Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Hepatitis B Virus Serological Screening
2.3. Hepatitis B Virus DNA Levels
2.4. DNA Extraction for Genotyping
2.5. HBV Genotyping
2.6. Data Analysis
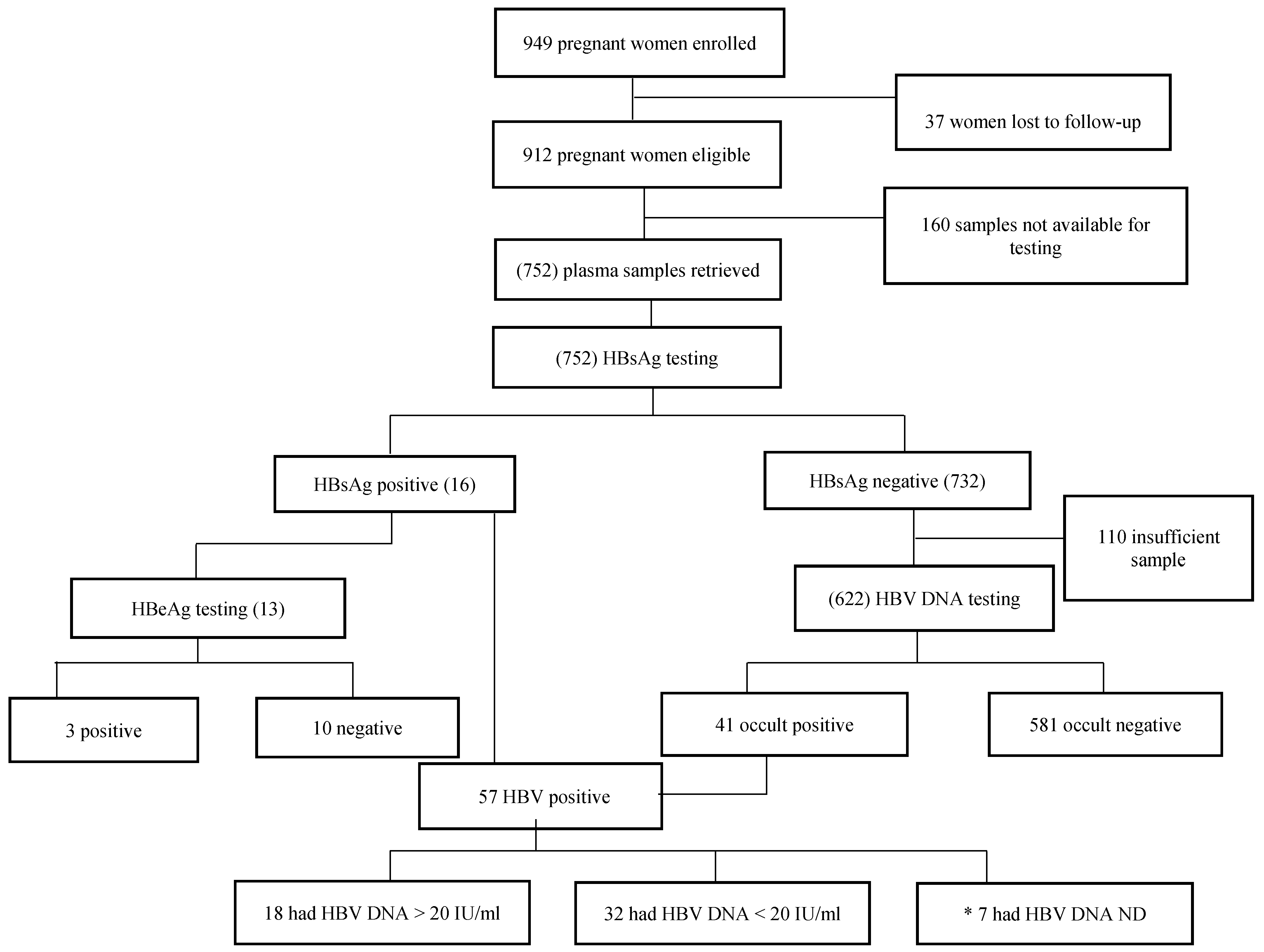
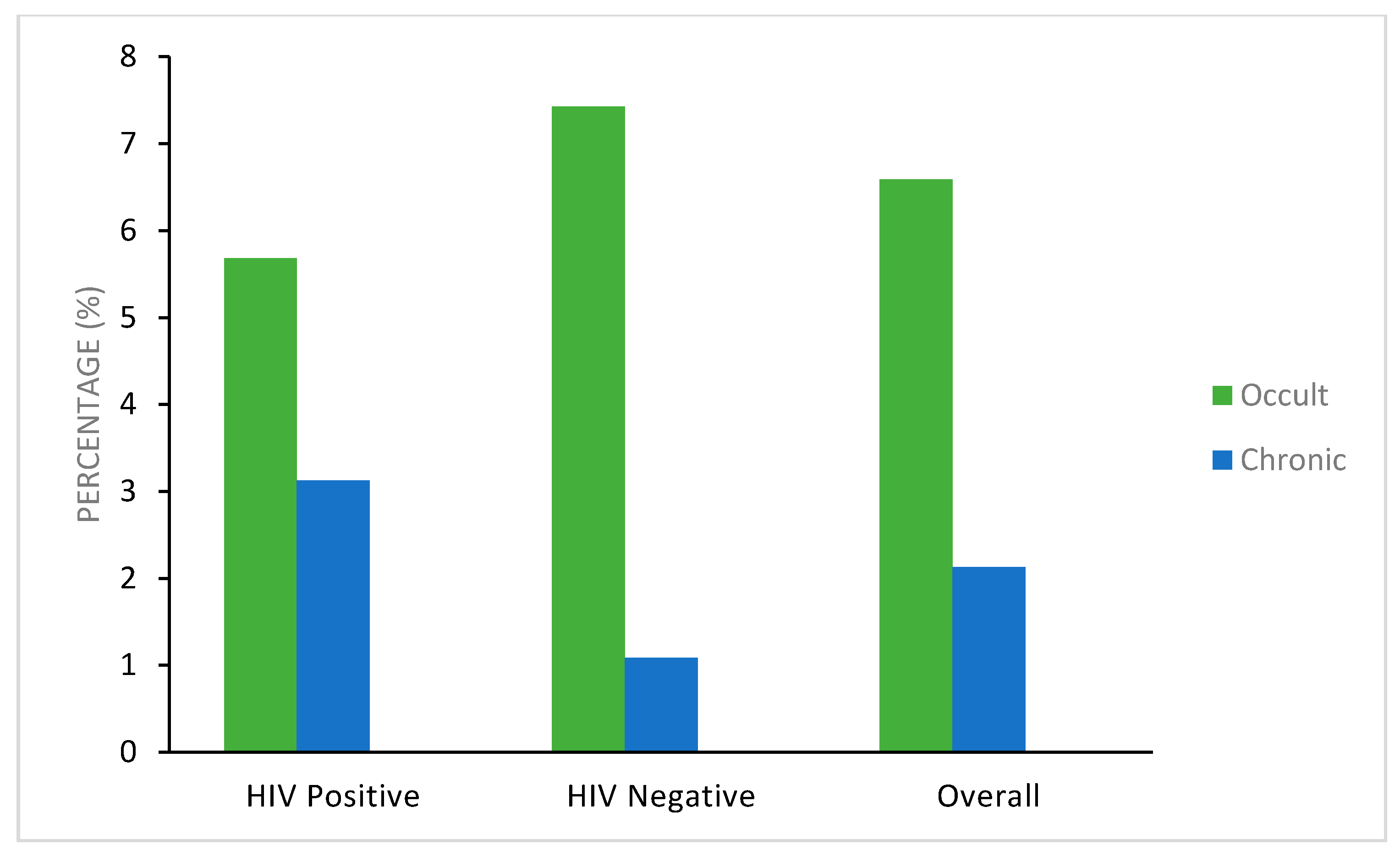
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Stanaway, J.D.; Flaxman, A.D.; Naghavi, M.; Fitzmaurice, C.; Vos, T.; Abubakar, I.; Abu-Raddad, L.J.; Assadi, R.; Bhala, N.; Cowie, B.; et al. The global burden of viral hepatitis from 1990 to 2013: Findings from the Global Burden of Disease Study 2013. Lancet 2016, 388, 1081–1088. [Google Scholar] [CrossRef]
- Bengsch, B.; Chang, K.M. Evolution in our understanding of hepatitis B virus virology and immunology. Clin. Liver Dis. 2016, 20, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, A.; Horn, J.; Mikolajczyk, R.T.; Krause, G.; Ott, J.J. Estimations of worldwide prevalence of chronic hepatitis B virus infection: A systematic review of data published between 1965 and 2013. Lancet 2015, 386, 1546–1555. [Google Scholar] [CrossRef]
- Anderson, M.; Gaseitsiwe, S.; Moyo, S.; Wessels, M.J.C.; Mohammed, T.; Sebunya, T.K.; Powell, E.A.; Makhema, J.; Blackard, J.T.; Marlink, R.; et al. Molecular characterisation of hepatitis B virus in HIV-1 subtype C infected patients in Botswana. BMC Infect. Dis. 2015, 15, 335. [Google Scholar] [CrossRef] [PubMed]
- Glebe, D.; Konig, A. Molecular virology of hepatitis B virus and targets for antiviral intervention. Intervirology 2014, 57, 134–140. [Google Scholar] [CrossRef] [PubMed]
- McGovern, B.H. The epidemiology, natural history and prevention of hepatitis B: Implications of HIV coinfection. Antivir. Ther. 2007, 12 (Suppl 3), H3–H13. [Google Scholar] [PubMed]
- Bodsworth, N.; Donovan, B.; Nightingale, B.N. The effect of concurrent human immunodeficiency virus infection on chronic hepatitis B: A study of 150 homosexual men. J. Infect. Dis. 1989, 160, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Colin, J.F.; Cazals-Hatem, D.; Loriot, M.A.; Martinot-Peignoux, M.; Pham, B.N.; Auperin, A.; Degott, C.; Benhamou, J.P.; Erlinger, S.; Valla, D.; et al. Influence of human immunodeficiency virus infection on chronic hepatitis B in homosexual men. Hepatology 1999, 29, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, S.M.; Kocharian, A.; Hopfensperger, D.; Davis, J.P. Prevention of perinatal transmission of hepatitis B virus: Assessment among Wisconsin Maternity Hospitals. WMJ Off. Publ. State Med. Soc. Wis. 2016, 115, 74–79. [Google Scholar]
- Pande, C.; Sarin, S.K.; Patra, S.; Kumar, A.; Mishra, S.; Srivastava, S.; Bhutia, K.; Gupta, E.; Mukhopadhyay, C.K.; Dutta, A.K.; et al. Hepatitis B vaccination with or without hepatitis B immunoglobulin at birth to babies born of HBsAg-positive mothers prevents overt HBV transmission but may not prevent occult HBV infection in babies: A randomized controlled trial. J. Viral Hepat. 2013, 20, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hou, X.; Cao, G. Is mother-to-infant transmission the most important factor for persistent HBV infection? Emerg. Microb. Infect. 2015, 4, e30. [Google Scholar] [CrossRef] [PubMed]
- WHO. Hepatits B; World Health Organization: Geneva, Switzerland, 2017; Available online: http://www.who.int/mediacentre/factsheets/fs204/en/ (accessed on 27 January 2018).
- Statistics Botswana. Botswana Impact AIDS Survey IV; Statistics Botswana: Gaborone, Botswana, 2013. [Google Scholar]
- Patel, P.; Davis, S.; Tolle, M.; Mabikwa, V.; Anabwani, G. Prevalence of hepatitis B and hepatitis C coinfections in an adult HIV centre population in Gaborone, Botswana. Am. J. Trop. Med. Hyg. 2011, 85, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Wester, C.W.; Bussmann, H.; Moyo, S.; Avalos, A.; Gaolathe, T.; Ndwapi, N.; Essex, M.; MacGregor, R.R.; Marlink, R.G. Serological evidence of HIV-associated infection among HIV-1-infected adults in Botswana. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2006, 43, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Matthews, P.C.; Beloukas, A.; Malik, A.; Carlson, J.M.; Jooste, P.; Ogwu, A.; Shapiro, R.; Riddell, L.; Chen, F.; Luzzi, G.; et al. Prevalence and characteristics of hepatitis B virus (HBV) coinfection among HIV-positive women in south Africa and Botswana. PLoS ONE 2015, 10, e0134037. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Williams, P.L.; Mayondi, G.K.; Leidner, J.; Holding, P.; Tepper, V.; Nichols, S.; Magetse, J.; Sakoi, M.; Moabi, K.; et al. Neurodevelopment of HIV-exposed and HIV-unexposed uninfected children at 24 months. Pediatrics 2017. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ding, Y.; Juan, F.; Dou, X.G. Genotyping the hepatitis B virus with a fragment of the HBV DNA polymerase gene in Shenyang, China. Virol. J. 2011, 8, 315. [Google Scholar] [CrossRef] [PubMed]
- Sequencher® DNA Sequence Analysis Software, version 5.4.6; Gene Codes Corporation: Ann Arbor, MI, USA; Available online: http://www.genecodes.com (accessed on 11 May 2018).
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, syy032. [Google Scholar] [CrossRef] [PubMed]
- Molecular Evolution, Phylogenetics and Epidemiology. FigTree, Version 1.3.1. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 May 2018).
- Martin, C.M.; Welge, J.A.; Blackard, J.T. Hepatitis B virus (HBV) X gene diversity and evidence of recombination in HBV/HIV co-infected persons. J. Med. Virol. 2011, 83, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.A.; Gededzha, M.P.; Rentz, M.; Rakgole, N.J.; Selabe, S.G.; Seleise, T.A.; Mphahlele, M.J.; Blackard, J.T. Mutations associated with occult hepatitis B in HIV-positive South Africans. J. Med. Virol. 2015, 87, 388–400. [Google Scholar] [CrossRef] [PubMed]
- StataCorp. Stata Statistical Software: Release 14; StataCorp LP: College Station, TX, USA, 2015. [Google Scholar]
- Seremba, E.; Ssempijja, V.; Kalibbala, S.; Gray, R.H.; Wawer, M.J.; Nalugoda, F.; Casper, C.; Phipps, W.; Ocama, P.; Serwadda, D.; et al. Hepatitis B incidence and prevention with antiretroviral therapy among HIV-positive individuals in Uganda. AIDS 2017, 31, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.J.; Mashabela, F.; Cohn, S.; Hoffmann, J.D.; Lala, S.; Martinson, N.A.; Chaisson, R.E. Maternal hepatitis B and infant infection among pregnant women living with HIV in South Africa. J. Int. AIDS Soc. 2014, 17, 18871. [Google Scholar] [CrossRef] [PubMed]
- Umare, A.; Seyoum, B.; Gobena, T.; Haile Mariyam, T. Hepatitis B virus infections and associated factors among pregnant women attending antenatal care clinic at Deder Hospital, Eastern Ethiopia. PLoS ONE 2016, 11, e0166936. [Google Scholar] [CrossRef] [PubMed]
- Bayo, P.; Ochola, E.; Oleo, C.; Mwaka, A.D. High prevalence of hepatitis B virus infection among pregnant women attending antenatal care: A cross-sectional study in two hospitals in northern Uganda. BMJ Open 2014, 4, e005889. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Anderson, M.; Gyurova, I.; Ambroggio, L.; Moyo, S.; Sebunya, T.; Makhema, J.; Marlink, R.; Essex, M.; Musonda, R.; et al. High rates of occult hepatitis B virus infection in HIV-positive individuals initiating antiretroviral therapy in Botswana. Open Forum Infect. Dis. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Belopolskaya, M.; Avrutin, V.; Firsov, S.; Yakovlev, A. HBsAg level and hepatitis B viral load correlation with focus on pregnancy. Ann. Gastroenterol. Q. Publ. Hell. Soc. Gastroenterol. 2015, 28, 379–384. [Google Scholar]
- Su, H.; Zhang, Y.; Xu, D.; Wang, B.; Zhang, L.; Li, D.; Xiao, D.; Li, F.; Zhang, J.; Yan, Y. Occult hepatitis B virus infection in anti-HBs-positive infants born to HBsAg-positive mothers in China. PLoS ONE 2013, 8, e70768. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liang, X.F.; Wang, F.Z.; Yan, L.; Li, R.C.; Li, Y.P.; Zhu, F.C.; Zhai, X.J.; Li, J.; Zhuang, H. Hepatitis B vaccine alone may be enough for preventing hepatitis B virus transmission in neonates of HBsAg (+)/HBeAg (-) mothers. Vaccine 2017, 35, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.Q.; Duan, Z.; Dai, E.; Zhang, S.; Han, G.; Wang, Y.; Zhang, H.; Zou, H.; Zhu, B.; Zhao, W.; et al. Tenofovir to prevent hepatitis B transmission in mothers with high viral load. N. Engl. J. Med. 2016, 374, 2324–2334. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kourtis, A.P.; Ellington, S.; Legardy-Williams, J.; Bulterys, M. Safety of tenofovir during pregnancy for the mother and fetus: A systematic review. Clin. Infect. Dis. 2013, 57, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Miailhes, P.; Trabaud, M.A.; Pradat, P.; Lebouche, B.; Chevallier, M.; Chevallier, P.; Zoulim, F.; Trepo, C. Impact of highly active antiretroviral therapy (HAART) on the natural history of hepatitis B virus (HBV) and HIV coinfection: Relationship between prolonged efficacy of HAART and HBV surface and early antigen seroconversion. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, 624–632. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ayoub, W.S.; Cohen, E. Hepatitis B management in the pregnant patient: An update. J. Clin. Transl. Hepatol. 2016, 4, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health. Handbook of the Botswana 2016 Integrated HIV Clinical Care Guidelines; Ministry of Health: Gaborone, Botswana, 2016.
- Cohen Stuart, J.W.T.; Velema, M.; Schuurman, R.; Boucher, C.A.B.; Hoepelman, A.I.M. Occult hepatitis B in persons infected with HIV is associated with low CD4 counts and resolves during antiretroviral therapy. J. Med. Virol. 2009, 81, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Khamduang, W.; Ngo-Giang-Huong, N.; Gaudy-Graffin, C.; Jourdain, G.; Suwankornsakul, W.; Jarupanich, T.; Chalermpolprapa, V.; Nanta, S.; Puarattana-aroonkorn, N.; Tonmat, S.; et al. Prevalence, risk factors, and impact of isolated antibody to hepatitis B core antigen and occult hepatitis b virus infection in HIV-1–infected pregnant women. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Lewden, C.; Chene, G.; Morlat, P.; Raffi, F.; Dupon, M.; Dellamonica, P.; Pellegrin, J.L.; Katlama, C.; Dabis, F.; Leport, C. HIV-infected adults with a CD4 cell count greater than 500 cells/mm3 on long-term combination antiretroviral therapy reach same mortality rates as the general population. J. Acquir. Immune Defic. Syndr. 2007, 46, 72–77. [Google Scholar] [PubMed]
- Institute of Medicine Committee on Social Security. HIV and Disability: Updating the Social Security Listings; National Academies Press (US): Washington, DC, USA, 2010. [Google Scholar]
- Lundgren, J.D.; Babiker, A.G.; Gordin, F.; Emery, S.; Grund, B.; Sharma, S.; Avihingsanon, A.; Cooper, D.A.; Fatkenheuer, G.; Llibre, J.M.; et al. Initiation of antiretroviral therapy in early asymptomatic HIV infection. N. Engl. J. Med. 2015, 373, 795–807. [Google Scholar] [PubMed]
- Makondo, E.; Bell, T.G.; Kramvis, A. Genotyping and molecular characterization of hepatitis B virus from human immunodeficiency virus-infected individuals in southern africa. PLoS ONE 2012, 7, e46345. [Google Scholar] [CrossRef] [PubMed]
- Torii, Y.; Kimura, H.; Hayashi, K.; Suzuki, M.; Kawada, J.-I.; Kojima, S.; Katano, Y.; Goto, H.; Ito, Y. Causes of vertical transmission of hepatitis B virus under the at-risk prevention strategy in Japan. Microbiol. Immunol. 2013, 57, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Poortahmasebi, V.; Poorebrahim, M.; Ghaziasadi, A.; Abazari, M.F.; Mozhgani, S.; Nouri Aleagha, M.; Shahbazi, F.; Alavian, S.M. Analysis of antigenic and conformational changes in hepatitis B surface antigen (HBsAg) identified in iranian patients with chronic hepatitis B. Hepat. Mon. 2017, 17, e57882. [Google Scholar] [CrossRef]
- Yan, B.; Lv, J.; Feng, Y.; Liu, J.; Ji, F.; Xu, A.; Zhang, L. Temporal trend of hepatitis B surface mutations in the post-immunization period: 9 years of surveillance (2005–2013) in eastern China. Sci. Rep. 2017, 7, 6669. [Google Scholar] [CrossRef] [PubMed]
- Asahina, Y.; Enomoto, N.; Ogura, Y.; Kurosaki, M.; Sakuma, I.; Izumi, N.; Marumo, F.; Sato, C. Sequential changes in full-length genomes of hepatitis B virus accompanying acute exacerbation of chronic hepatitis B. J. Hepatol. 1996, 25, 787–794. [Google Scholar] [CrossRef]
- Wilson, P.; Parr, J.B.; Jhaveri, R.; Meshnick, S.R. Call to action: Prevention of mother-to-child transmission of hepatitis B in Africa. J. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kiire, C.F.; The African Regional Study Group. Hepatitis B infection in sub-Saharan Africa. Vaccine 1990, 8, S107–S112; discussion S134–S108. [Google Scholar] [CrossRef]
- Matthews, P.C.; Geretti, A.M.; Goulder, P.J.; Klenerman, P. Epidemiology and impact of HIV coinfection with hepatitis B and hepatitis C viruses in Sub-Saharan Africa. J. Clin. Virol. 2014, 61, 20–33. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| All (N = 752) | Chronic HBV (N = 16) | Occult HBV (N = 41) | p-Value | Chronic or Occult HBV (N = 57) | HBV-Negative (N = 695) | p-Value | |
|---|---|---|---|---|---|---|---|
| Media maternal age in years (n = 581) (Q1, Q3) | 27 (24, 32) | 29 (25, 33) | 28 (23, 32) | 0.58 | 27 (23, 31) | 27 (23, 32) | 0.24 |
| HIV-positive women | 384 * and 299 + | 12 (3.1%) | 17 (5.7%) | 0.057 a | 29 (7.6%) | 355 (92.4%) | 0.54 |
| HIV-negative women | 368 * and 323 + | 4 (1.1%) | 24 (7.4%) | 0.38 b | 28 (7.6%) | 340 (92.3%) | |
| HIV viral load (copies/mL) (Q1, Q3) | 1549.5 (400, 13,808) | 508.5 (193, 3819) | 1568 (395, 9635) | 0.70 | 676 (400, 7762) | 1585 (400, 14,125) | 0.16 |
| CD4+ T-cell count (cells/μL) (Q1, Q3) | 422 (313–567) | 460 (340, 511) | 604 (322, 616) | 0.14 | 481 (289, 628) | 418 (313, 563) | 0.25 |
| Platelet count (Q1, Q3) | 267 (216–311) | 296 (211, 365) | 272 (228, 371) | 0.18 | 298 (233, 371) | 265 (211, 309) | 0.52 |
| Hemoglobin (g/dL) (Q1, Q3) | 11 (10–12) | 12 (11, 12) | 11 (9.0, 12) | 0.19 | 12 (10, 13) | 10.9 (9, 12) | 1.00 |
| ALT (U/L) (Q1, Q3) | 13 (11, 19) | 12 (10, 13) | 14 (12, 19) | 0.74 | 13 (10, 14) | 13 (11, 17) | 0.65 |
| AST (U/L) (Q1, Q3) | 23.4 (19, 30) | 24 (18, 30) | 22 (18, 30) | 0.30 | 30 (21, 35) | 23 (19, 30) | 0.35 |
| Total bilirubin (μmol/L) (Q1, Q3) | 6 (4, 8) | 4 (2.9, 5.7) | 4.7 (4, 12) | 0.10 | 4 (3, 6) | 6 (4, 8) | 1.00 |
| Participant Code | HBV DNA at Enrollment (IU/L) | HBV DNA at Delivery (IU/L) | HBeAg at Delivery | HIV Status | ART Regimen | Genotype |
|---|---|---|---|---|---|---|
| TAM 1 | NT | ND | NEG | POS | AZT | D |
| TAM 2 | NT | ND | NEG | POS | AZT | A |
| TAM 5 | 8710 | ND | NEG | POS | AZT | A |
| TAM 6 | <20 | ND | NEG | POS | CBV + NVP | A |
| TAM 7 | <20 | 2250 | NEG | POS | AZT | A |
| TAM 8 | >1.70 × 108 | >1.70 × 108 | POS | NEG | NA | D |
| TAM 9 | >1.70 × 108 | >1.70 × 108 | POS | NEG | NA | A |
| TAM 10 | >1.70 × 108 | >1.70 × 108 | POS | NEG | NA | D |
| TAM 11 | 170 | 396 | NT | NEG | NA | E |
| TAM 12 | NT | 12,900 | NT | POS | AZT | D |
| TAM 13 | NT | ND | NT | POS | NVP ± TRU | D |
| TAM 14 | 25 | ND | NEG | POS | CBV + NVP | NA |
| TAM 15 | 24.8 | <20 | NEG | POS | ATR | NA |
| TAM 16 | 237 | ND | NEG | POS | CBV + NVP | NA |
| TAM 17 | 82 | 166 | NEG | POS | AZT | NA |
| TAM 18 | ND | 2300 | NT | POS | AZT | NA |
| CD4+ T-Cell Count (cells/mL) | Chronic HBV infection (n = 382) a | Occult HBV infection (n = 297) b | AZT (n = 267) | HAART (n = 267) |
|---|---|---|---|---|
| > 500 | 3/129 (2.33%) | 10/96 (10.4%) | 78/96 (81.3%) | 15/96 (15.6%) |
| ≤ 500 | 9/253 (3.56%) | 7/201 (3.48%) | 124/171 (72.5%) | 43/171 (25.1%) |
| p-value | 0.511 | 0.01 | 0.110 | 0.07 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbangiwa, T.; Kasvosve, I.; Anderson, M.; Thami, P.K.; Choga, W.T.; Needleman, A.; Phinius, B.B.; Moyo, S.; Leteane, M.; Leidner, J.; et al. Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes 2018, 9, 259. https://doi.org/10.3390/genes9050259
Mbangiwa T, Kasvosve I, Anderson M, Thami PK, Choga WT, Needleman A, Phinius BB, Moyo S, Leteane M, Leidner J, et al. Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes. 2018; 9(5):259. https://doi.org/10.3390/genes9050259
Chicago/Turabian StyleMbangiwa, Tshepiso, Ishmael Kasvosve, Motswedi Anderson, Prisca K. Thami, Wonderful T. Choga, Austen Needleman, Bonolo B. Phinius, Sikhulile Moyo, Melvin Leteane, Jean Leidner, and et al. 2018. "Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana" Genes 9, no. 5: 259. https://doi.org/10.3390/genes9050259
APA StyleMbangiwa, T., Kasvosve, I., Anderson, M., Thami, P. K., Choga, W. T., Needleman, A., Phinius, B. B., Moyo, S., Leteane, M., Leidner, J., Blackard, J. T., Mayondi, G., Kammerer, B., Musonda, R. M., Essex, M., Lockman, S., & Gaseitsiwe, S. (2018). Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes, 9(5), 259. https://doi.org/10.3390/genes9050259