
Effects of Stratospheric Conditions on the Viability, Metabolism and Proteome of Prokaryotic Cells

 ,
, 

Abstract
:1. Introduction
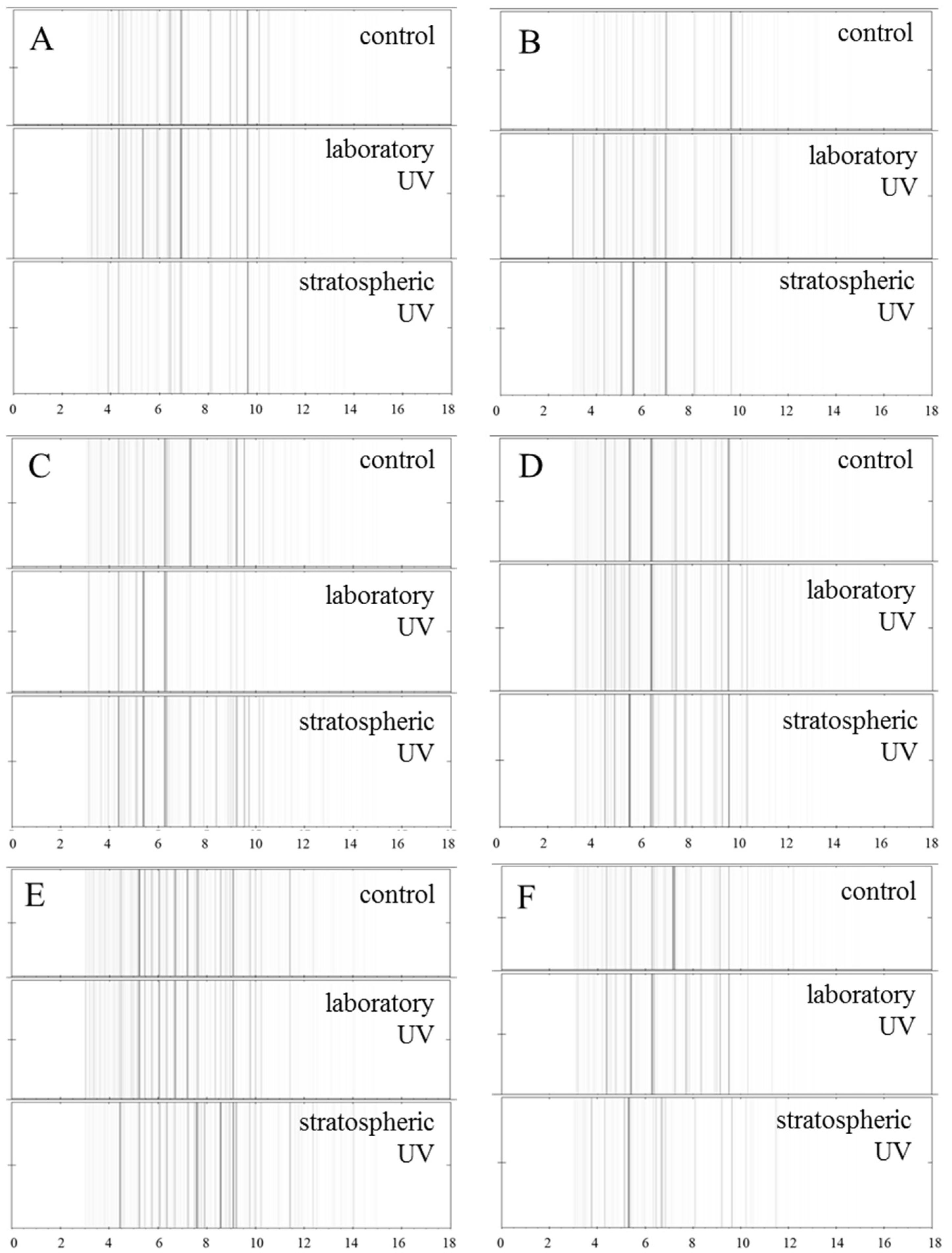
2. Results and Discussion
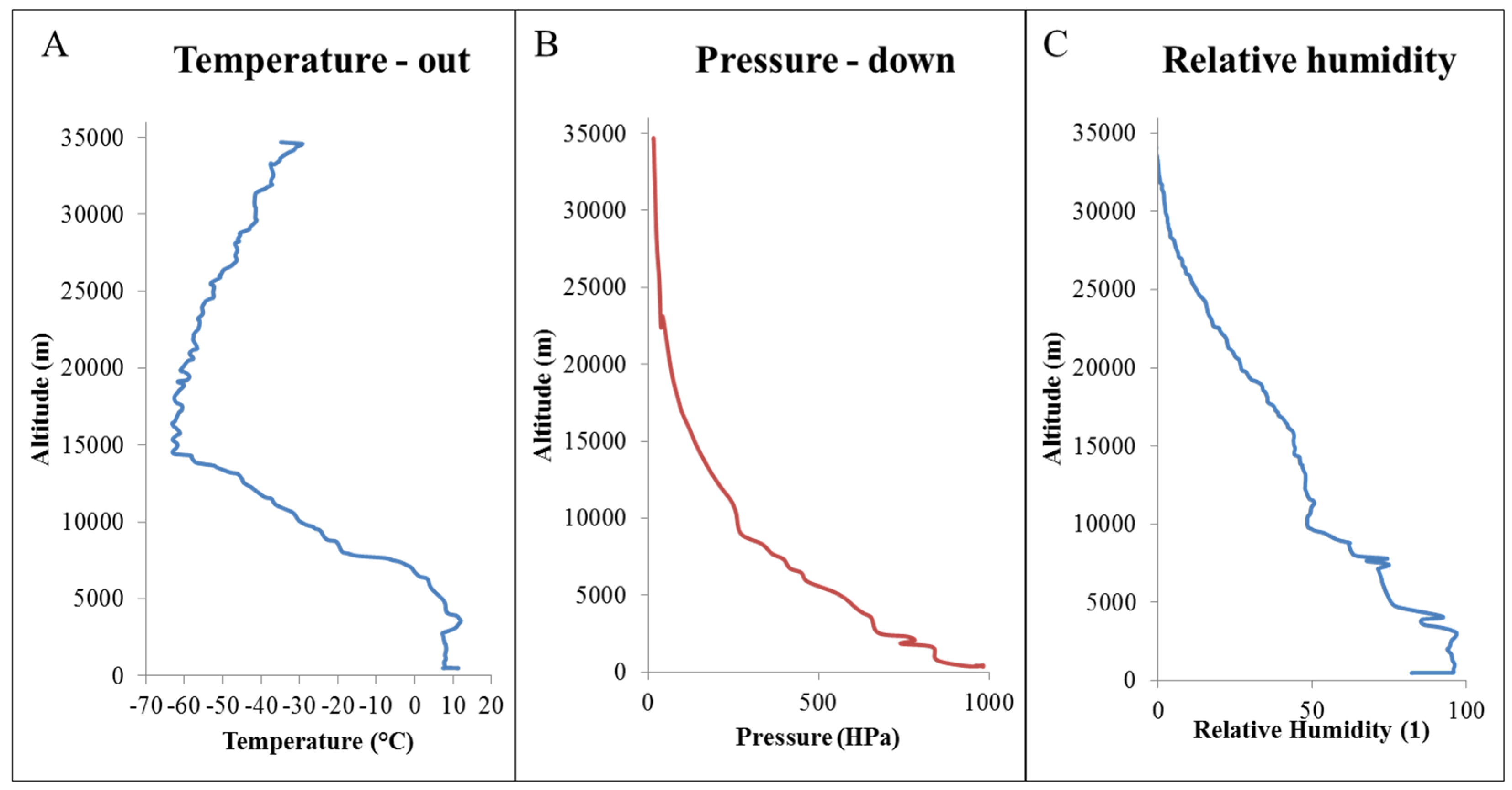
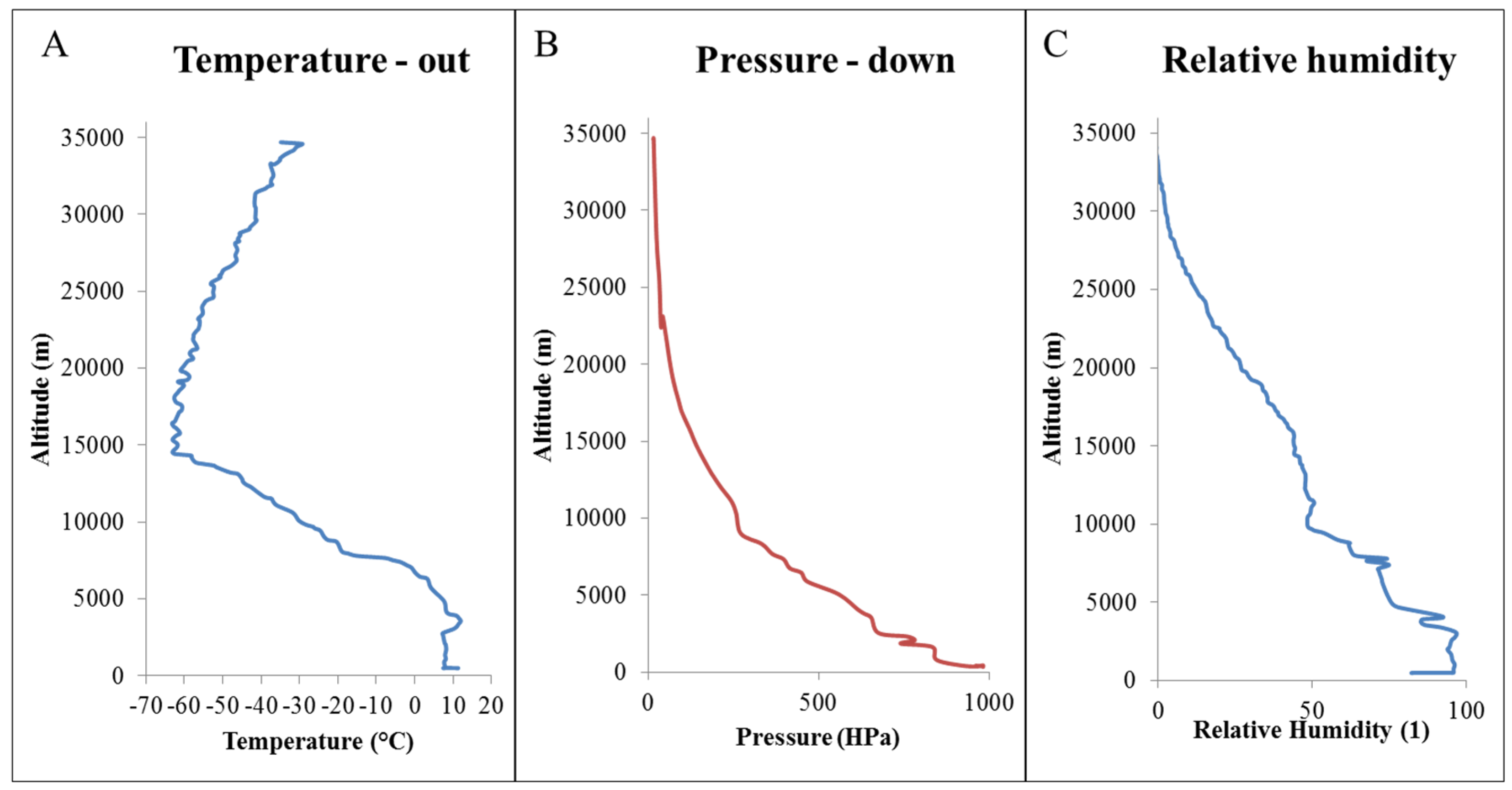
2.1. Atmospheric Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date, Time and Place of Flight | Climatic Data on the Earth’s Surface | ||||||
|---|---|---|---|---|---|---|---|
| 1 May 2015 Spisska Nova Ves 7:45 a.m.–9:45 a.m. | sunshine | daily air temperature | air pressure (hPa) | atmospheric precipitations | |||
| sunrise | sunset | / | daily minimum air pressure | daily maximum air pressure | average daily air pressure | / | |
| 4:18 | 18:47 | 13 °C | 980 | 1015 | 997 | 46 | |
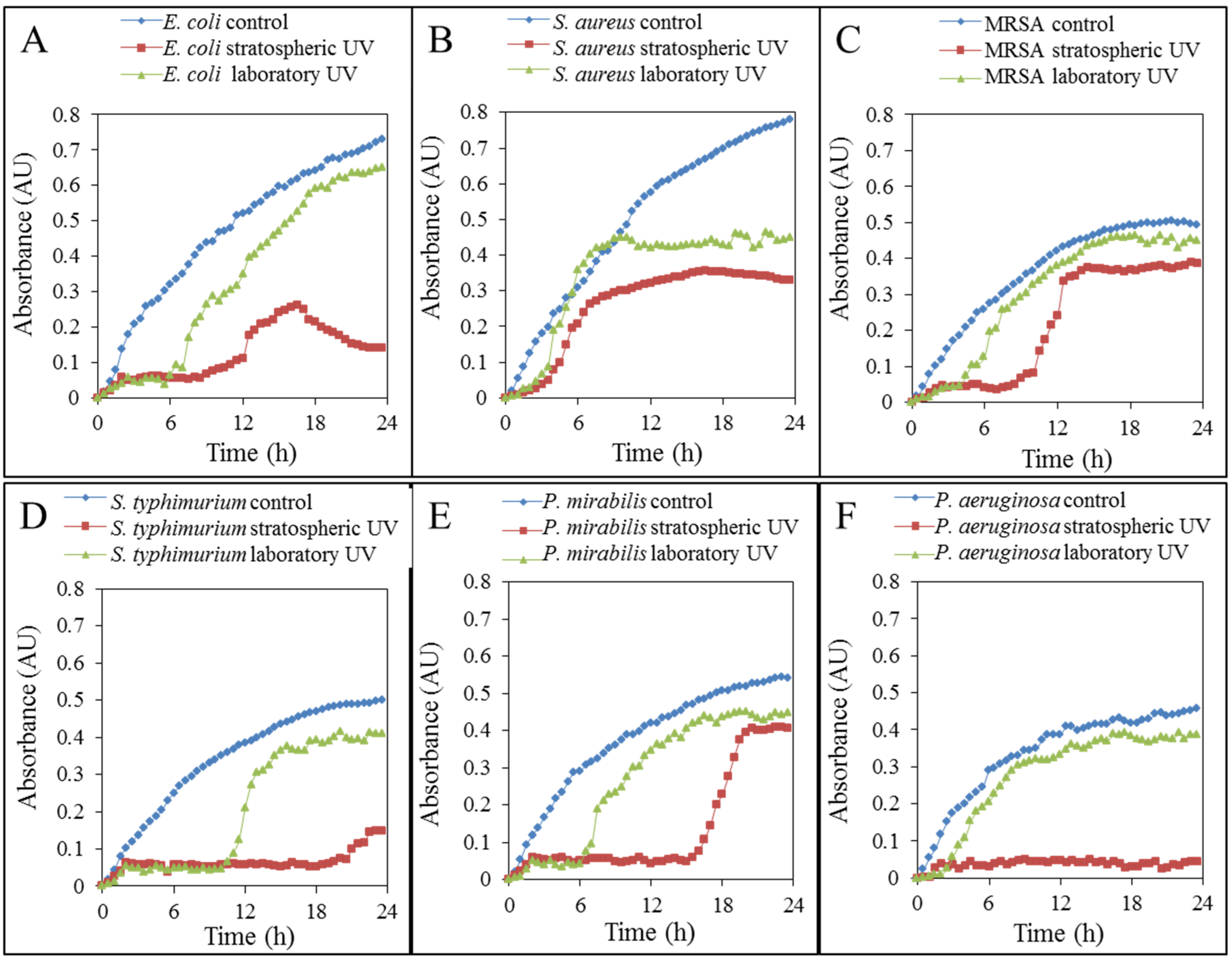
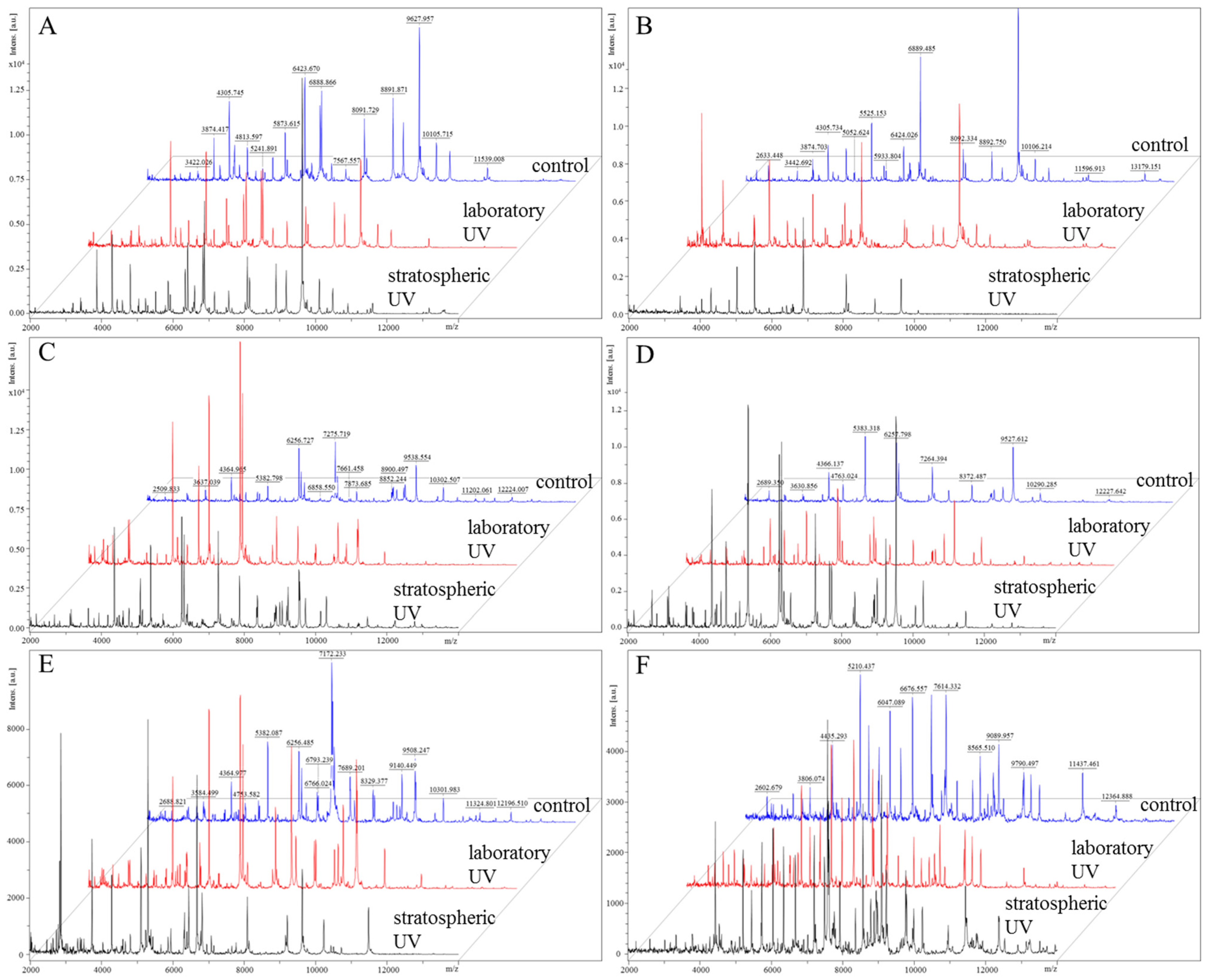
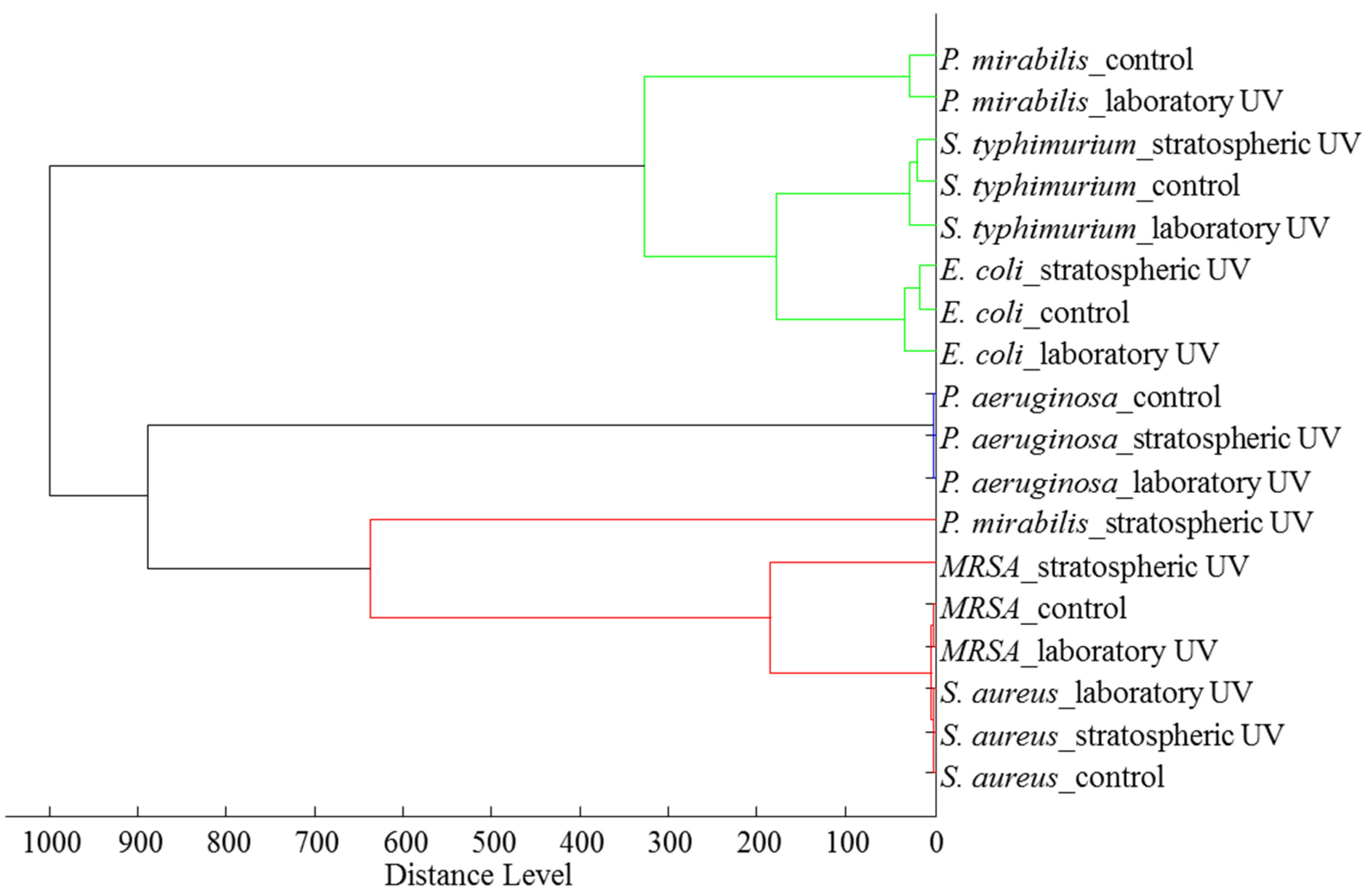
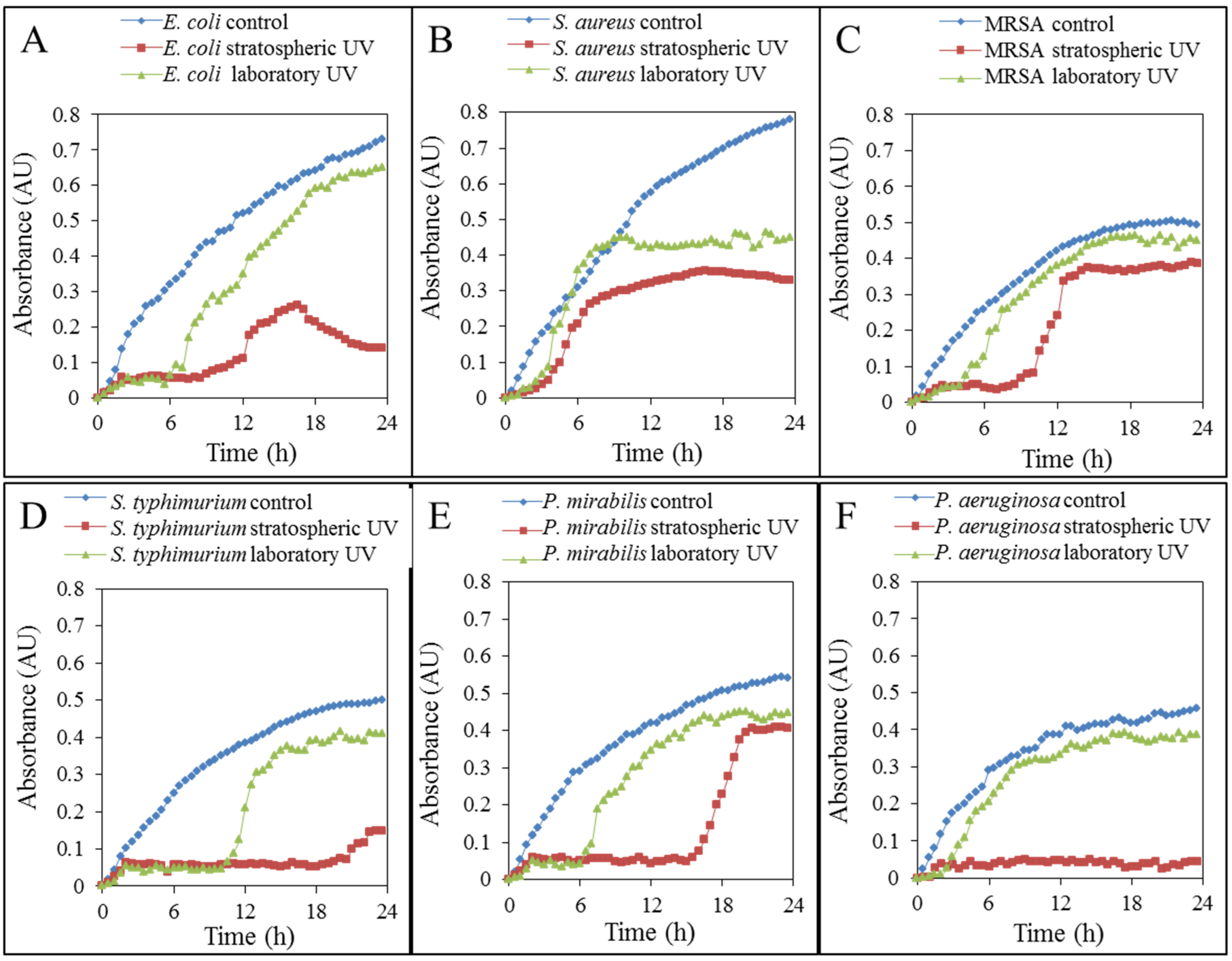
2.2. Effects on Bacteria
2.2.1. Growth Properties
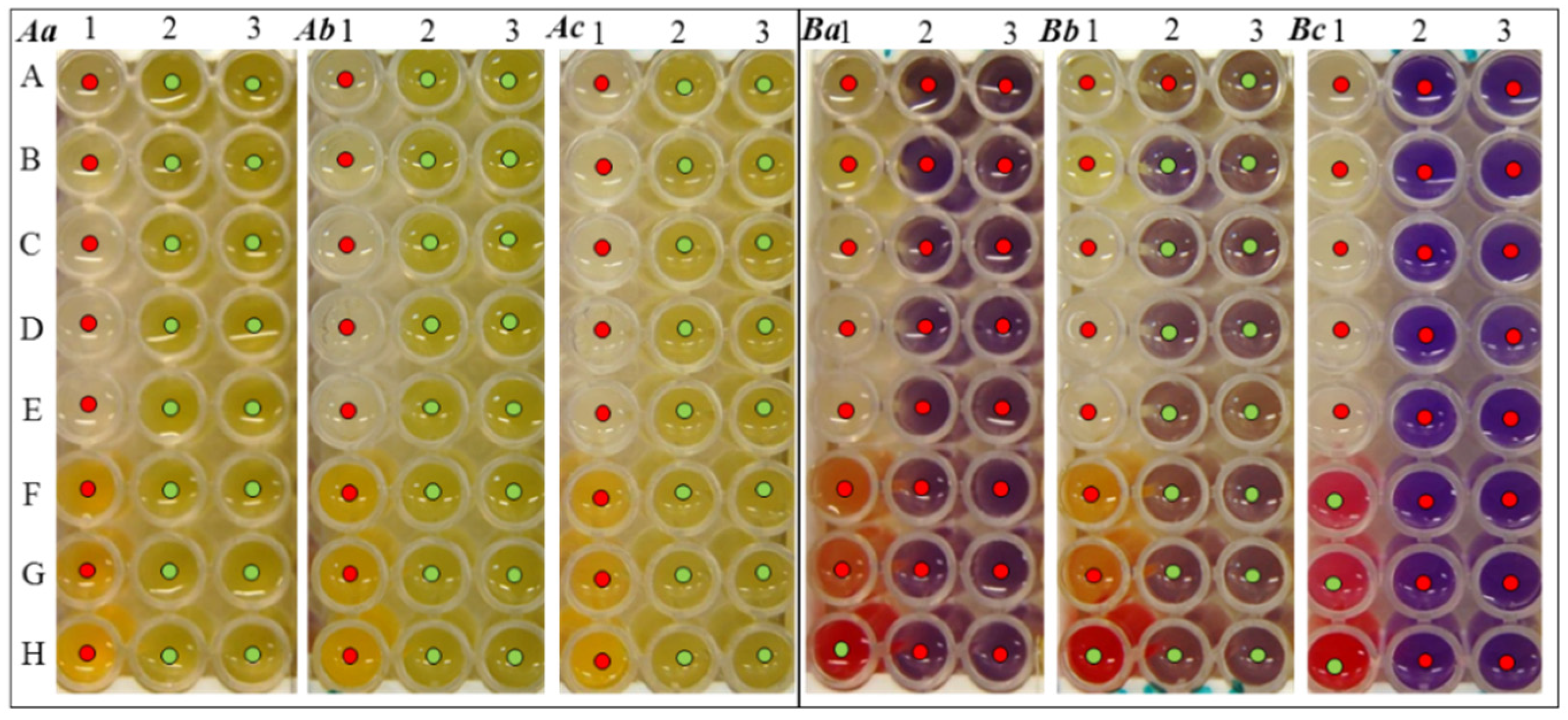
2.2.2. Resistance
| Bacterial Culture | Type of Exposure | The Size of The Inhibition Zone (mm) | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| ERY | PNC | AMX | TTC | LNC | SeNPs | ||
| S. aureus | control | 5 | 1 | 3 | 10 | 10 | 5 |
| laboratory UV | 11 | 5 | 10 | 11 | 11 | 7 | |
| stratospheric UV | 11 | 6 | 11 | 15 | 12 | 7 | |
| MRSA | control | 0 | 0 | 1 | 5 | 0 | 0 |
| laboratory UV | 0 | 0 | 2 | 6 | 0 | 8 | |
| stratospheric UV | 0 | 0 | 2 | 7 | 0 | 9 | |
| E. coli | control | 0 | 0 | 5 | 5 | 0 | 0 |
| laboratory UV | 0 | 0 | 8 | 7 | 0 | 0 | |
| stratospheric UV | 0 | 0 | 11 | 10 | 0 | 6 | |
| S. typhimurium | control | 2 | 0 | 0 | 8 | 0 | 0 |
| laboratory | 4 | 0 | 0 | 9 | 0 | 0 | |
| stratospheric UV | 7 | 4 | 7 | 14 | 2 | 0 | |
| P. mirabilis | control | 0 | 0 | 0 | 3 | 0 | 0 |
| laboratory UV | 0 | 0 | 0 | 5 | 0 | 0 | |
| stratospheric UV | 2 | 0 | 2 | 7 | 0 | 0 | |
| P. aeruginosa | control | 0 | 0 | 0 | 4 | 0 | 0 |
| laboratory UV | 0 | 0 | 5 | 5 | 0 | 0 | |
| stratospheric UV | 2 | 4 | 7 | 5 | 1 | 3 | |
2.2.3. Biochemical Markers


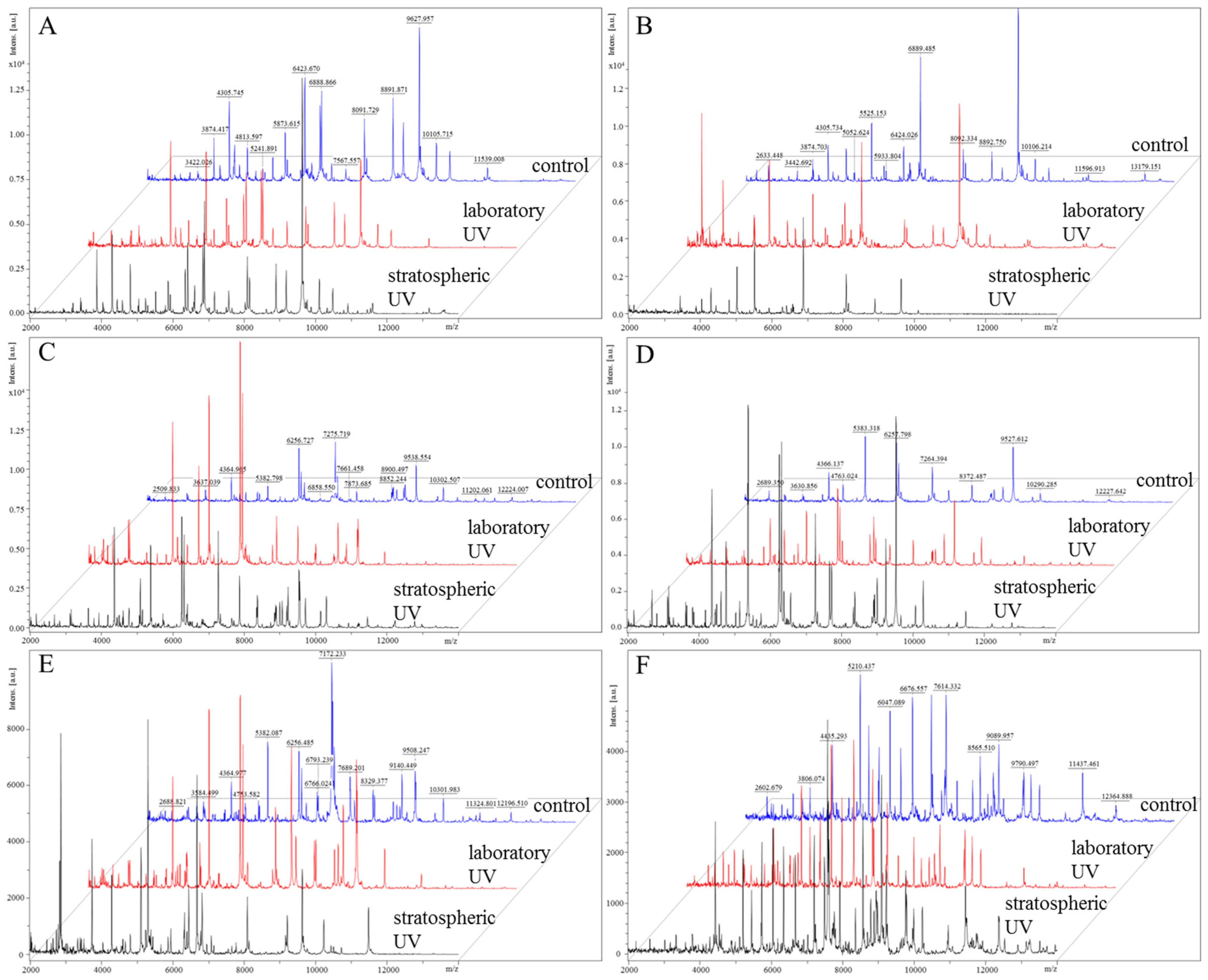
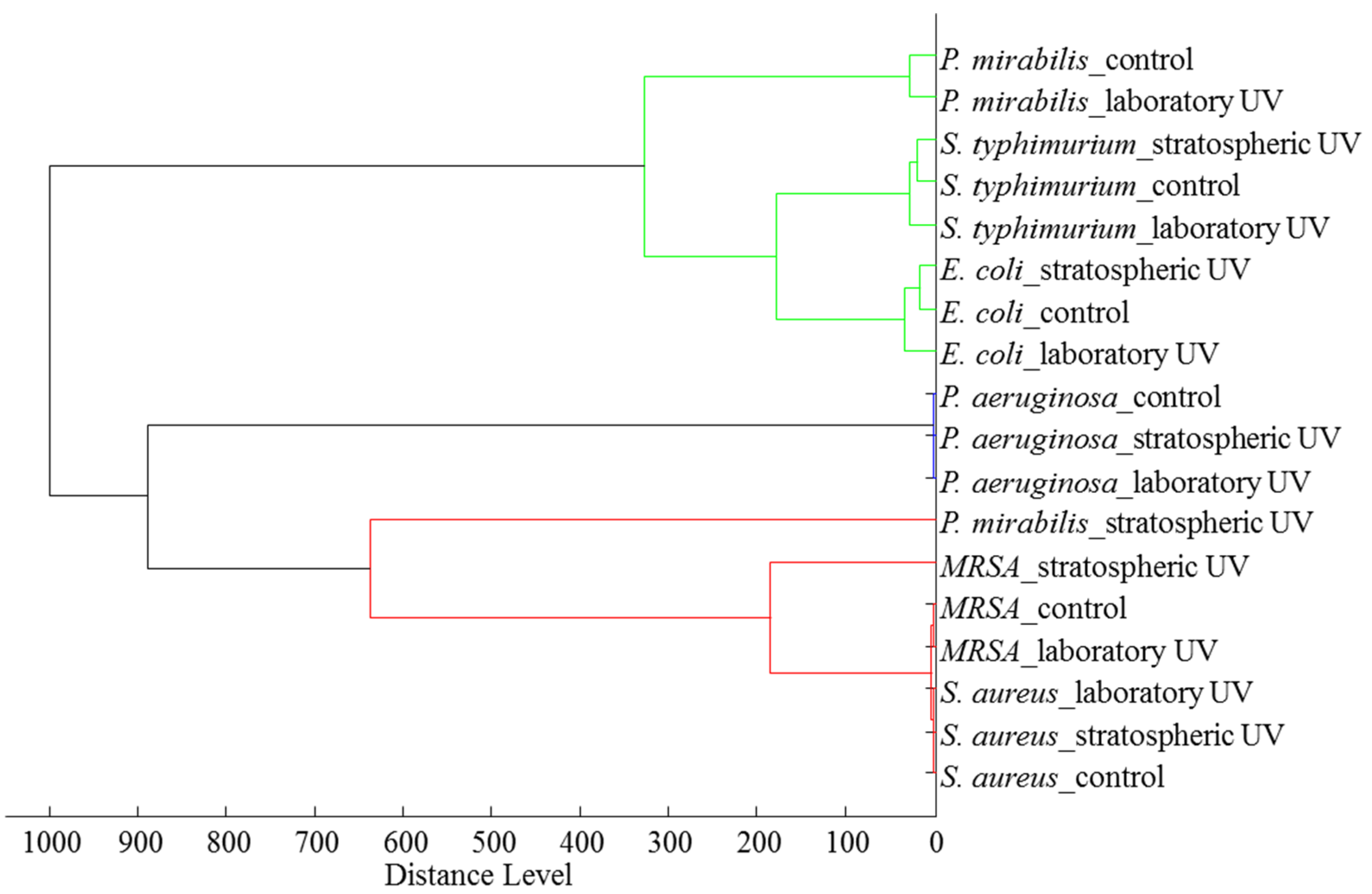
2.2.4. Proteomic Analyses
3. Experimental Section
3.1. Chemicals
3.2. Lyophilization of Bacterial Cultures
3.3. The Experimental Conditions
Exposure of Bacterial Cultures to UV Radiation in the Stratosphere
3.4. Measurement of Climatic Conditions
3.5. Cultivation of Bacterial Species
3.5.1. Determination of Growth Curves
3.5.2. Determination of Bacterial Strain Resistance to Antibiotic Drugs
3.6. Mass Spectrometry
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mone, M.J.; Volker, M.; Nikaido, O.; Mullenders, L.H.F.; van Zeeland, A.A.; Verschure, P.J.; Manders, E.M.M.; van Driel, R. Local UV induced DNA damage in cell nuclei results in local transcription inhibition. EMBO Rep. 2001, 2, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.A.; Bounty, S.; Linden, K.G. Long-range quantitative PCR for determining inactivation of adenovirus 2 by ultraviolet light. J. Appl. Microbiol. 2013, 114, 1854–1865. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Hader, D.P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated base damage to cellular DNA. Free Radic. Biol. Med. 2010, 49, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet radiation-mediated damage to cellular DNA. Mutat. Res. 2005, 571, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Douki, T. The variety of UV-induced pyrimidine dimeric photoproducts in DNA as shown by chromatographic quantification methods. Photochem. Photobiol. Sci. 2013, 12, 1286–1302. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, C. The universe: A cryogenic habitat for microbial life. Cryobiology 2004, 48, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Shivaji, S.; Chaturvedi, P.; Suresh, K.; Reddy, G.S.N.; Dutt, C.B.S.; Wainwright, M.; Narlikar, J.V.; Bhargava, P.M. Bacillus aerius sp nov., Bacillus aerophilus sp nov., Bacillus stratosphericus sp nov and Bacillus altitudinis sp nov., isolated from cryogenic tubes used for collecting air samples from high altitudes. Int. J. Syst. Evol. Microbiol. 2006, 56, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Griffin, D.W.; Schuerger, A.C. Stratospheric microbiology at 20 km over the Pacific Ocean. Aerobiologia 2010, 26, 35–46. [Google Scholar] [CrossRef]
- Griffin, D.W. Terrestrial microorganisms at an altitude of 20,000 m in Earth’s atmosphere. Aerobiologia 2004, 20, 135–140. [Google Scholar] [CrossRef]
- Wainwright, M.; Wickramasinghe, N.C.; Narlikar, J.V.; Rajaratnam, P.; Perkins, J. Bacteria in the stratosphere-confirmation of viable but non-cultureable forms. In Instruments, Methods, and Missions for Astrobiology VII; Hoover, R.B., Rozanov, A.Y., Eds.; Spie-Int Soc Optical Engineering: Bellingham, USA, 2004; Vol. 5163, pp. 218–221. [Google Scholar]
- Imshenetsky, A.A.; Lysenko, S.V.; Kazakov, G.A. Upper boundary of biosphere. Appl. Environ. Microbiol. 1978, 35, 1–5. [Google Scholar] [PubMed]
- Papke, R.T.; Ward, D.M. The importance of physical isolation to microbial diversification. FEMS Microbiol. Ecol. 2004, 48, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Griffin, D.W.; McPeters, R.D.; Ward, P.D.; Schuerger, A.C. Microbial survival in the stratosphere and implications for global dispersal. Aerobiologia 2011, 27, 319–332. [Google Scholar] [CrossRef]
- Thoma, F. Light and dark in chromatin repair: Repair of UV-induced DNA lesions by photolyase and nucleotide excision repair. EMBO J. 1999, 18, 6585–6598. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.C.; Stewart, M.; Granger, D.; Guzik, T.G.; Christner, B.C. A method for sampling microbial aerosols using high altitude balloons. J. Microbiol. Methods 2014, 107, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Yudin, V.; Khatattov, B. Introduction to atmospheric chemistry and constituent transport. In Data Assimilation; Lahoz, M., Khattatov, B., Eds.; Springer-Verlag Berlin: Berlin, France, 2010; pp. 409–430. [Google Scholar]
- Chudobova, D.; Cihalova, K.; Skalickova, S.; Zitka, J.; Rodrigo, M.A.M.; Milosavljevic, V.; Hynek, D.; Kopel, P.; Vesely, R.; Adam, V.; et al. 3D-printed chip for detection of methicillin-resistant Staphylococcus aureus labeled with gold nanoparticles. Electrophoresis 2015, 36, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Heger, Z.; Zitka, J.; Cernei, N.; Krizkova, S.; Sztalmachova, M.; Kopel, P.; Masarik, M.; Hodek, P.; Zitka, O.; Adam, V.; et al. 3D-printed biosensor with poly (dimethylsiloxane) reservoir for magnetic separation and quantum dots-based immunolabeling of metallothionein. Electrophoresis 2015, 36, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Krejcova, L.; Nejdl, L.; Rodrigo, M.A.M.; Zurek, M.; Matousek, M.; Hynek, D.; Zitka, O.; Kopel, P.; Adam, V.; Kizek, R. 3D printed chip for electrochemical detection of influenza virus labeled with CdS quantum dots. Biosens. Bioelectron. 2014, 54, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Vlachova, J.; Tmejova, K.; Kopel, P.; Korabik, M.; Zitka, J.; Hynek, D.; Kynicky, J.; Adam, V.; Kizek, R. A 3D microfluidic chip for electrochemical detection of hydrolysed nucleic bases by a modified glassy carbon electrode. Sensors 2015, 15, 2438–2452. [Google Scholar] [CrossRef] [PubMed]
- Poepping, C.; Beck, S.E.; Wright, H.; Linden, K.G. Evaluation of DNA damage reversal during medium-pressure UV disinfection. Water Res. 2014, 56, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Escudero, L.; Chong, G.; Demergasso, C.; Farias, M.E.; Cabrol, N.A.; Grin, E.; Minkley, E.; Yu, Y. Investigating microbial diversity and UV radiation impact at the high-altitude Lake Aguas Calientes, Chile. In Instruments, Methods, and Missions for Astrobiology X; Hoover, R.B., Levin, G.V., Rozanov, A.Y., Davies, P.C.W., Eds.; Spie-Int Soc Optical Engineering: Bellingham, USA, 2007; Vol. 6694, pp. 236–245. [Google Scholar]
- Chudobova, D.; Dostalova, S.; Blazkova, I.; Michalek, P.; Ruttkay-Nedecky, B.; Sklenar, M.; Nejdl, L.; Kudr, J.; Gumulec, J.; Tmejova, K.; et al. Effect of ampicillin, streptomycin, penicillin and tetracycline on metal resistant and non-resistant staphylococcus aureus. Int. J. Environ. Res. Public Health 2014, 11, 3233–3255. [Google Scholar] [CrossRef] [PubMed]
- Chudobova, D.; Cihalova, K.; Dostalova, S.; Ruttkay-Nedecky, B.; Rodrigo, M.A.M.; Tmejova, K.; Kopel, P.; Nejdl, L.; Kudr, J.; Gumulec, J.; et al. Comparison of the effects of silver phosphate and selenium nanoparticles on Staphylococcus aureus growth reveals potential for selenium particles to prevent infection. FEMS Microbiol. Lett. 2014, 351, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Gomes, N.C.M.; Henriques, I.; Almeida, A.; Correia, A.; Cunha, A. Contribution of reactive oxygen species to UV-B-induced damage in bacteria. J. Photochem. Photobiol. B 2012, 117, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Moreirinha, C.; Lopes, D.; Esteves, A.C.; Henriques, I.; Almeida, A.; Domingues, M.R.M.; Delgadillo, I.; Correia, A.; Cunha, A. Effects of UV radiation on the lipids and proteins of bacteria studied by mid-infrared spectroscopy. Environ. Sci. Technol. 2013, 47, 6306–6315. [Google Scholar] [CrossRef] [PubMed]
- Chudobova, D.; Dostalova, S.; Ruttkay-Nedecky, B.; Guran, R.; Rodrigo, M.A.M.; Tmejova, K.; Krizkova, S.; Zitka, O.; Adam, V.; Kizek, R. The effect of metal ions on Staphylococcus aureus revealed by biochemical and mass spectrometric analyses. Microbiol. Res. 2015, 170, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Chudobova, D.; Nejdl, L.; Gumulec, J.; Krystofova, O.; Rodrigo, M.A.M.; Kynicky, J.; Ruttkay-Nedecky, B.; Kopel, P.; Babula, P.; Adam, V.; et al. Complexes of silver(I) ions and silver phosphate nanoparticles with hyaluronic acid and/or chitosan as promising antimicrobial agents for vascular Grafts. Int. J. Mol. Sci. 2013, 14, 13592–13614. [Google Scholar] [CrossRef] [PubMed]
- Sauer, S.; Freiwald, A.; Maier, T.; Kube, M.; Reinhardt, R.; Kostrzewa, M.; Geider, K. Classification and identification of bacteria by mass spectrometry and computational analysis. PLoS ONE 2008, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chudobova, D.; Cihalova, K.; Jelinkova, P.; Zitka, J.; Nejdl, L.; Guran, R.; Klimanek, M.; Adam, V.; Kizek, R. Effects of Stratospheric Conditions on the Viability, Metabolism and Proteome of Prokaryotic Cells. Atmosphere 2015, 6, 1290-1306. https://doi.org/10.3390/atmos6091290
Chudobova D, Cihalova K, Jelinkova P, Zitka J, Nejdl L, Guran R, Klimanek M, Adam V, Kizek R. Effects of Stratospheric Conditions on the Viability, Metabolism and Proteome of Prokaryotic Cells. Atmosphere. 2015; 6(9):1290-1306. https://doi.org/10.3390/atmos6091290
Chicago/Turabian StyleChudobova, Dagmar, Kristyna Cihalova, Pavlina Jelinkova, Jan Zitka, Lukas Nejdl, Roman Guran, Martin Klimanek, Vojtech Adam, and Rene Kizek. 2015. "Effects of Stratospheric Conditions on the Viability, Metabolism and Proteome of Prokaryotic Cells" Atmosphere 6, no. 9: 1290-1306. https://doi.org/10.3390/atmos6091290
APA StyleChudobova, D., Cihalova, K., Jelinkova, P., Zitka, J., Nejdl, L., Guran, R., Klimanek, M., Adam, V., & Kizek, R. (2015). Effects of Stratospheric Conditions on the Viability, Metabolism and Proteome of Prokaryotic Cells. Atmosphere, 6(9), 1290-1306. https://doi.org/10.3390/atmos6091290