Carbon and Nitrogen Isotopic Signatures of Zooplankton Taxa in Five Small Subalpine Lakes along a Trophic Gradient

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
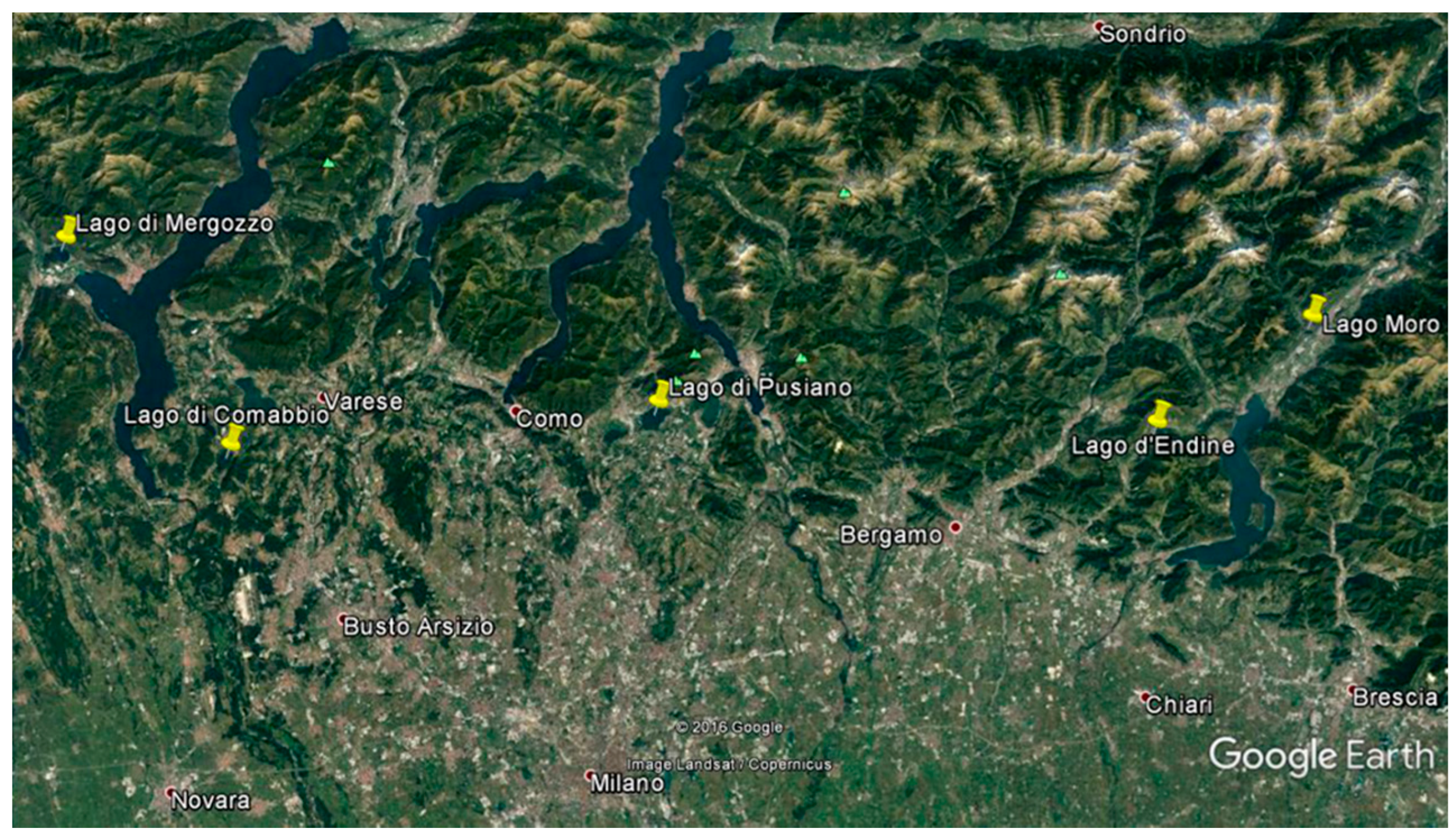
2.1. Study Sites
2.2. Zooplankton Sampling and SIA
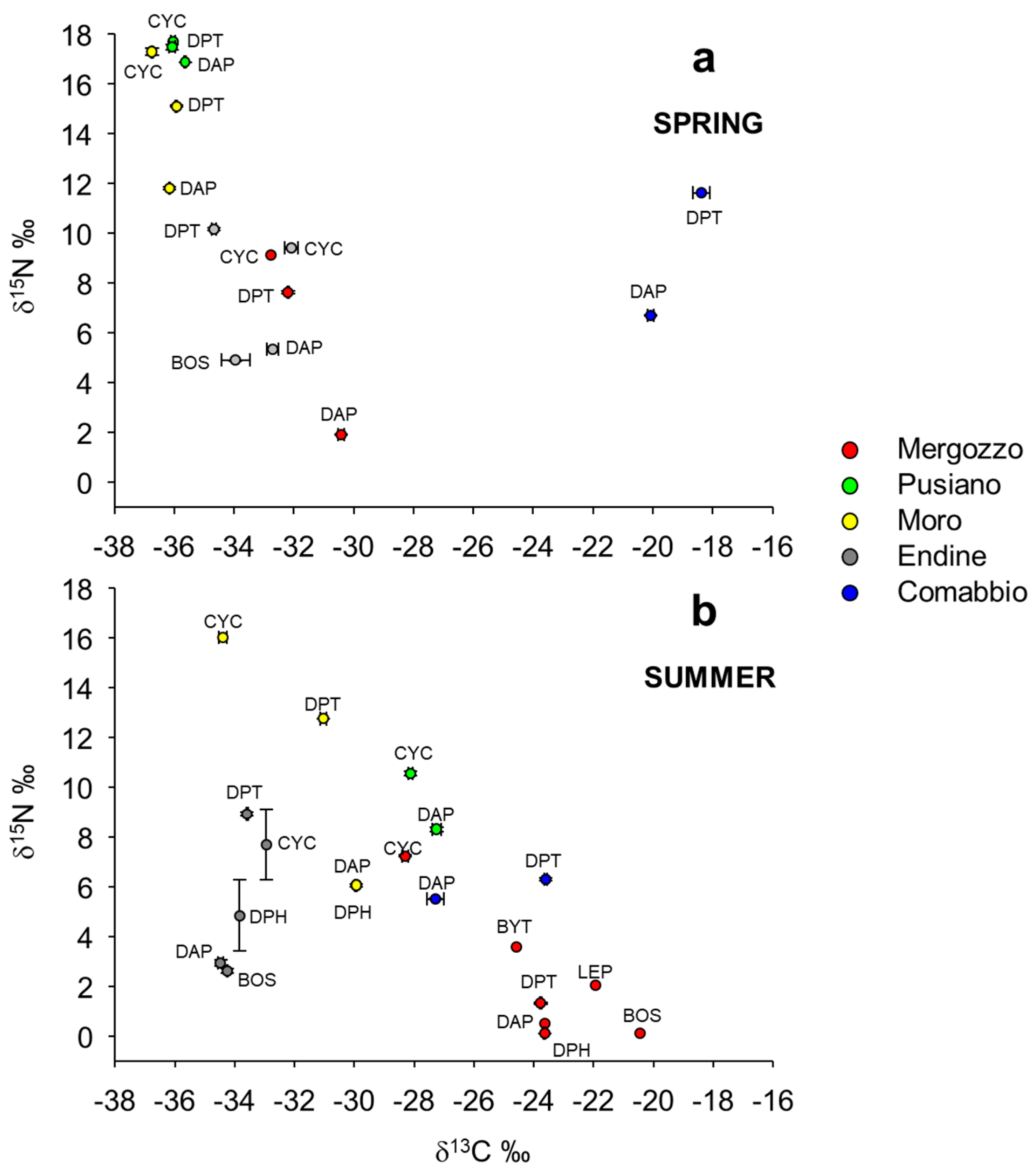
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Middelburg, J.J. Stable isotopes dissect aquatic food webs from the top to the bottom. Biogeosciences 2014, 11, 2357–2371. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Kling, G.W.; Fry, B.; O’Brien, W.J. Stable isotopes and planktonic trophic structure in arctic lakes. Ecology 1992, 73, 561–566. [Google Scholar] [CrossRef]
- Cabana, G.; Rasmussen, J.B. Comparison of aquatic food chains using nitrogen isotopes. Proc. Natl. Acad. Sci. USA 1996, 93, 10844–10847. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Visconti, A.; Manca, M. Seasonal changes in the δ13C and δ15N signatures of the Lago Maggiore pelagic food web. J. Limnol. 2011, 70, 263–271. [Google Scholar] [CrossRef]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. Camb. Philos. Soc. 2012, 87, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Fadda, A.; Manca, M.; Camin, F.; Ziller, L.; Buscarino, P.; Mariani, M.; Padedda, B.M.; Sechi, N.; Virdis, T.; Lugliè, A. Study on the suspended particulate matter of a Mediterranean artificial lake (Sos Canales Lake) using Stable Isotope Analysis of carbon and nitrogen. Ann. Limnol. Int. J. Limnol. 2016, 52, 401–412. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Casselman, J.M.; Rasmussen, J.B. Stable isotope evidence for the food web consequences of species invasions in lakes. Nature 1999, 401, 464–467. [Google Scholar] [CrossRef]
- Grey, J.; Jones, R.I.; Sleep, D. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia 2000, 123, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Manca, M.; Rasmussen, J.B. Peculiarities in the stable isotope composition of organisms from an alpine lake. Aquat. Sci. Res. Bound. 2004, 66, 440–445. [Google Scholar] [CrossRef]
- Visconti, A.; Volta, P.; Fadda, A.; Di Guardo, A.; Manca, M. Seasonality, littoral versus pelagic carbon sources, and stepwise 15N-enrichment of pelagic food web in a deep subalpine lake: The role of planktivorous fish. Can. J. Fish. Aquat. Sci. 2014, 71, 436–446. [Google Scholar] [CrossRef]
- Post, D.M.; Pace, M.L.; Hairston, N.G. Ecosystem size determines food-chain length in lakes. Nature 2000, 405, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- LIMNO Database Della Qualità dei Laghi Italiani. Available online: www.ise.cnr.it/limno/schede (accessed on 3 December 2004).
- Uso del Suolo in Regione Lombardia-Atlante Descrittivo. Available online: http://www.ersaf.lombardia.it/upload/ersaf/gestionedocumentale/AtlanteUsoDelSuolo2010_784_15401.pdf (accessed on 10 May 2012).
- Barbanti, L.; Bonacina, C.; Calderoni, A.; Carollo, A.; de Bernardi, R.; Guilizzoni, P.; Nocentini, A.M.; Ruggiu, D.; Saraceni, C.; Tonolli, L. Indagini Ecologiche sul Lago d’Endine; Edizioni dell’Istituto Italiano di Idrobiologia: Pallanza, Italy, 1974; 304p. [Google Scholar]
- Garibaldi, L.; Brizzio, M.C.; Varallo, A.; Mosello, R. The improving trophic conditions of Lake Endine (Northern Italy). Mem. Ist. Ital. Idrobiol. 1997, 56, 23–36. [Google Scholar]
- Garibaldi, L.; Mosello, R.; Brizzio, M.C.; Varallo, A. Chimica e Fitoplancton del Lago Moro (Alpi Orobiche Bresciane); Picazzo, M., Ed.; Atti A.I.O.L.: Genova, Italy, 2000; Volume 13, pp. 395–410. [Google Scholar]
- Osservatorio dei Laghi Lombardi. Qualità delle acque lacustri in Lombardia. In Primo Rapporto OLL 2004; Osservatorio dei Laghi Lombardi: Brugherio, Italy, 2005; 351p. [Google Scholar]
- Fadda, A.; Rawcliffe, R.; Padedda, B.M.; Luglié, A.; Sechi, N.; Camin, F.; Ziller, L.; Manca, M. Spatiotemporal dynamics of C and N isotopic signature of zooplankton: A seasonal study on a man-made lake in the Mediterranean region. Ann. Limnol. Int. J. Limnol. 2014, 50, 279–287. [Google Scholar] [CrossRef]
- Caroni, R.; Free, G.; Visconti, A.; Manca, M. Phytoplankton functional traits and seston stable isotopes ratio: A functional-based approach in a deep, subalpine lake, Lake Maggiore (N. Italy). J. Limnol. 2012, 71, 84–94. [Google Scholar] [CrossRef]
- Cabana, G.; Rasmussen, J.B. Modeling food chain structure and contaminant bioaccumulation using stable nitrogen isotopes. Nature 1994, 372, 255–257. [Google Scholar] [CrossRef]
- Degens, E.T.; Guillard, R.; Sackett, W.M.; Hellebust, J.A. Metabolic fractionation of carbon isotopes in marine plankton. Temperature and respiration experiments. Deep Sea Res. 1968, 15, 1–9. [Google Scholar] [CrossRef]
- Fry, B.; Wainright, S.C. Diatom sources of 13C-rich carbon in marine food webs. Mar. Ecol. Prog. Ser. 1991, 76, 149–157. [Google Scholar] [CrossRef]
- Zohary, T.; Erez, J.; Gophen, M.; Berman-Frank, I.; Stiller, M. Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnol. Oceanogr. 1994, 39, 1030–1104. [Google Scholar] [CrossRef]
- France, R.L.; Del Giorgio, P.A.; Westcott, K.A. Productivity and heterotrophy influences on zooplankton 13C in northern temperate lakes. Aquat. Microb. Ecol. 1997, 12, 85–93. [Google Scholar] [CrossRef]
- Leggett, M.F.; Servos, M.R.; Hesslein, R.; Johannsson, O.; Millard, E.S.; Dixon, D.G. Biogeochemical influences on the carbon isotope signatures of Lake Ontario biota. Can. J. Fish. Aquat. Sci. 1999, 56, 2211–2218. [Google Scholar] [CrossRef]
- Leggett, M.F.; Johannsson, O.; Hesslein, R.; Dixon, D.G.; Taylor, W.D.; Servos, M.R. Influence of inorganic nitrogen cycling on the δ15N of Lake Ontario biota. Can. J. Fish. Aquat. Sci. 2000, 57, 1489–1496. [Google Scholar] [CrossRef]
- Ruggiu, D.; Morabito, G.; Panzani, P.; Pugnetti, A. Trends and relations among basic phytoplankton characteristics in the course of the long-term oligotrophication of Lake Maggiore (Italy). Hydrobiologia 1998, 369, 243–257. [Google Scholar] [CrossRef]
- Matthews, B.; Mazumder, A. Compositional and interlake variability of zooplankton affect baseline stable isotope signature. Limnol. Oceanogr. 2003, 48, 1977–1987. [Google Scholar] [CrossRef]
- Marty, J.; Planas, D. Comparison of methods to determine algal δ13C in freshwater. Limnol. Oceanogr. Meth. 2008, 6, 51–63. [Google Scholar] [CrossRef]
- Perga, M.E.; Gerdeaux, D. Changes in the δ13C of pelagic food webs: The influence of lake area and trophic status on the isotopic signature of whitefish (Coregonus lavaretus). Can. J. Fish. Aquat. Sci. 2004, 61, 1485–1492. [Google Scholar] [CrossRef]
- Hamza, W.; Ruggiu, D.; Manca, M. Diel zooplankton migrations and their effect on the grazing impact in Lake Candia (Italy). Arch. Hydrobiol. 1993, 39, 175–185. [Google Scholar]
- Syväranta, J.; Haemaelaeinen, H.; Jones, R.I. Within-lake variability in carbon and nitrogen stable isotope signatures. Freshwat. Biol. 2006, 51, 1090–1102. [Google Scholar] [CrossRef]
- Grey, J.; Jones, R.I.; Sleep, D. Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol. Oceanogr. 2001, 46, 505–513. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- Santer, B.; Sommerwerk, N.; Grey, J. Food niches of cyclopoid copepods in eutrophic Plußsee determined by stable isotope analysis. Arch. Hydrobiol. 2006, 167, 301–316. [Google Scholar] [CrossRef]
- Van den Bosch, F.; Santer, B. Cannibalism in Cyclops abyssorum. Oikos 1993, 67, 19–28. [Google Scholar] [CrossRef]
- Santer, B.; Van den Bosch, F. Herbivorous nutrition of Cyclops vicinus: The effect of a pure algal diet on feeding, development, reproduction and life cycle. J. Plankton Res. 1994, 16, 171–195. [Google Scholar] [CrossRef]
- De Bernardi, R.; Soldavini, E. Long-term fluctuations of zooplankton in Lake Mergozzo, Northern Italy. Mem. Ist. Ital. Idrobiol. 1976, 33, 345–375. [Google Scholar]
- De Bernardi, R.; Soldavini, E. Seasonal dynamics of the zooplankton community in Lago di Mergozzo (Northern Italy). Mem. Ist. Ital. Idrobiol. 1977, 34, 137–154. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Altitude (m a.s.l.) | Geographic Coordinates (Latitude Longitude) | Mixing Regime | Volume (106 m3) | Depthmax (m) | Ptot (µg L−1) |
|---|---|---|---|---|---|---|
| Mergozzo | 194 | 45°57′20″ N 8°27′52″ E | monomictic | 83 | 73 | 1 |
| Moro | 389 | 45°52′47″ N 10°09′25″ E | meromictic | 4 | 42 | 8 |
| Endine | 334 | 45°46′41″ N 9°56′22″ E | dimictic | 12 | 9 | 17 |
| Comabbio | 243 | 45°45′55″ N 8°41′37″ E | polymictic | 16 | 8 | 72 |
| Pusiano | 259 | 45°48′09″ N 9°16′17″ E | monomictic | 69 | 24 | 74 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piscia, R.; Boggio, E.; Bettinetti, R.; Mazzoni, M.; Manca, M. Carbon and Nitrogen Isotopic Signatures of Zooplankton Taxa in Five Small Subalpine Lakes along a Trophic Gradient. Water 2018, 10, 94. https://doi.org/10.3390/w10010094
Piscia R, Boggio E, Bettinetti R, Mazzoni M, Manca M. Carbon and Nitrogen Isotopic Signatures of Zooplankton Taxa in Five Small Subalpine Lakes along a Trophic Gradient. Water. 2018; 10(1):94. https://doi.org/10.3390/w10010094
Chicago/Turabian StylePiscia, Roberta, Emanuela Boggio, Roberta Bettinetti, Michela Mazzoni, and Marina Manca. 2018. "Carbon and Nitrogen Isotopic Signatures of Zooplankton Taxa in Five Small Subalpine Lakes along a Trophic Gradient" Water 10, no. 1: 94. https://doi.org/10.3390/w10010094
APA StylePiscia, R., Boggio, E., Bettinetti, R., Mazzoni, M., & Manca, M. (2018). Carbon and Nitrogen Isotopic Signatures of Zooplankton Taxa in Five Small Subalpine Lakes along a Trophic Gradient. Water, 10(1), 94. https://doi.org/10.3390/w10010094