Molecular Cloning and Effects of Tm14-3-3ζ-Silencing on Larval Survivability Against E. coli and C. albicans in Tenebrio molitor


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Microbial Cultures and Bioassays
2.3. Isolation of Tm14-3-3ζ Full-Length Complementary DNA Sequence
2.4. Protein Domain and Phylogenetic Analysis
2.5. Quantitative Polymerase Chain Reaction Analysis
2.6. Generation of Peptide-Based Tm14-3-3ζ Antiserum and Reactivity Studies
2.7. Subcellular Localization of Tm14-3-3ζ by Immunofluorescence
2.8. Double Stranded RNA Treatment and Gene Silencing
2.9. Mortality Assay
3. Results
3.1. Cloning and Sequence Analysis of Tm14-3-3ζ Gene
3.2. Sequence Homology and Phylogenetic Analysis
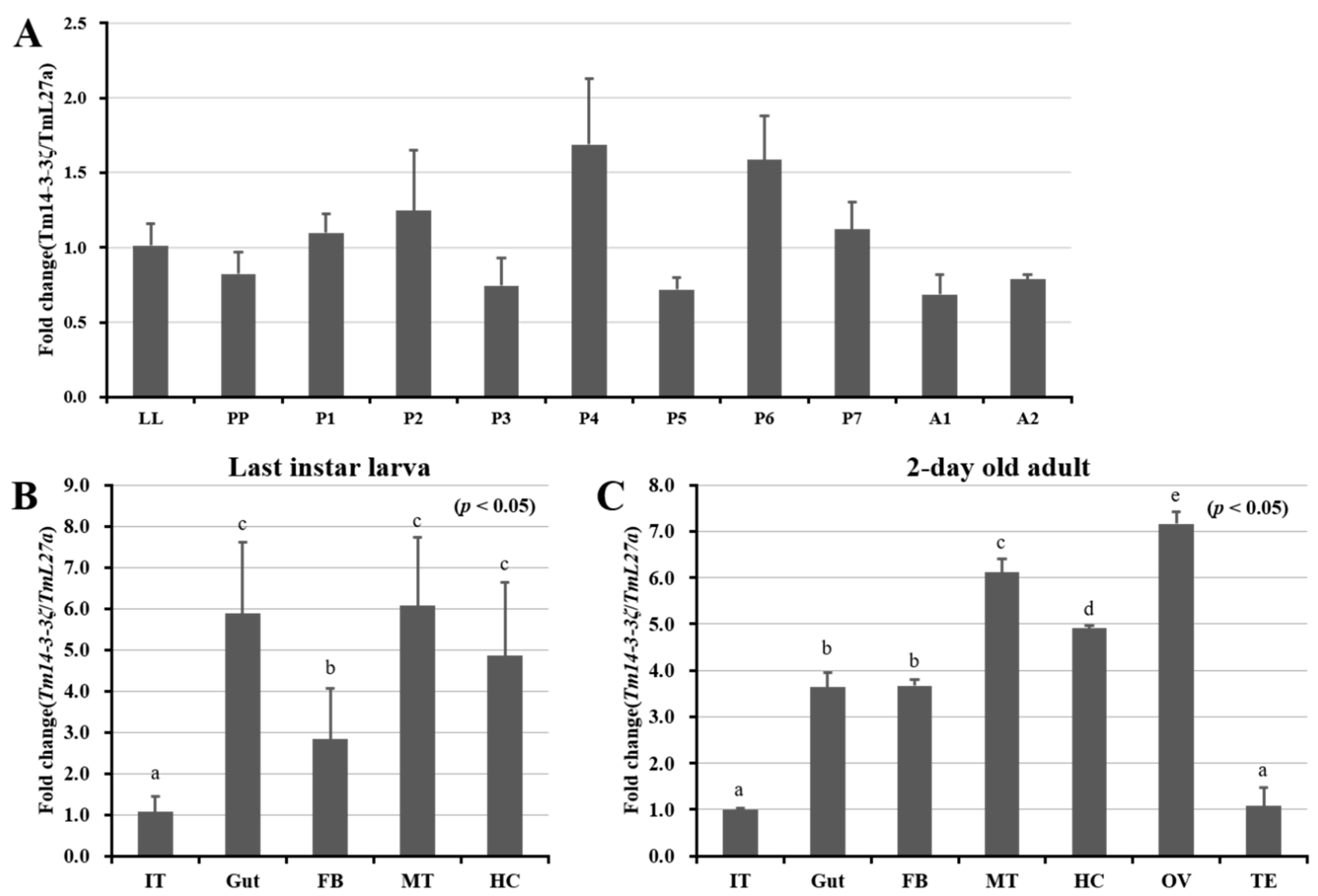
3.3. Developmental and Tissue Specific Expression Patterns of Tm14-3-3ζ
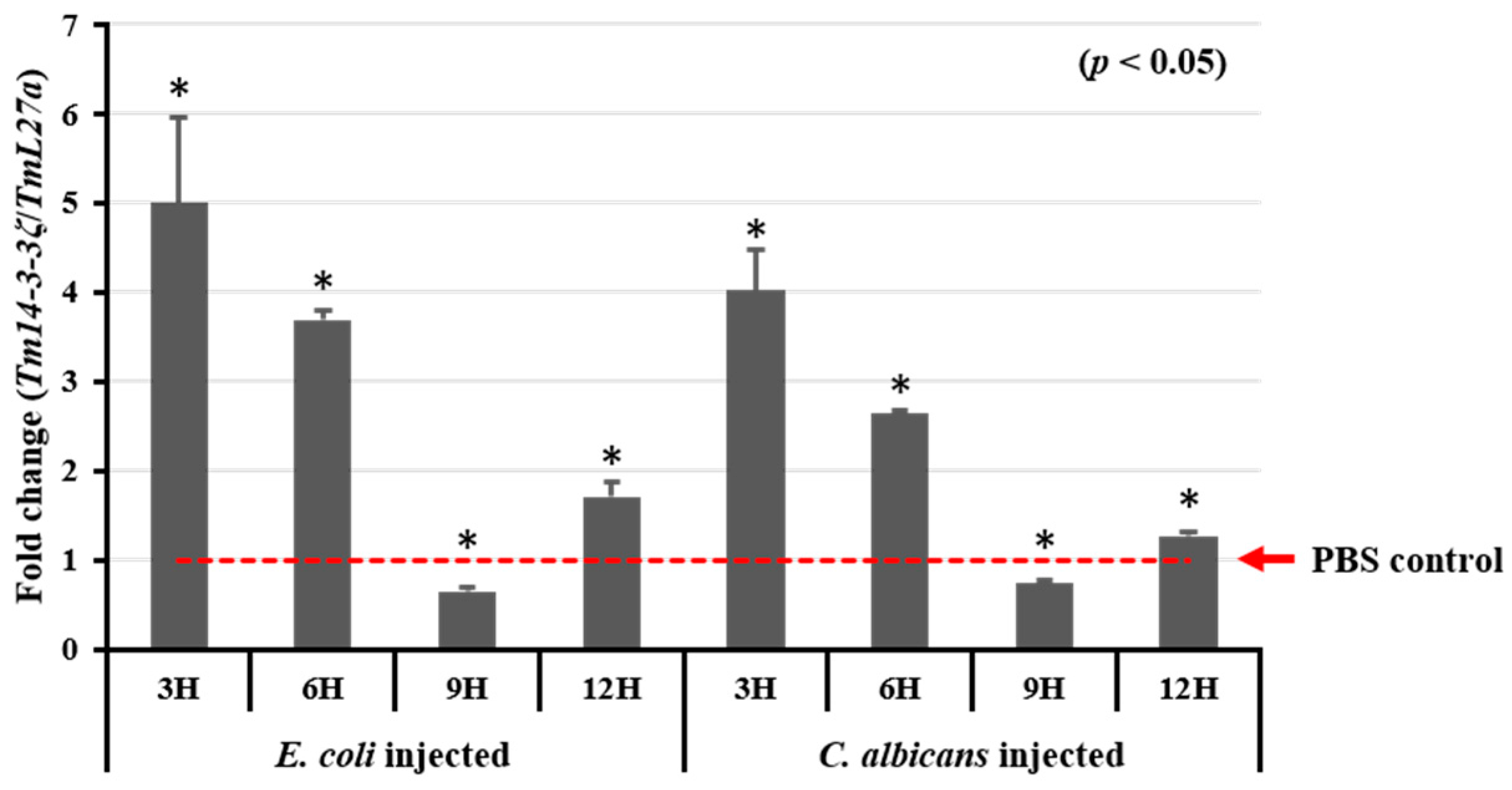
3.4. Expression Analysis of Tm14-3-3ζ in Response to Microorganisms
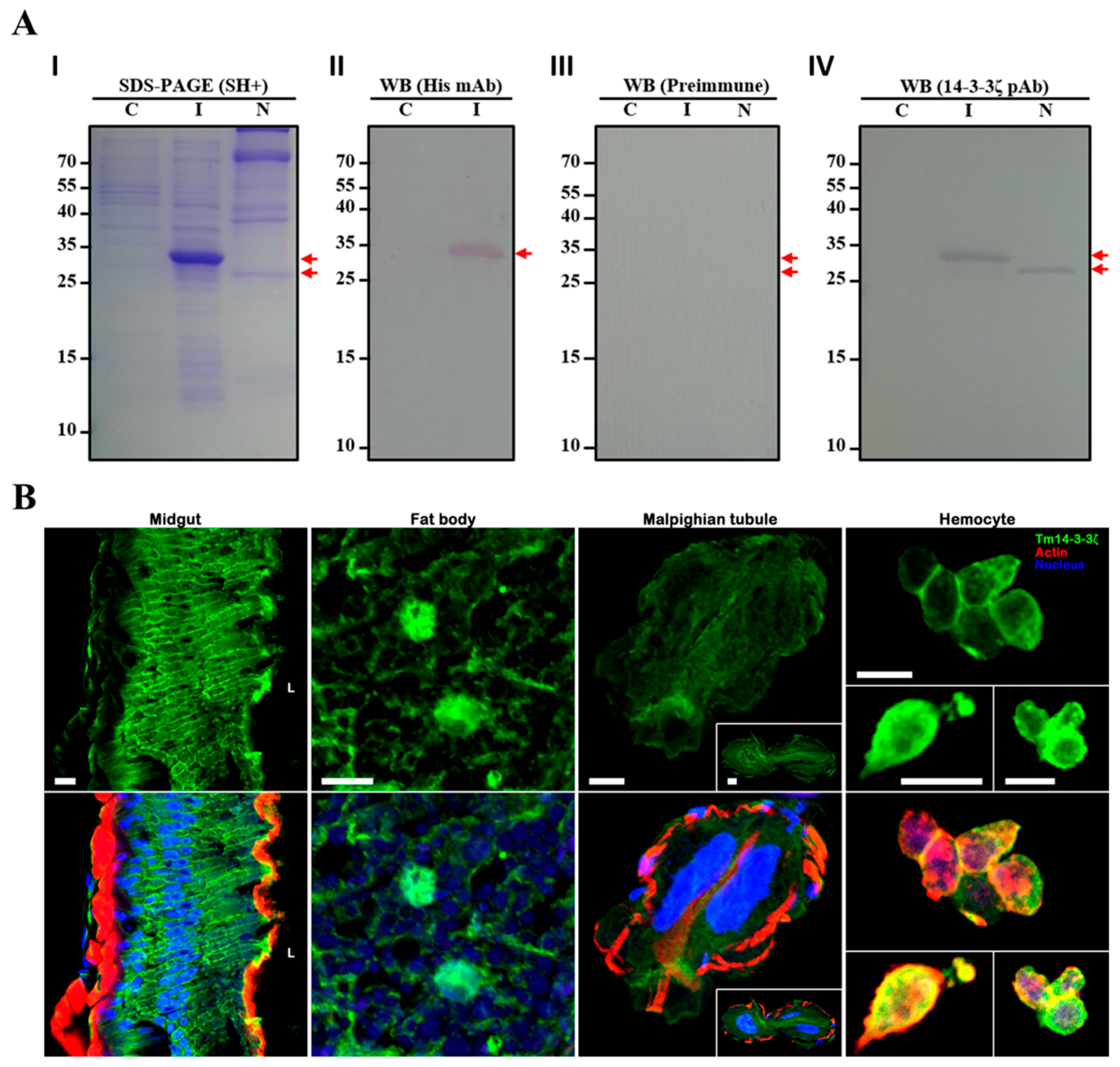
3.5. Characterization of Polyclonal Antibody and Sub-Cellular Localization of Tm14-3-3ζ
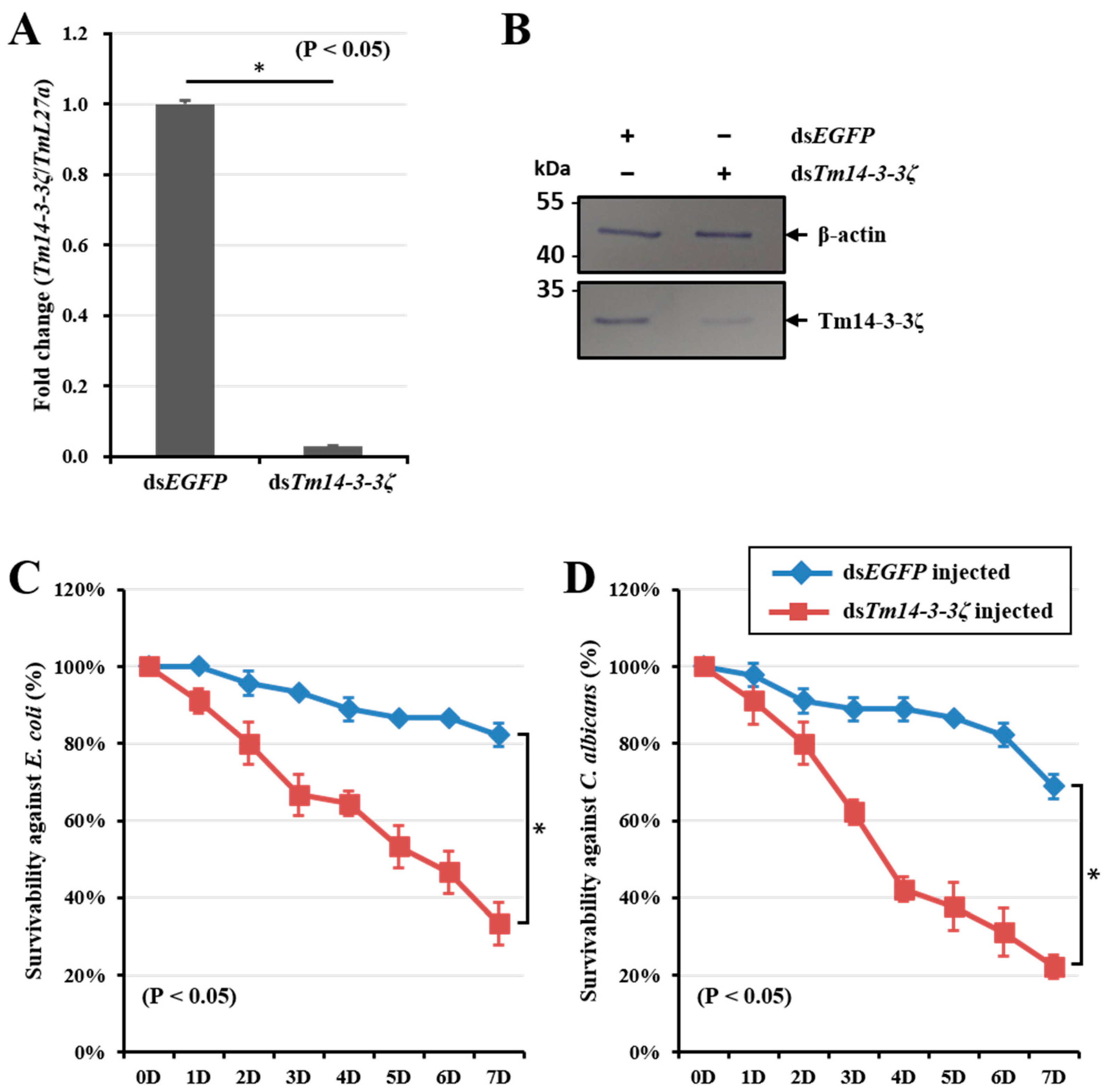
3.6. Tm14-3-3ζ Silencing Increases Susceptibility to Escherichia coli and Candida albicans Infection in Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cao, W.; Yang, X.; Zhou, J.; Teng, Z.; Cao, L.; Zhang, X.; Fei, Z. Targeting 14-3-3 protein, difopein induces apoptosis of human glioma cells and suppresses tumor growth in mice. Apoptosis 2010, 15, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Pozuelo-Rubio, M. 14-3-3 proteins are regulators of autophagy. Cells 2012, 1, 754–773. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Acharya, S.; Sahin, O.; Zhang, Q.; Saito, Y.; Yao, J.; Wang, H.; Li, P.; Zhang, L.; Lowery, F.J.; et al. 14-3-3ζ turns TGF-β’s function from tumor suppressor to metastasis promoter in breast cancer by contextual changes of smad partners from p53 to Gli2. Cancer Cell 2015, 27, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Subramanian, R.R.; Masters, S.C. 14-3-3 proteins: Structure, function, and regulation. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 617–647. [Google Scholar] [CrossRef] [PubMed]
- Masters, S.C.; Fu, H. 14-3-3 proteins mediate an essential anti-apoptotic signal. J. Biol. Chem. 2001, 276, 45193–45200. [Google Scholar] [CrossRef] [PubMed]
- Muda, K.; Bertinetti, D.; Gesellchen, F.; Hermann, J.S.; von Zweydorf, F.; Geerlof, A.; Jacob, A.; Ueffing, M.; Gloeckner, C.J.; Herberg, F.W. Parkinson-related LRRK2 mutation R1441C/G/H impairs PKA phosphorylation of LRRK2 and disrupts its interaction with 14-3-3. Proc. Natl. Acad. Sci. USA 2014, 111, E34–E43. [Google Scholar] [CrossRef] [PubMed]
- van Heusden, G.P.; Steensma, H.Y. Yeast 14-3-3 proteins. Yeast 2006, 23, 159–171. [Google Scholar] [CrossRef] [PubMed]
- BoseDasgupta, S.; Moes, S.; Jenoe, P.; Pieters, J. Cytokine-induced macropinocytosis in macrophages is regulated by 14-3-3ζ through its interaction with serine-phosphorylated coronin 1. FEBS J. 2015, 282, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.L.; Sehnke, P.C.; Ferl, R.J. Isoform-specific subcellular localization among 14-3-3 proteins in arabidopsis seems to be driven by client interactions. Mol. Biol. Cell 2005, 16, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Cognetti, D.; Davis, D.; Sturtevant, J. The Candida albicans 14-3-3 gene, BMH1, is essential for growth. Yeast 2002, 19, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, S.F.; Tsigkari, K.K.; Grammenoudi, S.; Skoulakis, E.M.C. In vivo functional specificity and homeostasis of drosophila 14-3-3 proteins. Genetics 2007, 177, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Lukasiuk, K.; Kontula, L.; Pitkanen, A. cDNA profiling of epileptogenesis in the rat brain. Eur. J. Neurosci. 2003, 17, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.P.; Jimenez-Mateos, E.M.; McKiernan, R.C.; Engel, T.; Tzivion, G.; Henshall, D.C. Transgenic overexpression of 14-3-3ζ protects hippocampus against endoplasmic reticulum stress and status epilepticus in vivo. PLoS ONE 2013, 8, e54491. [Google Scholar] [CrossRef] [PubMed]
- DeLille, J.M.; Sehnke, P.C.; Ferl, R.J. The arabidopsis 14-3-3 family of signaling regulators. Plant Physiol. 2001, 126, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Su, T.T.; Parry, D.H.; Donahoe, B.; Chien, C.T.; O’Farrell, P.H.; Purdy, A. Cell cycle roles for two 14-3-3 proteins during drosophila development. J. Cell Sci. 2001, 114, 3445–3454. [Google Scholar] [PubMed]
- Kong, L.; Lv, Z.; Chen, J.; Nie, Z.; Wang, D.; Shen, H.; Wang, X.; Wu, X.; Zhang, Y. Expression analysis and tissue distribution of two 14-3-3 proteins in silkworm (Bombyx mori). Biochim. Biophys. Acta 2007, 1770, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Noh, M.Y.; Jo, Y.H.; Oh, S.H.; Kim, I.; Lee, Y.S.; Kim, H.C.; Seo, S.J.; Bang, I.S.; Han, Y.S. Peptide-based polyclonal antibody against mosquito 14-3-3ζ recognizes 14-3-3 homolog from dipteran and lepidopteran insects. Entomol. Res. 2009, 39, 129–134. [Google Scholar] [CrossRef]
- Jo, Y.H.; Noh, M.Y.; Kang, S.W.; Kim, D.H.; Oh, S.H.; Lee, Y.S.; Bang, I.S.; Seo, S.J.; Kim, I.; Han, Y.S. Molecular cloning and expression pattern of 14-3-3ζ from the malaria vector, Anopheles sinensis. Entomol. Res. 2009, 39, 123–128. [Google Scholar] [CrossRef]
- Tabunoki, H.; Shimada, T.; Banno, Y.; Sato, R.; Kajiwara, H.; Mita, K.; Satoh, J. Identification of Bombyx mori 14-3-3 orthologs and the interactor Hsp60. Neurosci. Res. 2008, 61, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, R.Q.; Cheng, D.J.; Fan, W.; Zha, X.F.; Cheng, T.C.; Wu, Y.Q.; Wang, J.; Mita, K.; Xiang, Z.H.; et al. SilkDB v2.0: A platform for silkworm (Bombyx mori) genome biology. Nucleic Acids Res. 2010, 38, D453–D456. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.N.; Bian, D.D.; Ge, B.M.; Zhou, C.L.; Tang, B.P. Molecular characterization of a 14-3-3ζ gene from Plodia interpunctella: A potential marker for phylogenetic inference. Biochem. Syst. Ecol. 2015, 60, 171–176. [Google Scholar] [CrossRef]
- Shelby, K.S.; Popham, H.J.R. Analysis of ESTs generated from immune-stimulated hemocytes of larval Heliothis virescens. J. Invertebr. Pathol. 2009, 101, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Ulvila, J.; Vanha-aho, L.M.; Kleino, A.; Vaha-Makila, M.; Vuoksio, M.; Eskelinen, S.; Hultmark, D.; Kocks, C.; Hallman, M.; Parikka, M.; et al. Cofilin regulator 14-3-3ζ is an evolutionarily conserved protein required for phagocytosis and microbial resistance. J. Leukoc. Biol. 2011, 89, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Feng, E.Y.; Chen, H.; Li, Y.; Jiang, W.; Wang, Z.K.; Yin, Y.P. Gene cloning, expression, and function analysis of SpL14-3-3ζ in Spodoptera litura and its response to the entomopathogenic fungus Nomuraea rileyi. Comp. Biochem. Phys. B 2014, 172, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Ma, W.H.; Wang, X.P.; Niu, C.Y.; Lei, C.L. Analysis of pupal head proteome and its alteration in diapausing pupae of Helicoverpa armigera. J. Insect. Physiol. 2010, 56, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Roh, K.B.; Kim, C.H.; Lee, H.; Kwon, H.M.; Park, J.W.; Ryu, J.H.; Kurokawa, K.; Ha, N.C.; Lee, W.J.; Lemaitre, B.; et al. Proteolytic cascade for the activation of the insect toll pathway induced by the fungal cell wall component. J. Biol. Chem. 2009, 284, 19474–19481. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Park, J.W.; Kwon, H.M.; Hwang, H.O.; Jang, I.H.; Masuda, A.; Kurokawa, K.; Nakayama, H.; Lee, W.J.; Dohmae, N.; et al. Diversity of innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects. J. Biol. Chem. 2010, 285, 32937–32945. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, B.B.; Patnaik, H.H.; Seo, G.W.; Jo, Y.H.; Lee, Y.S.; Lee, B.L.; Han, Y.S. Gene structure, cDNA characterization and RNAi-based functional analysis of a myeloid differentiation factor 88 homolog in Tenebrio molitor larvae exposed to Staphylococcus aureus infection. Dev. Comp. Immunol. 2014, 46, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.H.; Kim, Y.J.; Park, K.B.; Seong, J.H.; Kim, S.G.; Park, S.; Noh, M.Y.; Lee, Y.S.; Han, Y.S. TmCactin plays an important role in Gram-negative and -positive bacterial infection by regulating expression of 7 AMP genes in Tenebrio molitor. Sci. Rep. 2017, 7, 46459. [Google Scholar] [CrossRef] [PubMed]
- Tindwa, H.; Patnaik, B.B.; Kim, D.H.; Mun, S.; Jo, Y.H.; Lee, B.L.; Lee, Y.S.; Kim, N.J.; Han, Y.S. Cloning, characterization and effect of TmPGRP-LE gene silencing on survival of Tenebrio molitor against Listeria monocytogenes infection. Int. J. Mol. Sci. 2013, 14, 22462–22482. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, B.B.; Kang, S.M.; Seo, G.W.; Lee, H.J.; Patnaik, H.H.; Jo, Y.H.; Tindwa, H.; Lee, Y.S.; Lee, B.L.; Kim, N.J.; et al. Molecular cloning, sequence characterization and expression analysis of a CD63 homologue from the coleopteran beetle, Tenebrio molitor. Int. J. Mol. Sci. 2013, 14, 20744–20767. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Patnaik, B.B.; Seo, G.W.; Kang, S.M.; Lee, Y.S.; Lee, B.L.; Han, Y.S. Identification and expression analysis of a novel R-type lectin from the coleopteran beetle, Tenebrio molitor. J. Invertebr. Pathol. 2013, 114, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.Y.; Patnaik, B.B.; Tindwa, H.; Seo, G.W.; Kim, D.H.; Patnaik, H.H.; Jo, Y.H.; Lee, Y.S.; Lee, B.L.; Kim, N.J.; et al. Genomic organization, sequence characterization and expression analysis of Tenebrio molitor apolipophorin-III in response to an intracellular pathogen, Listeria monocytogenes. Gene 2014, 534, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Seo, G.W.; Jo, Y.H.; Seong, J.H.; Park, K.B.; Patnaik, B.B.; Tindwa, H.; Kim, S.A.; Lee, Y.S.; Kim, Y.J.; Han, Y.S. The silencing of a 14-3-3varepsilon homolog in Tenebrio molitor leads to increased antimicrobial activity in hemocyte and reduces larval survivability. Genes 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Tindwa, H.; Jo, Y.H.; Patnaik, B.B.; Noh, M.Y.; Kim, D.H.; Kim, I.; Han, Y.S.; Lee, Y.S.; Lee, B.L.; Kim, N.J. Depletion of autophagy-related genes ATG3 and ATG5 in Tenebrio molitor leads to decreased survivability against an intracellular pathogen, Listeria monocytogenes. Arch. Insect Biochem. 2015, 88, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Tindwa, H.; Jo, Y.H.; Patnaik, B.B.; Lee, Y.S.; Kang, S.S.; Han, Y.S. Molecular cloning and characterization of autophagy-related gene TmATG8 in Listeria-invaded hemocytes of Tenebrio molitor. Dev. Comp. Immunol. 2015, 51, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.M.; Booker, M.; Silver, S.J.; Friedman, A.; Hong, P.; Perrimon, N.; Mathey-Prevot, B. Evidence of off-target effects associated with long dsRNAs in Drosophila melanogaster cell-based assays. Nat. Methods 2006, 3, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Creanga, A.; Lum, L.; Beachy, P.A. Prevalence of off-target effects in Drosophila RNA interference screens. Nature 2006, 443, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.K.; Morrison, D.K. 14-3-3 proteins: Diverse functions in cell proliferation and cancer progression. Semin. Cell Dev. Biol. 2011, 22, 681–687. [Google Scholar] [CrossRef] [PubMed]
- de Boer, A.H.; van Kleeff, P.J.; Gao, J. Plant 14-3-3 proteins as spiders in a web of phosphorylation. Protoplasma 2013, 250, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Matta, A.; Siu, K.W.M.; Ralhan, R. 14-3-3 zeta as novel molecular target for cancer therapy. Expert Opin. Ther. Targets. 2012, 16, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.; Saraiva, M.J. Transthyretin regulates lysosomal degradation of hippocampal 14-3-3ζ. FASEB J. 2013, 27, 557. [Google Scholar]
- Zhu, B.J.; Wang, D.J.; Peng, T.; Wang, L.; Wei, G.Q.; Liu, C.L. Characterization and function of a gene Pc 14-3-3 isoform from red crayfish, Procambarus clarkii. Pak. J. Zool. 2014, 46, 107–113. [Google Scholar]
- Morrison, D.K. The 14-3-3 proteins: Integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009, 19, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wu, S.; Zhen, H.; Deng, H.; Song, Q.; Ma, K.; Cao, Z.; Pang, Q.; Zhao, B. 14-3-3α and 14-3-3ζ contribute to immune responses in planarian Dugesia japonica. Gene 2017, 615, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Obsilova, V.; Kopecka, M.; Kosek, D.; Kacirova, M.; Kylarova, S.; Rezabkova, L.; Obsil, T. Mechanisms of the 14-3-3 protein function: Regulation of protein function through conformational modulation. Physiol. Res. 2014, 63, S155–S164. [Google Scholar] [PubMed]
- Alexander, R.D.; Morris, P.C. A proteomic analysis of 14-3-3 binding proteins from developing barley grains. Proteomics 2006, 6, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Roth, D.; Morgan, A.; Burgoyne, R.D. Identification of a key domain in annexin and 14-3-3 proteins that stimulate calcium-dependent exocytosis in permeabilized adrenal chromaffin cells. FEBS Lett. 1993, 320, 207–210. [Google Scholar] [CrossRef]
- Trujillo-Ocampo, A.; Cazares-Raga, F.E.; Celestino-Montes, A.; Cortes-Martinez, L.; Rodriguez, M.H.; Hernandez-Hernandez, F.C. Identification and expression analysis of two 14-3-3 proteins in the mosquito Aedes aegypti, an important arboviruses vector. Arch. Insect Biochem. Physiol. 2016, 93, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, J.G.; Ying, S.H.; Wang, J.J.; Sun, W.L.; Tian, C.G.; Feng, M.G. Unveiling equal importance of two 14-3-3 proteins for morphogenesis, conidiation, stress tolerance and virulence of an insect pathogen. Environ. Microbiol. 2015, 17, 1444–1462. [Google Scholar] [CrossRef] [PubMed]
- Funami, K.; Matsumoto, M.; Obuse, C.; Seya, T. 14-3-3-zeta participates in TLR3-mediated TICAM-1 signal-platform formation. Mol. Immunol. 2016, 73, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Han, Y.; Jiao, H.; Jie, Y. 14-3-3zeta regulates immune response through Stat3 signaling in oral squamous cell carcinoma. Mol. Cells 2015, 38, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Bonner, H.P.; Ward, M.W.; Murphy, B.M.; Prehn, J.H.; Henshall, D.C. Depletion of 14-3-3ζ zeta elicits endoplasmic reticulum stress and cell death, and increases vulnerability to kainate-induced injury in mouse hippocampal cultures. J. Neurochem. 2008, 106, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Heverin, M.; Brennan, G.P.; Koehler, C.J.; Treumann, A.; Henshall, D.C. Proteomic analysis of 14-3-3ζ binding proteins in the mouse hippocampus. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 74–83. [Google Scholar] [PubMed]
- Kafle, A.; Puchadapirom, P.; Plumworasawat, S.; Dontumprai, R.; Chan-On, W.; Buates, S.; Laha, T.; Sripa, B.; Suttiprapa, S. Identification and characterization of protein 14-3-3 in carcinogenic liver fluke Opisthorchis viverrini. Parasitol. Int. 2017, 66, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, B.B.; Patnaik, H.H.; Park, K.B.; Jo, Y.H.; Lee, Y.S.; Han, Y.S. Silencing of apolipophorin-III causes abnormal adult morphological phenotype and susceptibility to Listeria monocytogenes infection in Tenebrio molitor. Entomol. Res. 2015, 45, 116–121. [Google Scholar] [CrossRef]
- Pozuelo-Rubio, M. Regulation of autophagic activity by 14-3-3ζ proteins associated with class III phosphatidylinositol-3-kinase. Cell Death Differ. 2011, 18, 479–492. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequences |
|---|---|
| Tm14-3-3ζ 5′-RACE GSP1 | 5′-GATGAGAGTGCTGTCCTT-3′ |
| Tm14-3-3ζ 5′-RACE nGSP2 | 5′-TAATCTGATGGGATGTGTGGGCGT-3′ |
| Tm14-3-3ζ 3′-RACE GSP1 | 5′-ATTTGATCCCCAAAGCAAGC-3′ |
| Tm14-3-3ζ 3′-RACE nGSP2 | 5′-CCCACACATCCCATCAGATT-3′ |
| Tm14-3-3ζ Ex(BamHI) Fw | 5′-GGGGGATCCATGTCCGTCACTGACAAAGATGAG-3′ |
| Tm14-3-3ζ Ex(HindIII) Rv | 5′-GGGAAGCTTCAATTATCACCAGTCTCTTGAGGC-3′ |
| Tm14-3-3ζ qPCR Fw | 5′-TTTGGCGGAAGTAGCCACAGGAGA-3′ |
| Tm14-3-3ζ qPCR Rv | 5′-TAATCTGATGGGATGTGTGGGCGT-3′ |
| TmL27a qPCR Fw | 5′-TCATCCTGAAGGCAAAGCTCCAGT-3′ |
| TmL27a qPCR Rv | 5′-AGGTTGGTTAGGCAGGCACCTTTA-3′ |
| BamHI and HindIII recognition sites have been underlined. | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seong, J.H.; Jo, Y.H.; Seo, G.W.; Park, S.; Park, K.B.; Cho, J.H.; Ko, H.J.; Kim, C.E.; Patnaik, B.B.; Jun, S.A.; et al. Molecular Cloning and Effects of Tm14-3-3ζ-Silencing on Larval Survivability Against E. coli and C. albicans in Tenebrio molitor. Genes 2018, 9, 330. https://doi.org/10.3390/genes9070330
Seong JH, Jo YH, Seo GW, Park S, Park KB, Cho JH, Ko HJ, Kim CE, Patnaik BB, Jun SA, et al. Molecular Cloning and Effects of Tm14-3-3ζ-Silencing on Larval Survivability Against E. coli and C. albicans in Tenebrio molitor. Genes. 2018; 9(7):330. https://doi.org/10.3390/genes9070330
Chicago/Turabian StyleSeong, Jeong Hwan, Yong Hun Jo, Gi Won Seo, Soyi Park, Ki Beom Park, Jun Ho Cho, Hye Jin Ko, Chang Eun Kim, Bharat Bhusan Patnaik, Sung Ah Jun, and et al. 2018. "Molecular Cloning and Effects of Tm14-3-3ζ-Silencing on Larval Survivability Against E. coli and C. albicans in Tenebrio molitor" Genes 9, no. 7: 330. https://doi.org/10.3390/genes9070330
APA StyleSeong, J. H., Jo, Y. H., Seo, G. W., Park, S., Park, K. B., Cho, J. H., Ko, H. J., Kim, C. E., Patnaik, B. B., Jun, S. A., Choi, Y. S., Kim, Y. W., Bang, I. S., Lee, Y. S., & Han, Y. S. (2018). Molecular Cloning and Effects of Tm14-3-3ζ-Silencing on Larval Survivability Against E. coli and C. albicans in Tenebrio molitor. Genes, 9(7), 330. https://doi.org/10.3390/genes9070330