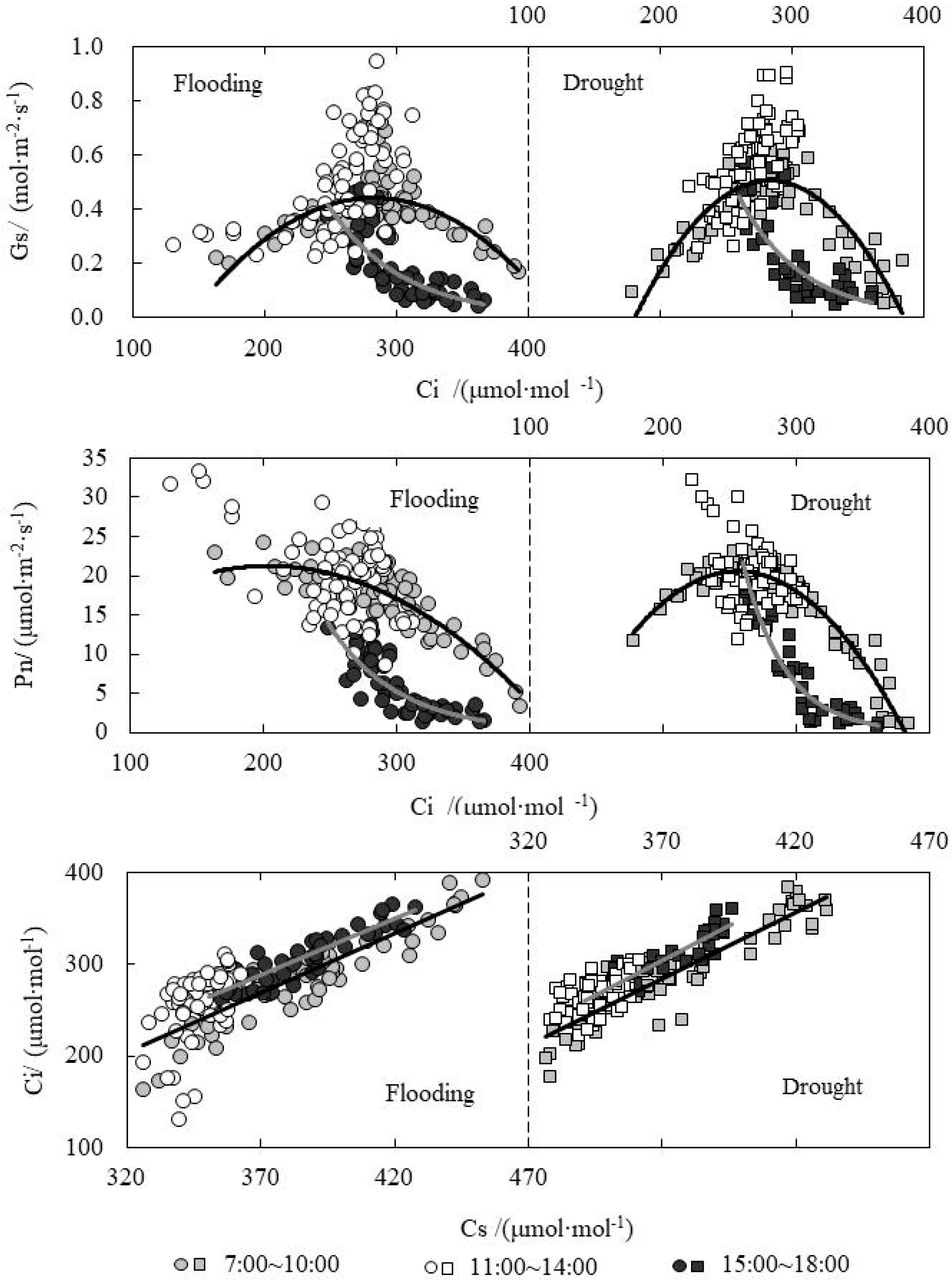
3.1. Collaborative Analysis on Change in Intercellular CO2 Concentration (Ci) and Stomatal/Non-Stomatal Limitation of Paddy Rice under Water Level Control
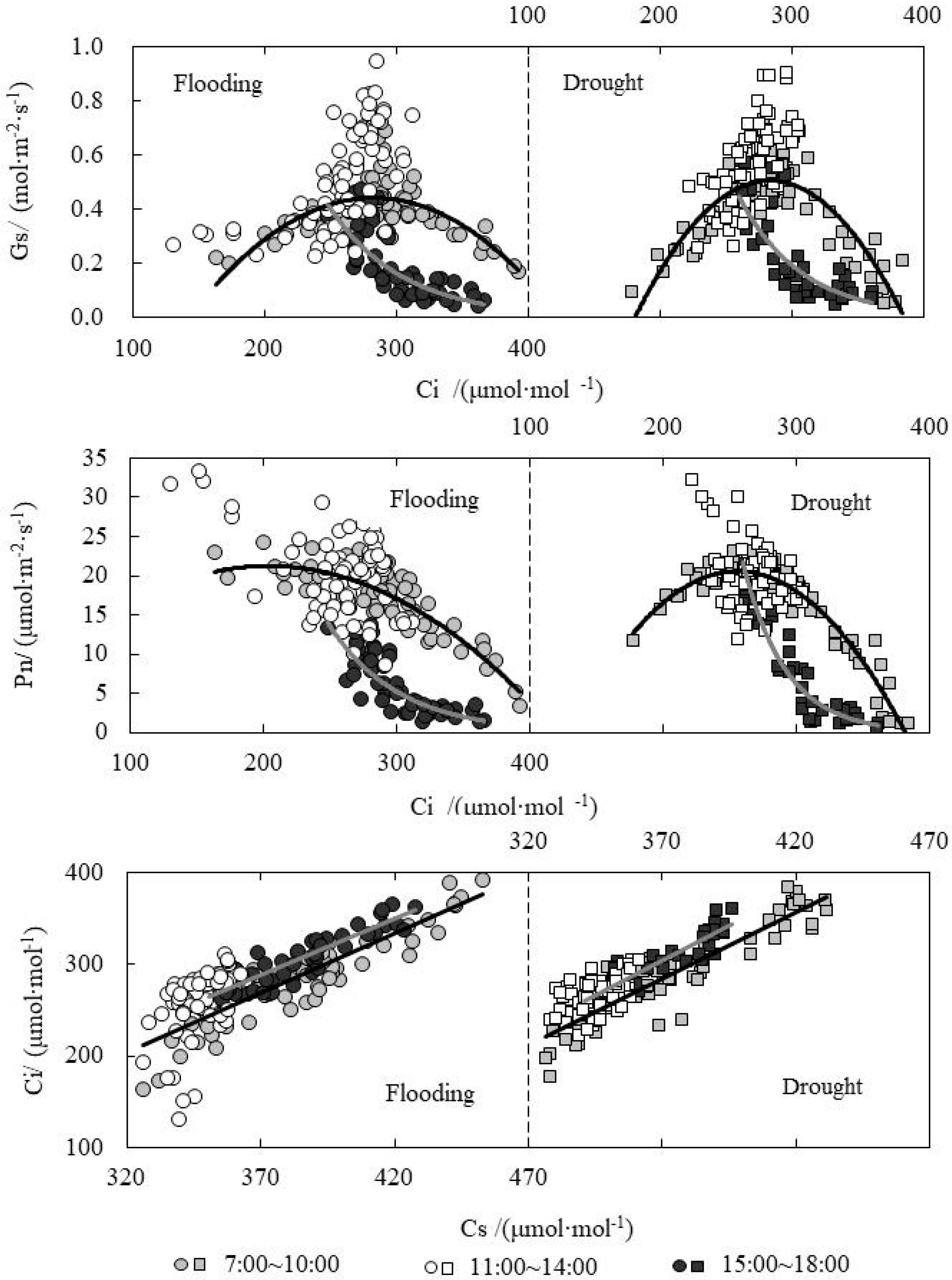
3.2. Response Relationship between Ci and Influencing Factors of Paddy Rice under Water Level Control
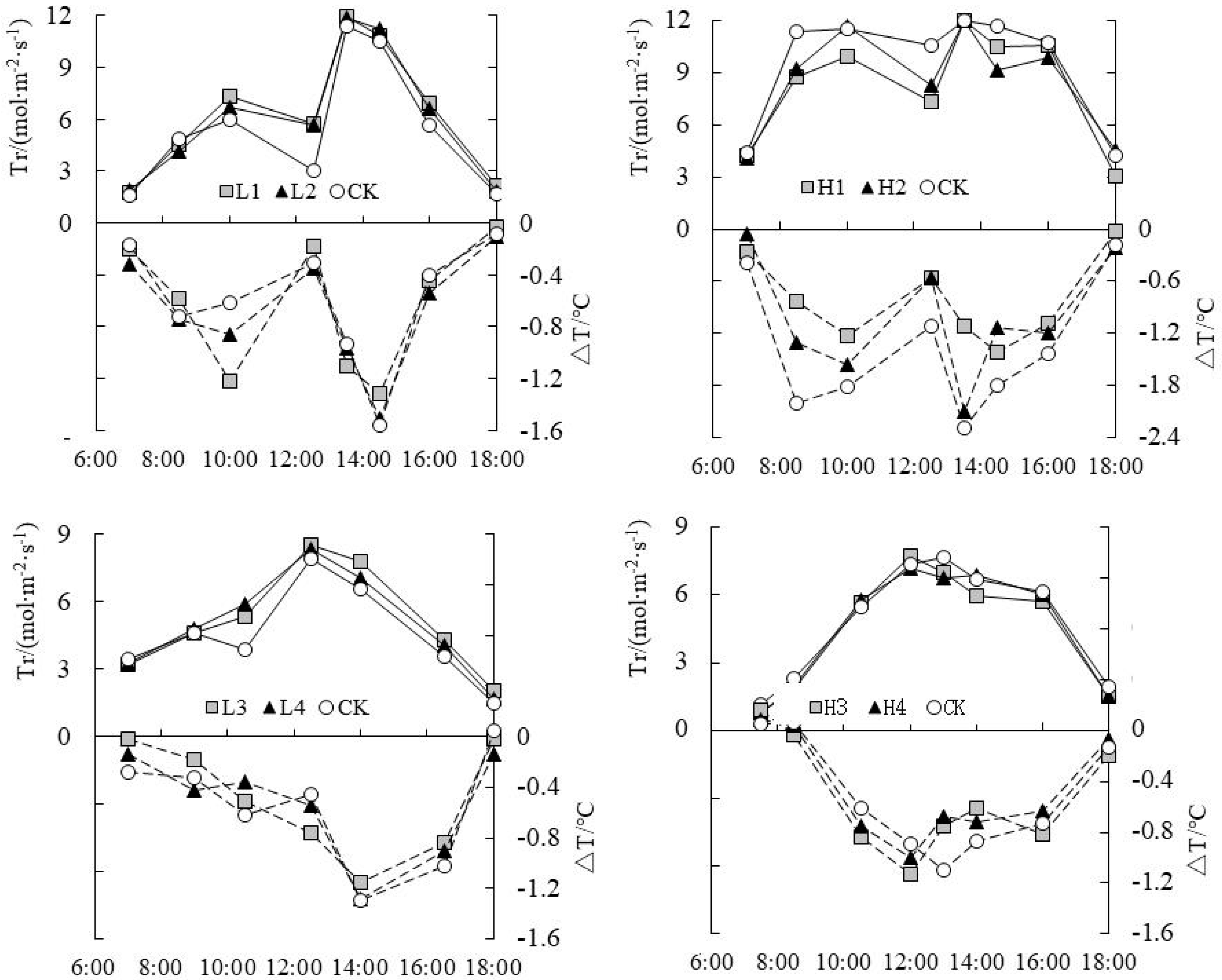
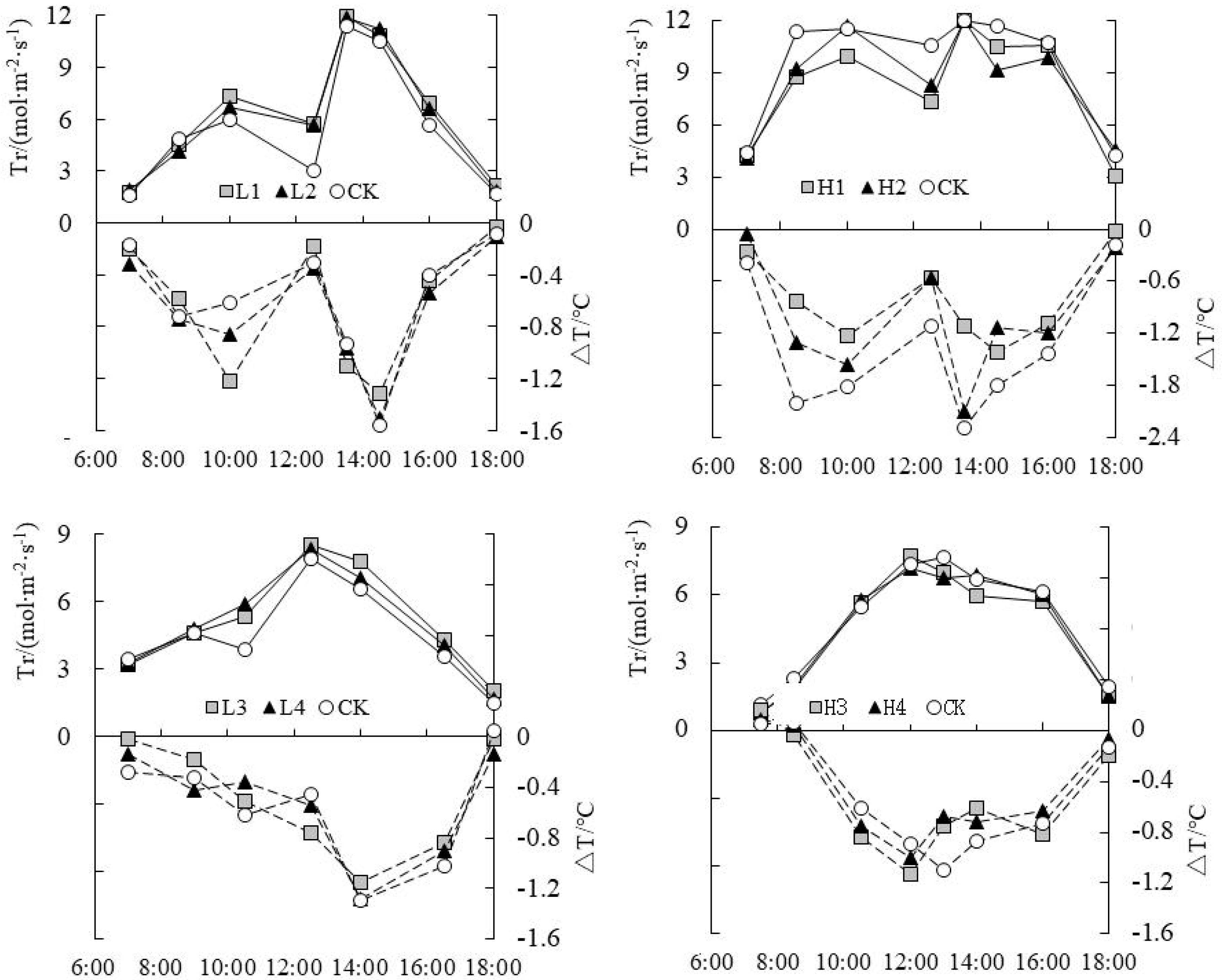
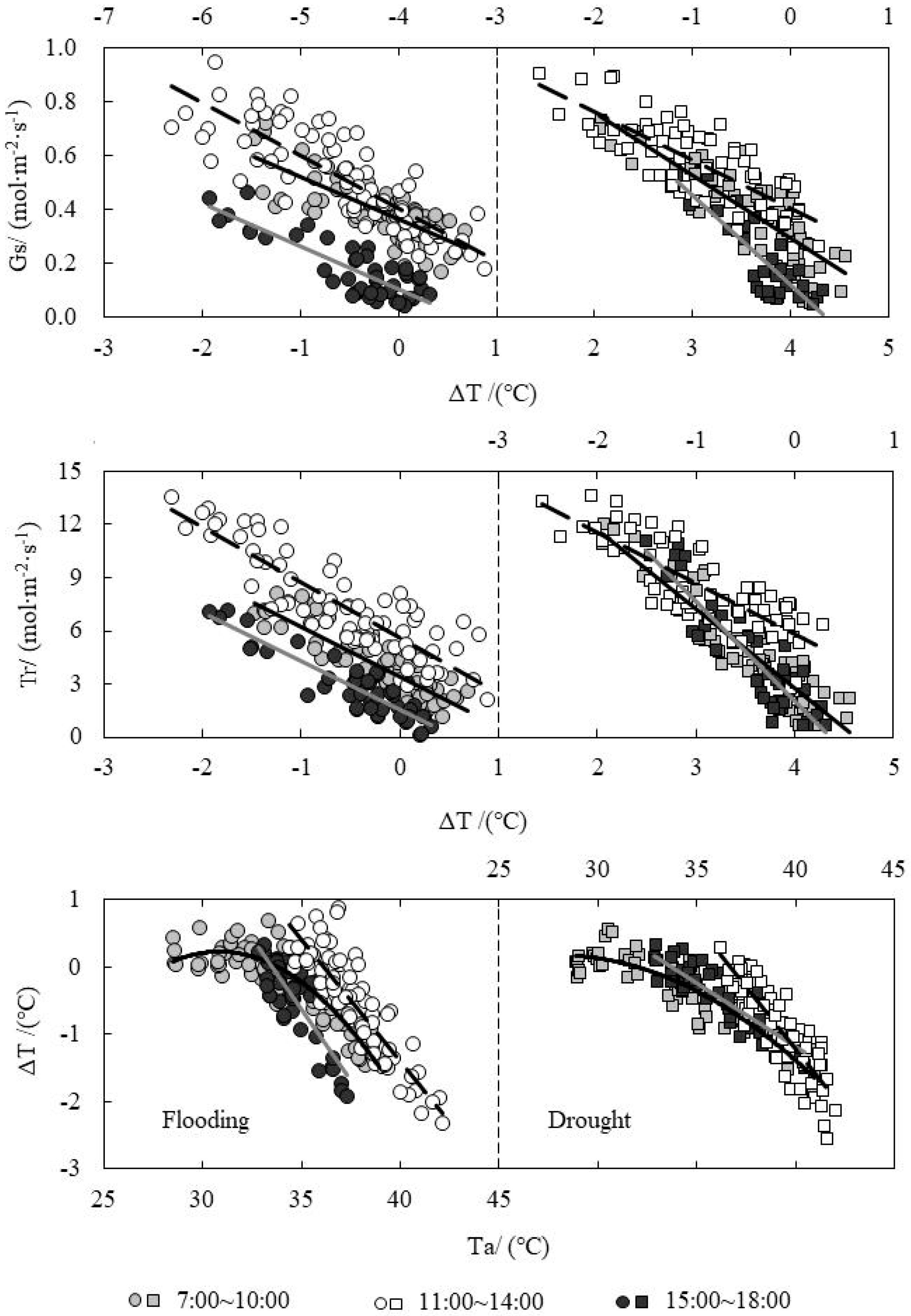
3.3. Analysis on Daily Change Law of Temperature Difference (ΔT) and Tr under Water Level Control
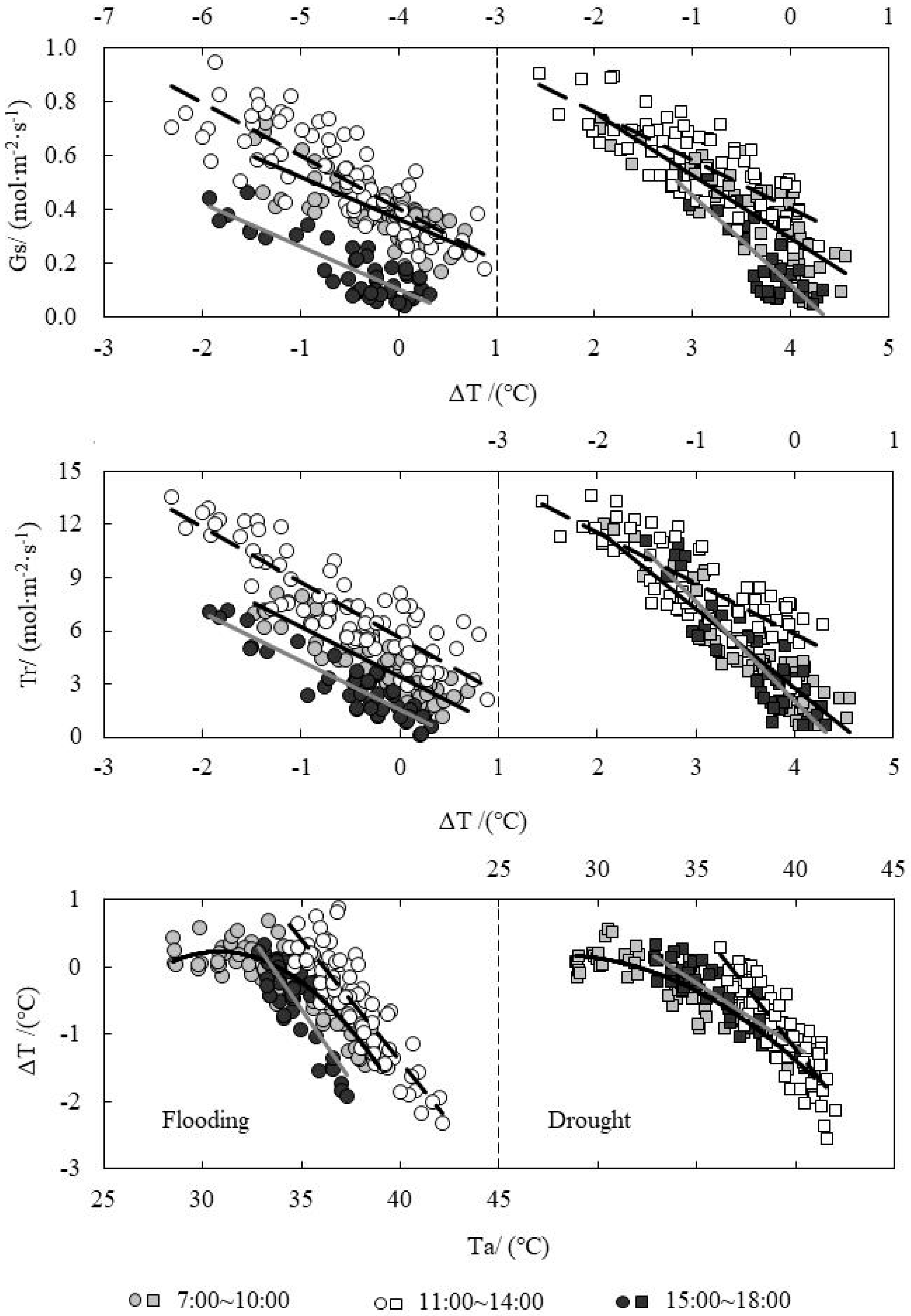
3.4. Response Relationship between ΔT and Influencing Factors under Water Level Control
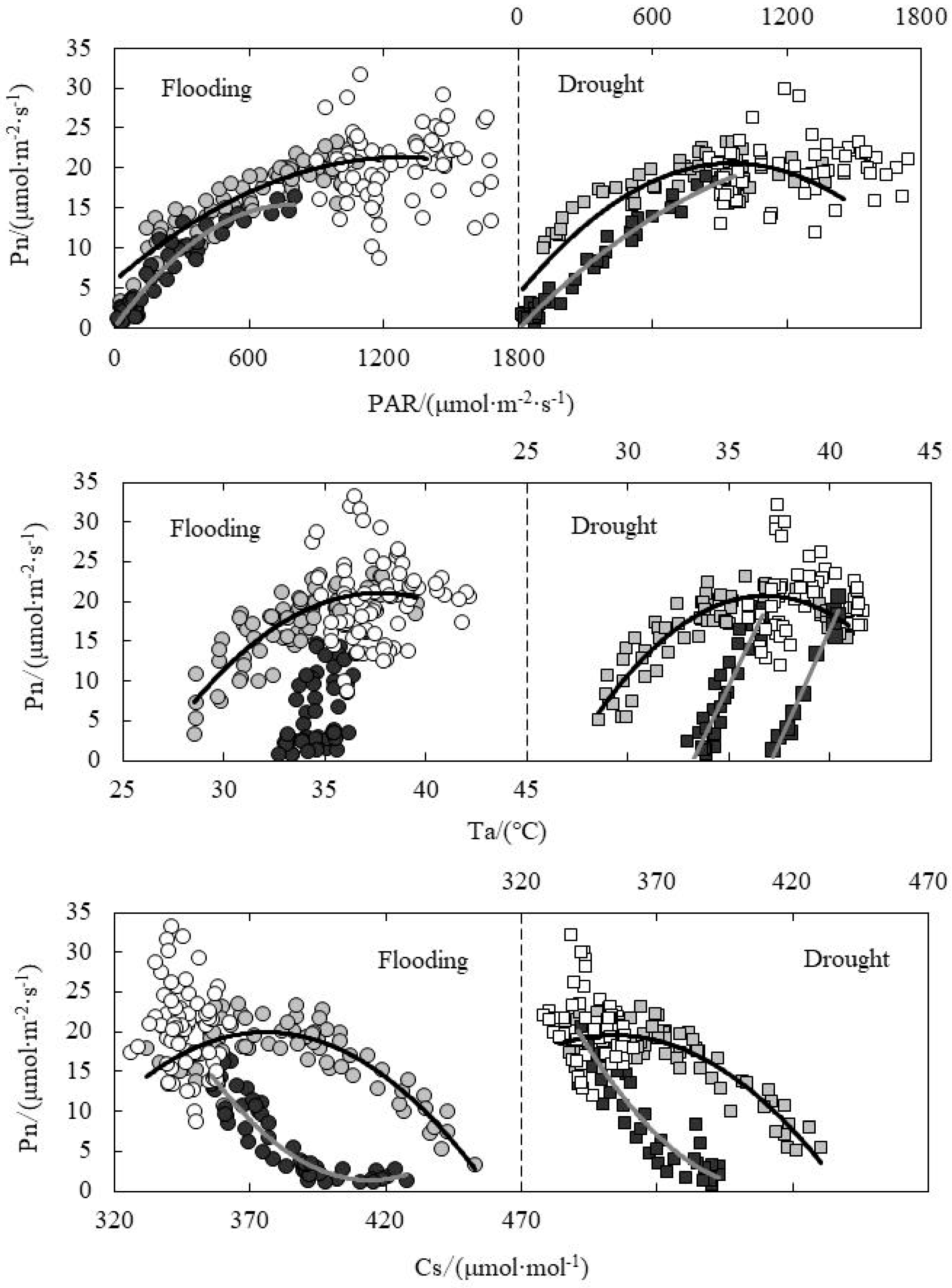
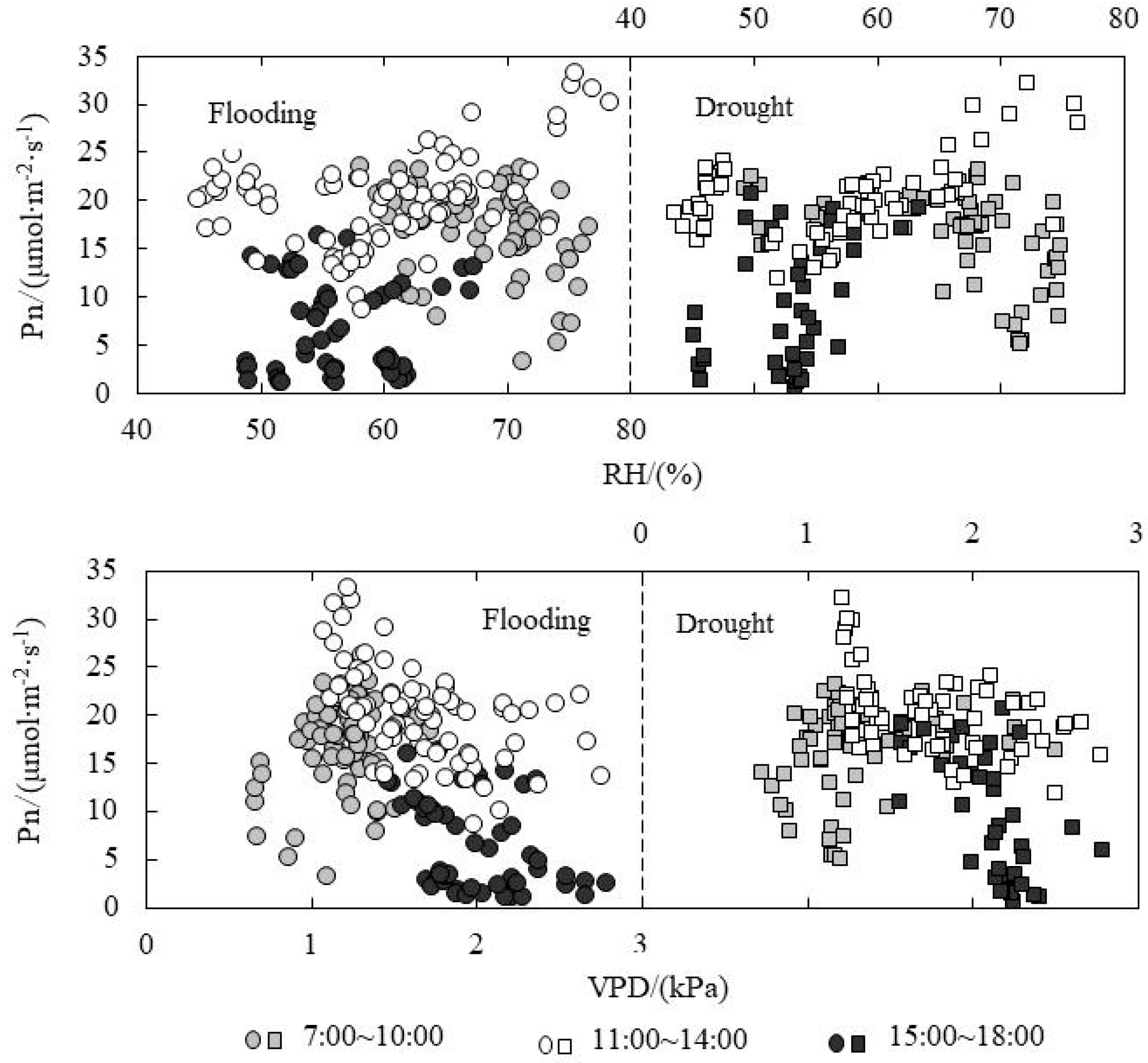
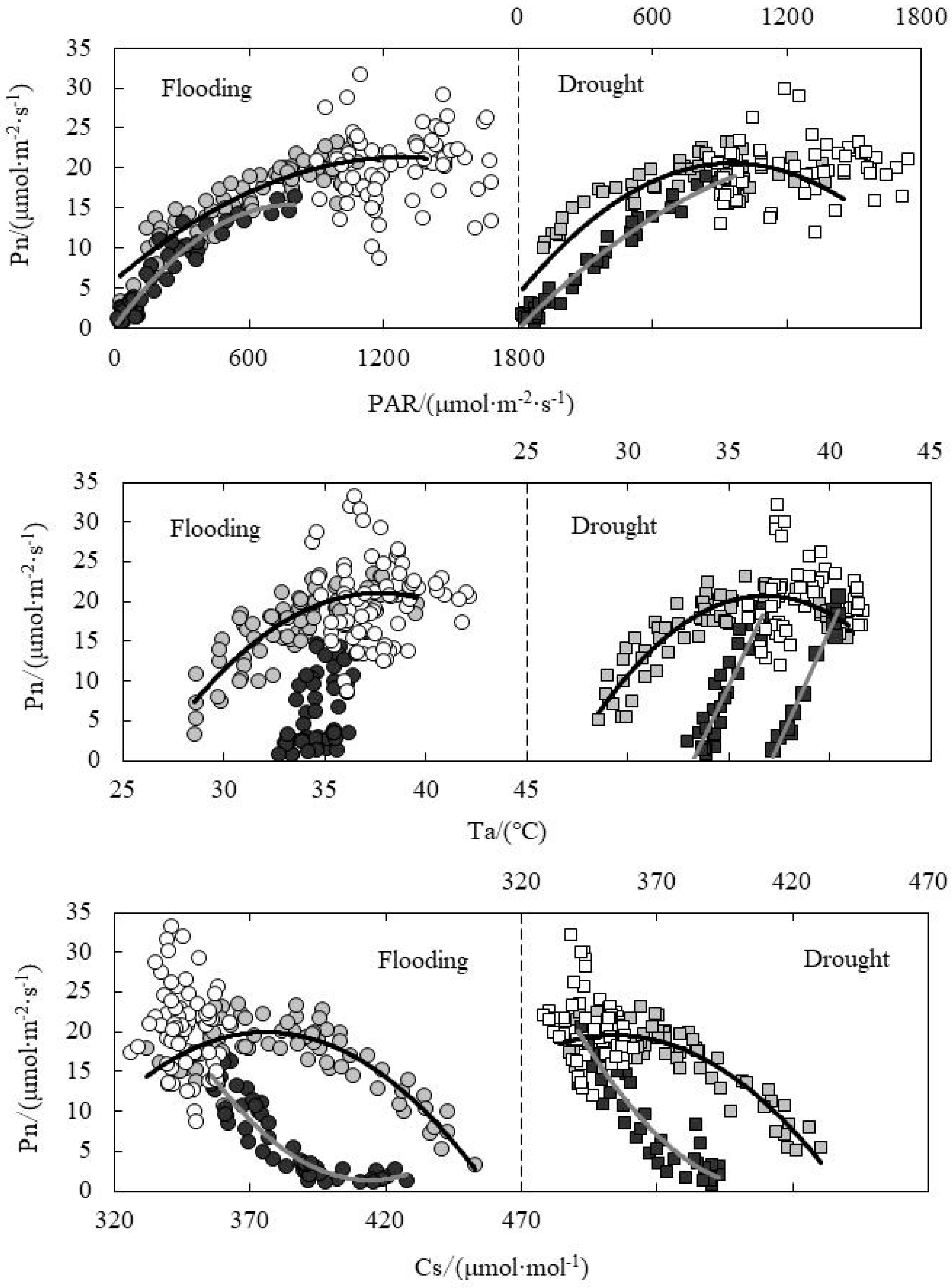
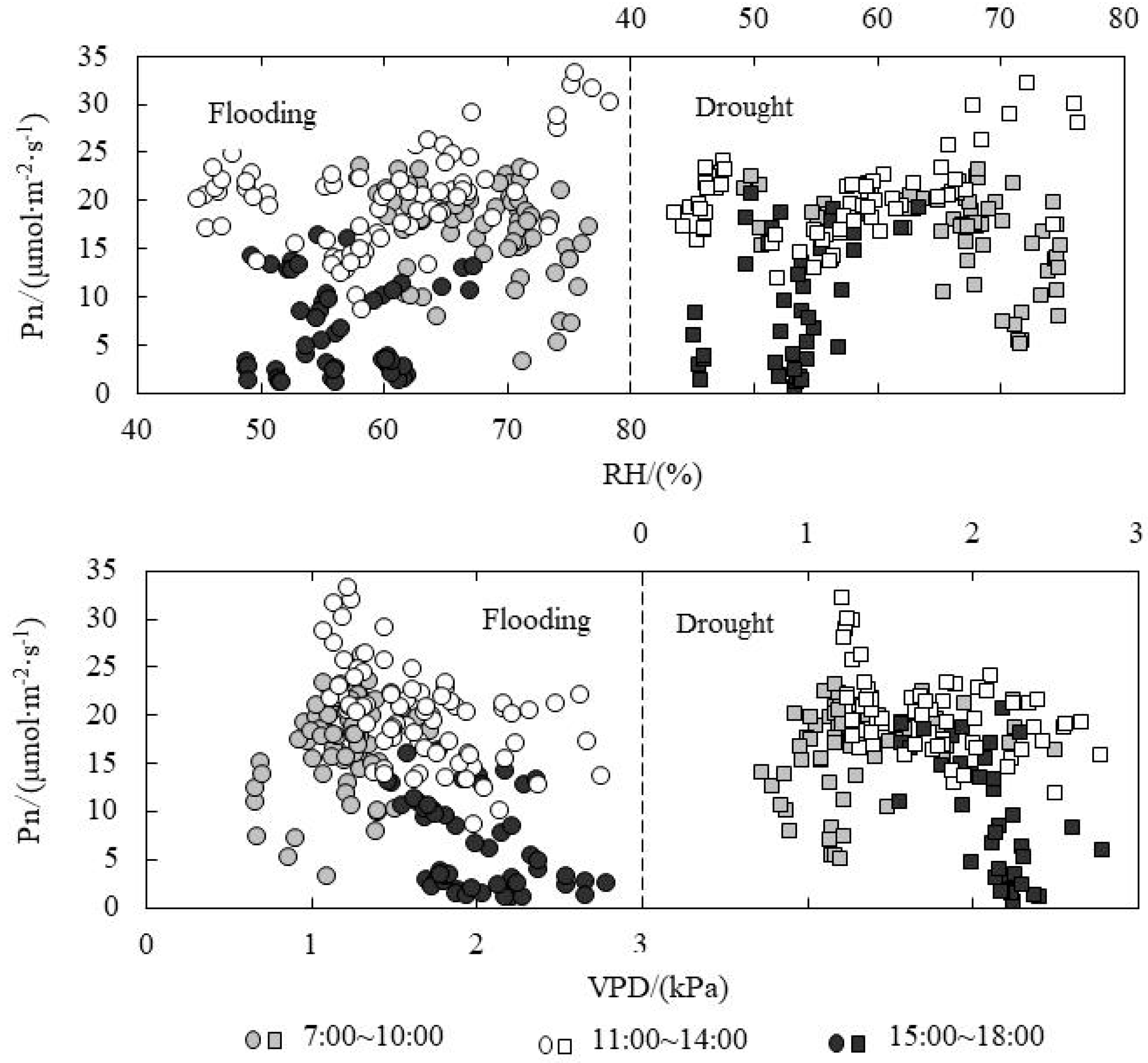
3.5. Response of Pn to Environmental Factors under Paddy Field Water Level Control
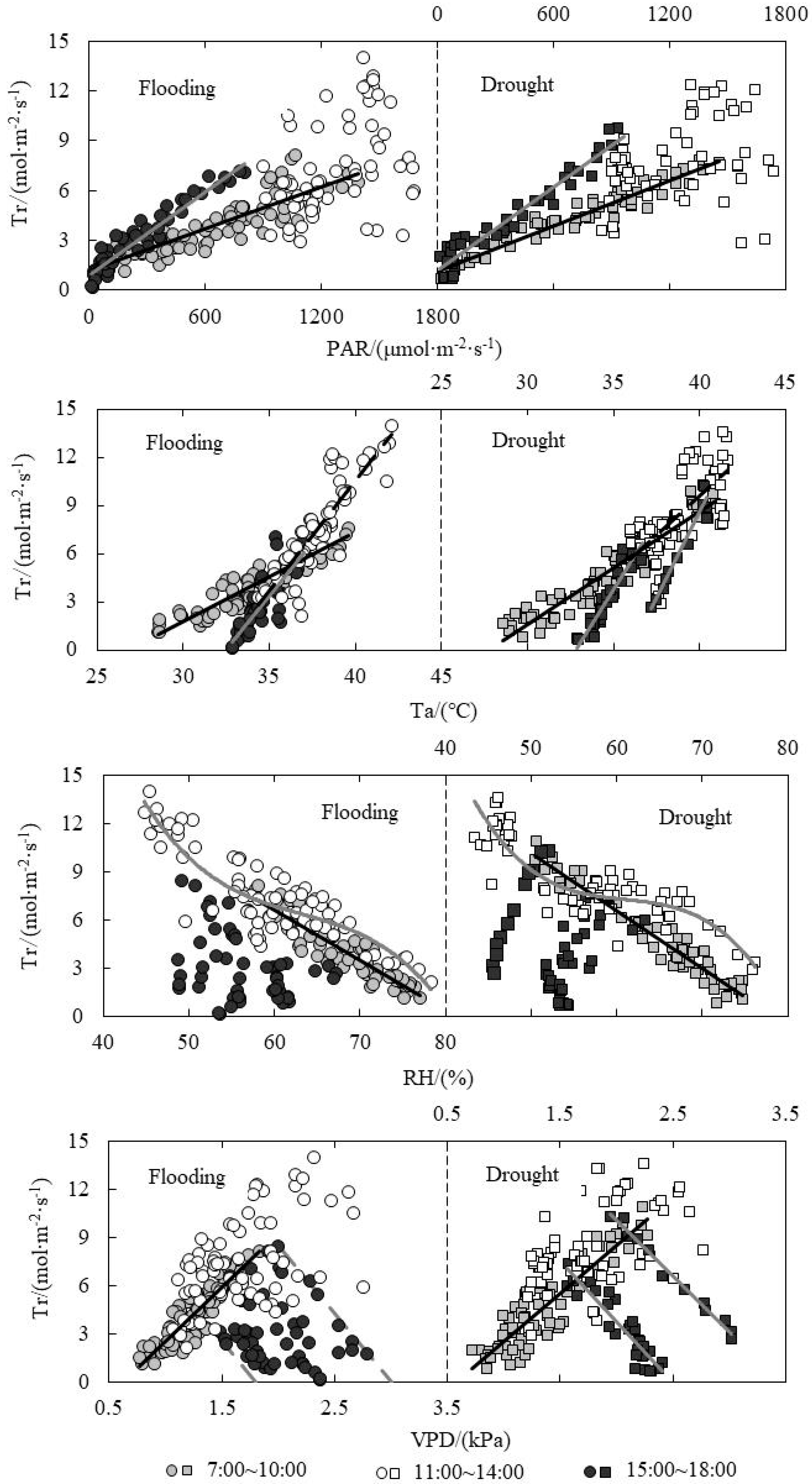
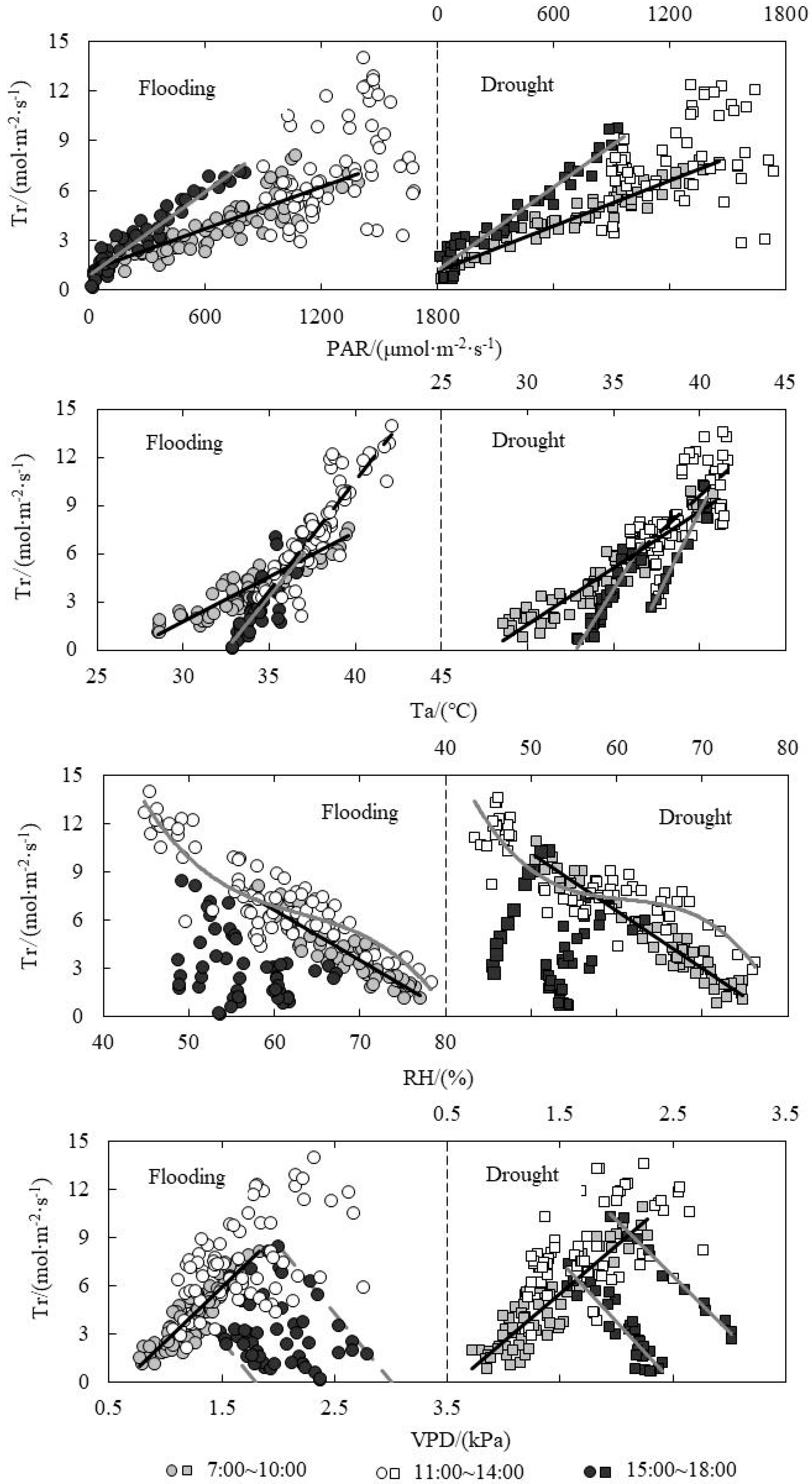
3.6. Response of Tr to Environmental Factors under Paddy Field Water Level Control
3.7. Regression Analysis between Physiological Indicators and Environmental Factors under Water Level Control
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mao, Z. Water Saving Irrigation for Rice and Its Effect on Environment. Eng. Sci. 2002, 4, 8–16. [Google Scholar]
- Xu, J.Z.; Peng, S.Z.; Yang, S.H.; Wang, W.G. Ammonia volatilization losses from a rice paddy with different irrigation and nitrogen managements. Agric. Water Manag. 2012, 104, 184–192. [Google Scholar] [CrossRef]
- Tan, X.; Shao, D.; Liu, H.; Yang, F.; Xiao, C.; Yang, H. Effects of alternate wetting and drying irrigation on percolation and nitrogen leaching in paddy fields. Paddy Water Environ. 2013, 11, 381–395. [Google Scholar] [CrossRef]
- Wang, S.L.; Wang, X.G.; Brown, L.C.; Qu, X. Current status and prospects of agricultural drainage in China. Irrig. Drain. 2010, 56, 47–58. [Google Scholar]
- Tian, S.; Youssef, M.A.; Skaggs, R.W.; Amatya, D.M.; Chescheir, G.M. Drainmod-forest: Integrated modeling of hydrology, soil carbon and nitrogen dynamics, and plant growth for drained forests. J. Environ. Qual. 2012, 41, 764. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.E.; Miao, Z.M.; Xing, W.G.; Shao, G.C.; Jiang, Y.X. Research advance on irrigation and drainage for rice by using water level as regulation index. J. Irrig. Drain. 2010, 29, 134–136. [Google Scholar]
- Xiao, M.H.; Yu, S.E.; She, D.; Hu, X.J.; Chu, L.L. Nitrogen and phosphorus loss and optimal drainage time of paddy field under controlled drainage condition. Arab. J. Geosci. 2015, 8, 4411–4420. [Google Scholar] [CrossRef]
- Xiao, M.H.; Miao, Z.M.; Li, Y.Y. Changes of root-zone soil environment in flooded paddy field under controlled drainage conditions. Pol. J. Environ. Stud. 2017, 2, 881–892. [Google Scholar] [CrossRef]
- Jiang, Q.; Roche, D.; Monaco, T.A.; Durham, S. Gas exchange, chlorophyll fluorescence parameters and carbon isotope discrimination of 14 barley genetic lines in response to salinity. Field Crops Res. 2006, 96, 269–278. [Google Scholar] [CrossRef]
- Kiani-Pouya, A. Changes in activities of antioxidant enzymes and photosynthetic attributes in triticale (Triticosecale, Wittmack) genotypes in response to long-term salt stress at two distinct growth stages. Acta Physiol. Plant. 2015, 37, 1–11. [Google Scholar] [CrossRef]
- Miao, Z.M.; Yu, S.E.; Lu, B.; Ding, J.H.; Yu, Z.H. Relationships of water requirement photosynthesis production for paddy rice using structural equation modeling. Trans. Chin. Soc. Agric. Eng. 2013, 29, 91–98. [Google Scholar]
- Wang, W.; Xia, M.X.; Chen, J.; Yuan, R.; Deng, F.N.; Shen, F.F. Gene expression characteristics and regulation mechanisms of superoxide dismutase and its physiological roles in plants under stress. Biochemistry 2016, 81, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Li, S.S. Effect of submergence stress on the distribution of starch granules in leaf sheath and leaf of rice. J. Wuhan Bot. Res. 2000, 18, 528–530. [Google Scholar]
- Hirano, T.; Uchida, N.; Azuma, T.; Yasuda, T. Effect of submergence on distribution of photoassimilates and activities of sucrose metabolizing enzymes in sink organs of floating rice. Jpn. J. Crop Sci. 2008, 65, 540–548. [Google Scholar] [CrossRef]
- Hu, J.; Cao, W.; Jiang, D.; Luo, W. Quantification of water stress factor for crop growth simulation I. Effects of drought and waterlogging stress on photosynthesis, transpiration and dry matter partitioning in winter wheat. Acta Agron. Sin. 2004, 30, 315–320. [Google Scholar]
- Lv, J.; Li, C.; Zhang, J.; Li, F. Effect of drought on distribution of carbon assimilation in different genotypes of spring wheat. Agric. Res. Arid Areas 2005, 21, 77–81. [Google Scholar]
- Zeppel, M.J.B.; Lewis, J.D.; Chaszar, B.; Smith, R.A.; Medlyn, B.E.; Huxman, T.E. Nocturnal stomatal conductance responses to rising [CO2], temperature and drought. New Phytol. 2012, 193, 929–938. [Google Scholar] [CrossRef]
- Wu, X.H.; Wang, W.; Xie, X.L.; Yin, C.M.; Xie, K.J. Photosynthetic and yield responses of rice (Oryza sativa, L.) to different water management strategies in subtropical China. Photosynthetica 2018, 1, 1–8. [Google Scholar] [CrossRef]
- Zhang, L.X.; Peng, L.W.; Lin, R.C.; Lu, C.M.; Kuang, T.Y. Advances and Prospects of Photosynthesis Research. China Basic Sci. 2016, 1, 13–20. [Google Scholar]
- Ming, D.F.; Pei, Z.F.; Naeem, M.S.; Gong, H.J.; Zhou, W.J. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Wang, J.Y.; Jia, J.X.; Xiong, Z.Q.; Khalil, M.A.K.; Xing, G.X. Water regime–nitrogen fertilizer–straw incorporation interaction: Field study on nitrous oxide emissions from a rice agroecosystem in Nanjing, China. Agric. Ecosyst. Environ. 2011, 141, 437–446. [Google Scholar] [CrossRef]
- Tan, X.; Shao, D.; Gu, W.; Liu, H. Field analysis of water and nitrogen fate in lowland paddy fields under different water managements using HYDRUS–D. Agric. Water Manag. 2015, 150, 67–80. [Google Scholar] [CrossRef]
- He, Y.; Zhang, J.; Xu, J.; Hong, D.; Yang, S. Regulation and control of water transformation through coupling irrigation and drainage in paddy field. Trans. Chin. Soc. Agric. Eng. 2016, 32, 144–149. [Google Scholar]
- Huang, M.; Shan, S.L.; Zhou, X.F.; Chen, J.; Cao, F.B.; Jiang, L.G.; Zou, Y.B. Leaf photosynthetic performance related to higher radiation use efficiency and grain yield in hybrid rice. Field Crops Res. 2016, 193, 87–93. [Google Scholar] [CrossRef]
- Xu, J.Z.; Lv, Y.; Liu, X.; Dalson, T.; Yang, S.; Wu, J. Diagnosing crop water stress of rice using infrared thermal imager under water deficit condition. Int. J. Agric. Biol. 2016, 18, 565–572. [Google Scholar] [CrossRef]
- Xia, J.; Zhang, S.; Guo, J.; Rong, Q.; Zhang, G. Critical effects of gas exchange parameters in Tamarix Chinensis, Lour on soil water and its relevant environmental factors on a shell ridge island in China’s Yellow River Delta. Ecol. Eng. 2015, 76, 36–46. [Google Scholar] [CrossRef]
- Lima Neto, M.C.; Martins, M.D.O.; Ferreirasilva, S.L.; Silveira, J.A.G. Jatropha curcas and Ricinus communis display contrasting photosynthetic mechanisms in response to environmental conditions. Sci. Agric. 2015, 72, 260–269. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Tillering | Jointing-Booting | Heading-Flowering | Milky | Starting/ Ending Time | |
|---|---|---|---|---|---|---|
| (07.05~08.05) | (08.06~08.26) | (08.27~09.09) | (09.10~10.10) | |||
| Water flooding | L1 | 120 mm/2 mm/day | −300~30 mm | −300~30 mm | 300~30 mm | 07.19~07.28 |
| L2 | 120 mm/4 mm/day | −300~30 mm | −300~30 mm | 300~30 mm | ||
| L3 | −200~20 mm | 250 mm/2 mm/day | −300~30 mm | 300~30 mm | 08.14~08.23 | |
| L4 | −200~20 mm | 250 mm/4 mm/day | −300~30 mm | 300~30 mm | ||
| L5 | −200~20 mm | −300~30 mm | 250 mm/2 mm/day | 300~30 mm | 08.30~09.08 | |
| L6 | −200~20 mm | −300~30 mm | 250 mm/4 mm/day | 300~30 mm | ||
| L7 | −200~20 mm | −300~30 mm | −300~30 mm | 250 mm/2 mm/day | 09.12~09.21 | |
| L8 | −200~20 mm | −300~30 mm | −300~30 mm | 250 mm/2 mm/day | ||
| Water drought | H1 | −300 mm | −300~30 mm | −300~30 mm | −300~30 mm | 07.19~ |
| H2 | −500 mm | −300~30 mm | −300~30 mm | −300~30 mm | ||
| H3 | −200~20 mm | −400 mm | −300~30 mm | −300~30 mm | 08.14~ | |
| H4 | −200~20 mm | −700 mm | −300~30 mm | −300~30 mm | ||
| H5 | −200~20 mm | −300~30 mm | −400 mm | −300~30 mm | 08.30~ | |
| H6 | −200~20 mm | −300~30 mm | −700 mm | −300~30 mm | ||
| H7 | −200~20 mm | −300~30 mm | −300~30 mm | −400 mm | 09.12~ | |
| H8 | −200~20 mm | −300~30 mm | −300~30 mm | −700 mm | ||
| Contrast treatment | CK | −300~30 mm | −300~30 mm | −300~30 mm | −300~30 mm | |
| Relationship | Treatment | Time | Fitting Formula | R2 | F | p |
|---|---|---|---|---|---|---|
| Relationship between Gs and Ci | Flooding | A.M. | y = −2 × 10−5x2 + 0.0126x − 1.3325 | 0.46 | 29.383 | <0.001 |
| P.M. | y = 35.813e−0.018x | 0.6479 | 92.013 | <0.001 | ||
| Drought | A.M. | y = −5 × 10−5x2 + 0.0281x − 3.4711 | 0.7378 | 74.416 | <0.001 | |
| P.M. | y = 99.209e−0.0208x | 0.7006 | 100.643 | <0.001 | ||
| Relationship between Pn and Ci | Flooding | A.M. | y = −0.0005x2 + 0.1841x + 2.4961 | 0.6772 | 72.362 | <0.001 |
| P.M. | y = 1981.8e−0.0197x | 0.73 | 119.998 | <0.001 | ||
| Drought | A.M. | y = −0.0013x2 + 0.6602x − 63.89 | 0.8893 | 245.122 | <0.001 | |
| P.M. | y = 65729e−0.0308x | 0.851 | 241.335 | <0.001 | ||
| Relationship between Cs and Ci | Flooding | A.M. | y = 1.3025x − 212.41 | 0.7598 | 221.447 | <0.001 |
| P.M. | y = 1.2263x − 165.55 | 0.8077 | 210.064 | <0.001 | ||
| Drought | A.M. | y = 1.4527x − 252.94 | 0.8248 | 291.951 | <0.001 | |
| P.M. | y = 1.5296x − 261.17 | 0.8484 | 218.588 | <0.001 | ||
| Relationship between Gs and ΔT | Flooding | A.M. | y = −0.1557x + 0.3665 | 0.5753 | 94.831 | <0.001 |
| P.M. | y = −0.1596x + 0.1036 | 0.7219 | 112.732 | <0.001 | ||
| Drought | A.M. | y = −0.233x + 0.2943 | 0.702 | 146.03 | <0.001 | |
| P.M. | y = −0.3347x + 0.1182 | 0.7189 | 79.273 | <0.001 | ||
| Relationship between Tr and ΔT | Flooding | A.M. | y = −2.837x + 3.3961 | 0.7076 | 169.394 | <0.001 |
| P.M. | y = −2.7517x + 1.5614 | 0.8123 | 199.008 | <0.001 | ||
| Drought | A.M. | y = −4.4699x + 2.7667 | 0.8357 | 315.382 | <0.001 | |
| P.M | y = −5.6112x + 2.0958 | 0.8182 | 193.488 | <0.001 | ||
| Relationship between Ta and ΔT | Flooding | A.M. | y = −0.0262x2 + 1.6172x − 24.73 | 0.7504 | 103.715 | <0.001 |
| P.M | y = −0.4235x + 14.177 | 0.7206 | 118.612 | <0.001 | ||
| Drought | A.M. | y = −0.0108x2 + 0.6023x − 8.2518 | 0.8446 | 165.825 | <0.001 | |
| P.M | y = −0.1878x + 6.3287 | 0.8092 | 182.344 | <0.001 | ||
| Relationship between Pn and PAR | Flooding | A.M. | y = −9 × 10−6x2 + 0.0240x + 5.9349 | 0.8053 | 142.733 | <0.001 |
| P.M. | y = −3 × 10−5x2 + 0.0426x + 0.0023 | 0.9101 | 280.226 | <0.001 | ||
| Drought | A.M. | y = −2 × 10−5x2 + 0.0346x + 4.0830 | 0.8302 | 149.14 | <0.001 | |
| P.M. | y = −9 × 10−6x2 + 0.0287x − 0.1358 | 0.9684 | 643.306 | <0.001 | ||
| Relationship between Pn and Ta | Flooding | A.M. | y = −0.1648x2 + 12.414x − 212.73 | 0.7026 | 81.496 | <0.001 |
| P.M. | y = 2.4912x − 80.012 | 0.3081 | 22.26 | <0.001 | ||
| Drought | A.M. | y = −0.2178x2 + 16.019x − 273.79 | 0.8019 | 123.476 | <0.001 | |
| P.M. | y = 6.7255x − 252.53 | 0.971 | 309.36 | <0.001 | ||
| Relationship between Pn and Cs | Flooding | A.M. | y = −0.0029x2 + 2.1674x − 387.24 | 0.7322 | 94.308 | <0.001 |
| P.M. | y = 0.0039x2 − 3.2062x + 665.26 | 0.8504 | 139.257 | <0.001 | ||
| Drought | A.M. | y = −0.0028x2 + 1.9558x − 326.94 | 0.8036 | 124.807 | <0.001 | |
| P.M. | y = 0.004x2 − 3.3269x + 683.67 | 0.8392 | 109.567 | <0.001 | ||
| Relationship between Tr and PAR | Flooding | A.M. | y = 0.0042x + 1.1463 | 0.7271 | 186.521 | <0.001 |
| P.M. | y = 0.0084x + 0.9127 | 0.9188 | 565.615 | <0.001 | ||
| Drought | A.M. | y = 0.0046x + 1.0649 | 0.9396 | 964.164 | <0.001 | |
| P.M. | y = 0.0084x + 1.1153 | 0.9452 | 741.373 | <0.001 | ||
| Relationship between Tr and Ta | Flooding | A.M. | y = 0.5614x − 15.064 | 0.8208 | 320.659 | <0.001 |
| P.M. | y = 1.3036x − 42.231 | 0.7071 | 111.074 | <0.001 | ||
| Drought | A.M. | y = 0.7032x − 19.464 | 0.8986 | 549.549 | <0.001 | |
| P.M. | y = 2.1467x − 77.274 | 0.9396 | 155.531 | <0.001 | ||
| Relationship between Tr and RH | Flooding | A.M. | y = −0.314x + 25.466 | 0.8291 | 339.604 | <0.001 |
| P.M. | y = −0.0007x3 + 0.1317x2 − 8.4573x + 190.44 | 0.7841 | 113.719 | <0.001 | ||
| Drought | A.M. | y = −0.3563x + 27.942 | 0.9402 | 974.249 | <0.001 | |
| P.M. | y = −0.0009x3 + 0.1736x2 − 10.642x + 225.78 | 0.7801 | 83.152 | <0.001 | ||
| Relationship between Tr and VPD | Flooding | A.M. | y = 6.6981x − 4.1526 | 0.747 | 206.728 | <0.001 |
| Drought | A.M. | y = 5.9954x − 3.5076 | 0.8193 | 281.098 | <0.001 | |
| P.M. | y = −7.6055x + 18.933 | 0.8481 | 173.092 | <0.001 | ||
| y = −6.8669x + 23.748 (Tillering) | 0.9831 | 583.217 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, M.; Li, Y.; Lu, B.; Miao, Z. Response of Physiological Indicators to Environmental Factors under Water Level Regulation of Paddy Fields in Southern China. Water 2018, 10, 1772. https://doi.org/10.3390/w10121772
Xiao M, Li Y, Lu B, Miao Z. Response of Physiological Indicators to Environmental Factors under Water Level Regulation of Paddy Fields in Southern China. Water. 2018; 10(12):1772. https://doi.org/10.3390/w10121772
Chicago/Turabian StyleXiao, Menghua, Yuanyuan Li, Bin Lu, and Zimei Miao. 2018. "Response of Physiological Indicators to Environmental Factors under Water Level Regulation of Paddy Fields in Southern China" Water 10, no. 12: 1772. https://doi.org/10.3390/w10121772
APA StyleXiao, M., Li, Y., Lu, B., & Miao, Z. (2018). Response of Physiological Indicators to Environmental Factors under Water Level Regulation of Paddy Fields in Southern China. Water, 10(12), 1772. https://doi.org/10.3390/w10121772