Journey into Bone Models: A Review


Abstract
1. Introduction
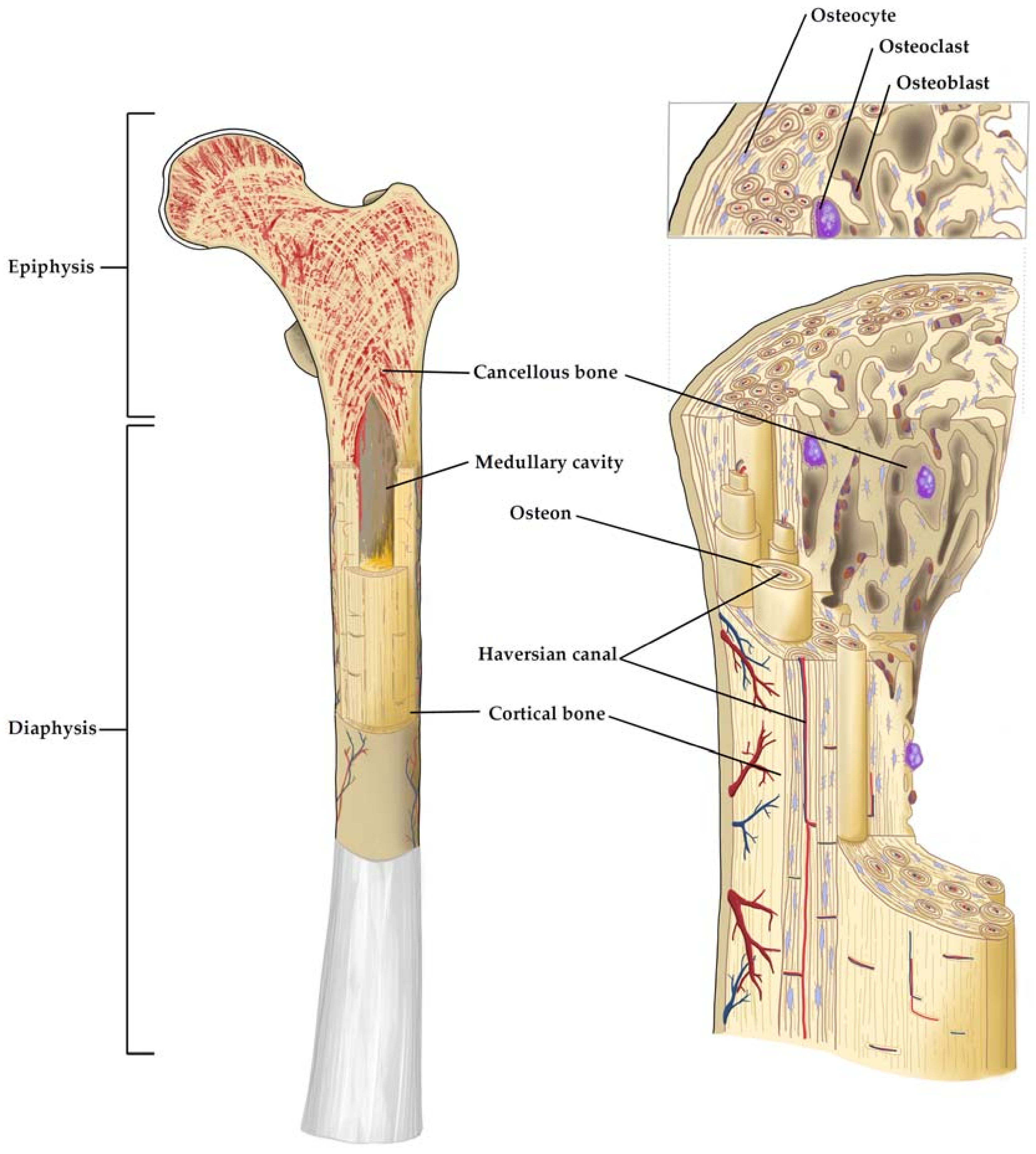
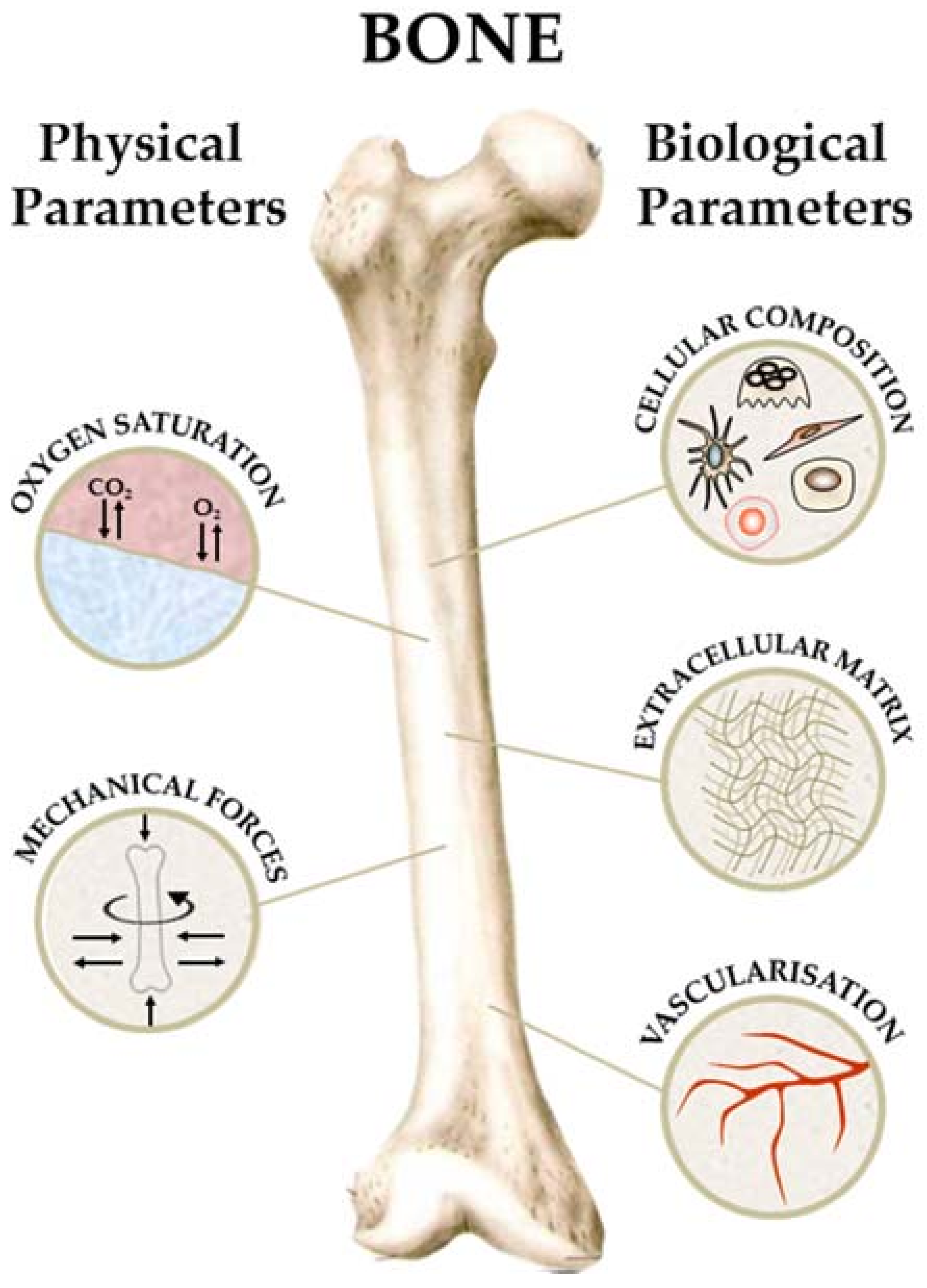
2. Bone Physiology
3. Cell Types and Sources for Bone Tissue Engineering
3.1. Mesenchymal Stromal Cells—The Work Horse in Bone Research
3.2 Bone-Forming and -Resorbing Cells, Cell Lines and Stem Cells
4. Approaches for Creating a Bone-Like Organoid
4.1. Scaffold-Free Self-Organisation in Spheroids
4.2. Scaffold-Based Approaches for Bone Tissue Engineering
4.2.1. Hydroxyapatite and its Composites as Scaffold Materials in Human Bone Models
4.2.2. Tricalcium Phosphate and Its Composites as Scaffolds in Human Bone Models
4.3. Hydrogel-Based Bone Models
4.3.1. Naturally Derived Polymer Hydrogels
4.3.2. Synthetic Polymer Hydrogels
4.3.3. Scaffold–Hydrogels Hybrids
4.4. Bead-Based Tissue Engineering
4.5. 3D Printing
5. Beyond the Dish Culturing Bone Models under Controllable Conditions
5.1. Bioreactors for Culturing Bone
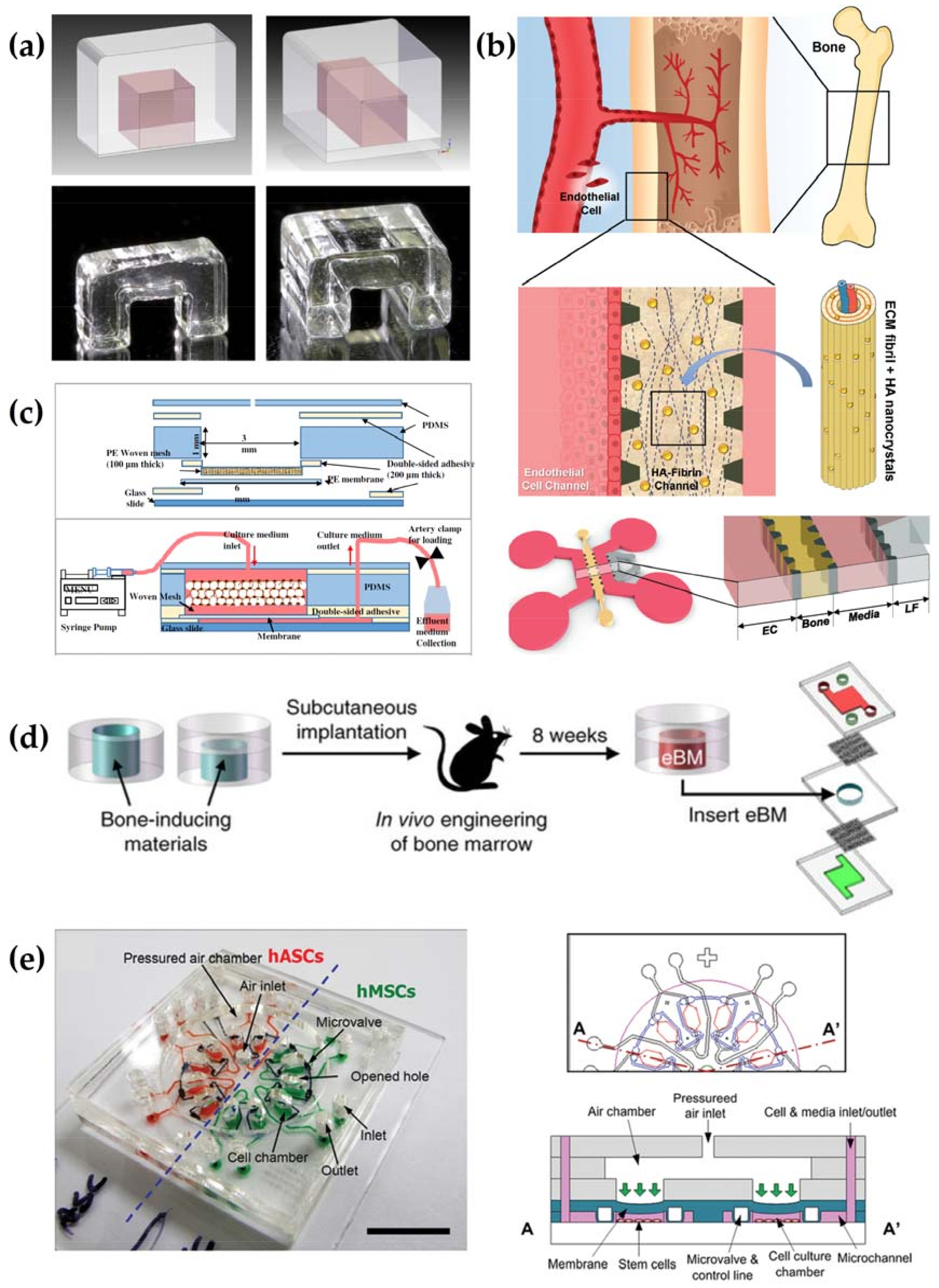
5.2. Microfluidics and the Next Generation Bone Bioreactor
6. Concluding Remarks
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kling, J.M.; Clarke, B.L.; Sandhu, N.P. Osteoporosis prevention, screening and treatment: A review. J. Womens Health 2014, 23, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Learmonth, I.D.; Young, C.; Rorabeck, C. The operation of the century: Total hip replacement. Lancet 2007, 370, 1508–1519. [Google Scholar] [CrossRef]
- Knight, A. Systematic reviews of animal experiments demonstrate poor human clinical and toxicological utility. Altern. Lab. Anim. 2007, 35, 641–659. [Google Scholar] [CrossRef] [PubMed]
- Weidner, C.; Steinfath, M.; Opitz, E.; Oelgeschlager, M.; Schonfelder, G. Defining the optimal animal model for translational research using gene set enrichment analysis. EMBO Mol. Med. 2016, 8, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Marx, U.; Walles, H.; Hoffmann, S.; Lindner, G.; Horland, R.; Sonntag, F.; Klotzbach, U.; Sakharov, D.; Tonevitsky, A.; Lauster, R. ‘Human-on-a-chip’ developments: A translational cutting-edge alternative to systemic safety assessment and efficiency evaluation of substances in laboratory animals and man? Altern. Lab. Anim. 2012, 40, 235–257. [Google Scholar] [PubMed]
- Zheng, F.; Fu, F.; Cheng, Y.; Wang, C.; Zhao, Y.; Gu, Z. Organ-on-a-chip systems: Microengineering to biomimic living systems. Small 2016, 12, 2253–2282. [Google Scholar] [CrossRef] [PubMed]
- Hutmacher, D.W.; Schantz, J.T.; Lam, C.X.F.; Tan, K.C.; Lim, T.C. State of the art and future directions of scaffold-based bone engineering from a biomaterials perspective. J. Tissue Eng. Regen. Med. 2007, 1, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Hydrogels for tissue engineering. Chem. Rev. 2001, 101, 1869–1879. [Google Scholar] [CrossRef] [PubMed]
- Rauh, J.; Milan, F.; Gunther, K.P.; Stiehler, M. Bioreactor systems for bone tissue engineering. Tissue Eng. Part B Rev. 2011, 17, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Robey, P.G. Cell sources for bone regeneration: The good, the bad and the ugly (but promising). Tissue Eng. Part B Rev. 2011, 17, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3, S131–S139. [Google Scholar] [CrossRef] [PubMed]
- Jez, M.; Rozman, P.; Ivanovic, Z.; Bas, T. Concise review: The role of oxygen in hematopoietic stem cell physiology. J. Cell. Physiol. 2015, 230, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.J.; Lanyon, L.E. Mechanical strain and bone cell function: A review. Osteoporos. Int. 2002, 13, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Datta, H.K.; Ng, W.F.; Walker, J.A.; Tuck, S.P.; Varanasi, S.S. The cell biology of bone metabolism. J. Clin. Pathol. 2008, 61, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Robling, A.G.; Castillo, A.B.; Turner, C.H. Biomechanical and molecular regulation of bone remodelling. Annu. Rev. Biomed. Eng. 2006, 8, 455–498. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, H.P.; Meyer, U.; Plate, U.; Hohling, H.J. Aspects of collagen mineralization in hard tissue formation. Int. Rev. Cytol. 2005, 242, 121–156. [Google Scholar] [PubMed]
- Buckwalter, J.A.; Glimcher, M.J.; Cooper, R.R.; Recker, R. Bone biology. Part 1: Structure, blood-supply, cells, matrix and mineralization. J. Bone Jt. Surg. Am. 1995, 77, 1256–1275. [Google Scholar] [CrossRef]
- Frost, H.M. Bone’s mechanostat: A 2003 update. Anat. Rec. Part A 2003, 275, 1081–1101. [Google Scholar] [CrossRef] [PubMed]
- Boulais, P.E.; Frenette, P.S. Making sense of hematopoietic stem cell niches. Blood 2015, 125, 2621–2629. [Google Scholar] [CrossRef] [PubMed]
- Marks, S.C.; Popoff, S.N. Bone cell biology—The regulation of development, structure and function in the skeleton. Am. J. Anat. 1988, 183, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Franz-Odendaal, T.A.; Hall, B.K.; Witten, P.E. Buried alive: How osteoblasts become osteocytes. Dev. Dyn. 2006, 235, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Tate, M.L.K.; Adamson, J.R.; Tami, A.E.; Bauer, T.W. The osteocyte. Int. J. Biochem. Cell B 2004, 36, 1–8. [Google Scholar] [CrossRef]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL–RANK signalling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Vaananen, H.K.; Laitala-Leinonen, T. Osteoclast lineage and function. Arch. Biochem. Biophys. 2008, 473, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell B 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, A.D.; Olsen, B.R. Bone development. Bone 2015, 80, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, G.G.; Ransom, R.C.; Zielins, E.R.; Leavitt, T.; Flacco, J.S.; Hu, M.S.; Lee, A.S.; Longaker, M.T.; Wan, D.C. Stem cells in bone regeneration. Stem Cell Rev. 2016, 12, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Jager, M.; Jelinek, E.M.; Wess, K.M.; Scharfstadt, A.; Jacobson, M.; Kevy, S.V.; Krauspe, R. Bone marrow concentrate: A novel strategy for bone defect treatment. Curr. Stem Cell Res. Ther. 2009, 4, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Ruetze, M.; Richter, W. Adipose-derived stromal cells for osteoarticular repair: Trophic function versus stem cell activity. Expert Rev. Mol. Med. 2014, 16, e9. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.T.; Chen, C.T. Osteogenic potential: Comparison between bone marrow and adipose-derived mesenchymal stem cells. World J. Stem Cells 2014, 6, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Al-Nbaheen, M.; Vishnubalaji, R.; Ali, D.; Bouslimi, A.; Al-Jassir, F.; Megges, M.; Prigione, A.; Adjaye, J.; Kassem, M.; Aldahmash, A. Human stromal (mesenchymal) stem cells from bone marrow, adipose tissue and skin exhibit differences in molecular phenotype and differentiation potential. Stem Cell Rev. 2013, 9, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Han, I.; Kwon, B.S.; Park, H.K.; Kim, K.S. Differentiation potential of mesenchymal stem cells is related to their intrinsic mechanical properties. Int. Neurourol. J. 2017, 21, S24–S31. [Google Scholar] [CrossRef] [PubMed]
- Noel, D.; Caton, D.; Roche, S.; Bony, C.; Lehmann, S.; Casteilla, L.; Jorgensen, C.; Cousin, B. Cell specific differences between human adipose-derived and mesenchymal-stromal cells despite similar differentiation potentials. Exp. Cell Res. 2008, 314, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jia, Z.; Yin, X.; Zhang, X.; Liu, Y.; Chen, P.; Ma, K.; Zhou, C. Comparative analysis of mesenchymal stem cells from bone marrow, cartilage and adipose tissue. Stem Cells Dev. 2008, 17, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Rebelatto, C.K.; Aguiar, A.M.; Moretao, M.P.; Senegaglia, A.C.; Hansen, P.; Barchiki, F.; Oliveira, J.; Martins, J.; Kuligovski, C.; Mansur, F.; et al. Dissimilar differentiation of mesenchymal stem cells from bone marrow, umbilical cord blood and adipose tissue. Exp. Biol. Med. (Maywood) 2008, 233, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Geissler, S.; Textor, M.; Kuhnisch, J.; Konnig, D.; Klein, O.; Ode, A.; Pfitzner, T.; Adjaye, J.; Kasper, G.; Duda, G.N. Functional comparison of chronological and in vitro aging: Differential role of the cytoskeleton and mitochondria in mesenchymal stromal cells. PLoS ONE 2012, 7, e52700. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Akintoye, S.O.; Lam, T.; Shi, S.; Brahim, J.; Collins, M.T.; Robey, P.G. Skeletal site-specific characterization of orofacial and iliac crest human bone marrow stromal cells in same individuals. Bone 2006, 38, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Takase, K.; Suehiro, F.; Murata, H. Candidates cell sources to regenerate alveolar bone from oral tissue. Int. J. Dent. Oral Health 2012, 2012, 857192. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Rosada, C.; Justesen, J.; Melsvik, D.; Ebbesen, P.; Kassem, M. The human umbilical cord blood: A potential source for osteoblast progenitor cells. Calcif. Tissue Int. 2003, 72, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Campagnoli, C.; Roberts, I.A.; Kumar, S.; Bennett, P.R.; Bellantuono, I.; Fisk, N.M. Identification of mesenchymal stem/progenitor cells in human first-trimester foetal blood, liver and bone marrow. Blood 2001, 98, 2396–2402. [Google Scholar] [CrossRef] [PubMed]
- Toma, J.G.; McKenzie, I.A.; Bagli, D.; Miller, F.D. Isolation and characterization of multipotent skin-derived precursors from human skin. Stem Cells 2005, 23, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, E.; Morita, R.; Nakao, K.; Ishida, K.; Nakamura, T.; Takano-Yamamoto, T.; Ogawa, M.; Mizuno, M.; Kasugai, S.; Tsuji, T. Fully functional bioengineered tooth replacement as an organ replacement therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 13475–13480. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Yamato, M.; Zhang, Z.; Mukobata, S.; Washio, K.; Ando, T.; Feijen, J.; Okano, T.; Ishikawa, I. Validation of human periodontal ligament-derived cells as a reliable source for cytotherapeutic use. J. Clin Periodontol. 2010, 37, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Seeberger, K.L.; Dufour, J.M.; Shapiro, A.M.; Lakey, J.R.; Rajotte, R.V.; Korbutt, G.S. Expansion of mesenchymal stem cells from human pancreatic ductal epithelium. Lab. Investig. 2006, 86, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Wenceslau, C.V.; Miglino, M.A.; Martins, D.S.; Ambrosio, C.E.; Lizier, N.F.; Pignatari, G.C.; Kerkis, I. Mesenchymal progenitor cells from canine foetal tissues: Yolk sac, liver and bone marrow. Tissue Eng. Part A 2011, 17, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Capulli, M.; Paone, R.; Rucci, N. Osteoblast and osteocyte: Games without frontiers. Arch. Biochem. Biophys. 2014, 561, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Prideaux, M.; Schutz, C.; Wijenayaka, A.R.; Findlay, D.M.; Campbell, D.G.; Solomon, L.B.; Atkins, G.J. Isolation of osteocytes from human trabecular bone. Bone 2016, 88, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, K.B.; Frost, A.; Nilsson, O.; Ljunghall, S.; Ljunggren, O. Three isolation techniques for primary culture of human osteoblast-like cells: A comparison. Acta Orthop. Scand. 1999, 70, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, S.; Gehron Robey, P. Species differences in growth requirements for bone marrow stromal fibroblast colony formation in vitro. Calcif. Tissue Int. 1996, 59, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Eghbali-Fatourechi, G.Z.; Modder, U.I.; Charatcharoenwitthaya, N.; Sanyal, A.; Undale, A.H.; Clowes, J.A.; Tarara, J.E.; Khosla, S. Characterization of circulating osteoblast lineage cells in humans. Bone 2007, 40, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Colnot, C. Cell sources for bone tissue engineering: Insights from basic science. Tissue Eng. Part B Rev. 2011, 17, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Reinke, S.; Dienelt, A.; Blankenstein, A.; Duda, G.N.; Geissler, S. Qualifying stem cell sources: How to overcome potential pitfalls in regenerative medicine? J. Tissue Eng. Regen. Med. 2016, 10, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.; Horowitz, M.; Choi, Y. Osteoimmunology: Interactions of the bone and immune system. Endocr. Rev. 2008, 29, 403–440. [Google Scholar] [CrossRef] [PubMed]
- Reinke, S.; Geissler, S.; Taylor, W.R.; Schmidt-Bleek, K.; Juelke, K.; Schwachmeyer, V.; Dahne, M.; Hartwig, T.; Akyuz, L.; Meisel, C.; et al. Terminally differentiated CD8⁺ T cells negatively affect bone regeneration in humans. Sci. Transl. Med. 2013, 5, 177ra136. [Google Scholar] [CrossRef] [PubMed]
- Marolt, D.; Campos, I.M.; Bhumiratana, S.; Koren, A.; Petridis, P.; Zhang, G.; Spitalnik, P.F.; Grayson, W.L.; Vunjak-Novakovic, G. Engineering bone tissue from human embryonic stem cells. Proc. Natl Acad. Sci USA 2012, 109, 8705–8709. [Google Scholar] [CrossRef] [PubMed]
- Csobonyeiova, M.; Polak, S.; Zamborsky, R.; Danisovic, L. Ips cell technologies and their prospect for bone regeneration and disease modeling: A mini review. J. Adv. Res. 2017, 8, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Ralphs, J.R.; Richards, R.G.; Hayes, J.S. A phenotypic comparison of osteoblast cell lines versus human primary osteoblasts for biomaterials testing. J. Biomed. Mater. Res. A 2014, 102, 2636–2643. [Google Scholar] [CrossRef] [PubMed]
- Kartsogiannis, V.; Ng, K.W. Cell lines and primary cell cultures in the study of bone cell biology. Mol. Cell. Endocrinol 2004, 228, 79–102. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Boskey, A.; Spevak, L.; Dallas, M.; Hori, M.; Bonewald, L.F. Establishment of an osteoid preosteocyte-like cell MLO-A5that spontaneously mineralizes in culture. J. Bone Miner. Res. 2001, 16, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Achilli, T.-M.; Meyer, J.; Morgan, J.R. Advances in the formation, use and understanding of multi-cellular spheroids. Expert Opin. Biol. Ther. 2012, 12, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Menger, M.D. Life is 3d: Boosting spheroid function for tissue engineering. Trends Biotechnol. 2017, 35, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Cuddihy, M.J.; Cater, G.M.; Kotov, N.A. Engineering liver tissue spheroids with inverted colloidal crystal scaffolds. Biomaterials 2009, 30, 4687–4694. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.J.; Friend, J.R.; Hsiao, C.C.; Zilliox, M.J.; Ko, W.J.; Cerra, F.B.; Hu, W.S. Efficient assembly of rat hepatocyte spheroids for tissue engineering applications. Biotechnol. Bioeng. 1996, 50, 404–415. [Google Scholar] [CrossRef]
- Pamies, D.; Wiersma, D.M.M.; Block, K.; Pardo, C.A.; Barreras, P.; Kyro, K.; Leist, M.; Hartung, T.; Hogberg, H. A 3-D in vitro mini-brain model to study Parkinson’s disease. Birth Defects Res. A 2015, 103, 394. [Google Scholar]
- Hirschhaeuser, F.; Menne, H.; Dittfeld, C.; West, J.; Mueller-Klieser, W.; Kunz-Schughart, L.A. Multicellular tumour spheroids: An underestimated tool is catching up again. J. Biotechnol. 2010, 148, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Kelm, J.M.; Timmins, N.E.; Brown, C.J.; Fussenegger, M.; Nielsen, L.K. Method for generation of homogeneous multicellular tumour spheroids applicable to a wide variety of cell types. Biotechnol. Bioeng. 2003, 83, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, C.; Büth, H.; Thielecke, H. A scaffold-free in vitro model for osteogenesis of human mesenchymal stem cells. Tissue Cell 2011, 43, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Metzger, W.; Schimmelpfennig, L.; Schwab, B.; Sossong, D.; Dorst, N.; Bubel, M.; Gorg, A.; Putz, N.; Wennemuth, G.; Pohlemann, T.; et al. Expansion and differentiation of human primary osteoblasts in two- and three-dimensional culture. Biotech. Histochem. 2013, 88, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.C.; Henstock, J.R.; Reinwald, Y.; El Haj, A.J. Dynamic 3D culture: Models of chondrogenesis and endochondral ossification. Birth Defects Res. C Embryo Today 2015, 105, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, J.I.; Matsumoto, T.; Egusa, H.; Matsusaki, M.; Nishiguchi, A.; Nakano, T.; Akashi, M.; Imazato, S.; Yatani, H. In vitro reproduction of endochondral ossification using a 3D mesenchymal stem cell construct. Integr. Biol.-UK 2012, 4, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Kapur, S.K.; Wang, X.; Shang, H.; Yun, S.; Li, X.; Feng, G.; Khurgel, M.; Katz, A.J. Human adipose stem cells maintain proliferative, synthetic and multipotential properties when suspension cultured as self-assembling spheroids. Biofabrication 2012, 4, 025004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, P.; Chen, L.; Wang, Y.; Wang, Z.; Zhang, B. The effects of spheroid formation of adipose-derived stem cells in a microgravity bioreactor on stemness properties and therapeutic potential. Biomaterials 2015, 41, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Gurumurthy, B.; Bierdeman, P.C.; Janorkar, A.V. Spheroid model for functional osteogenic evaluation of human adipose derived stem cells. J. Biomed. Mater. Res. A 2017, 105, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- de Barros, A.P.; Takiya, C.M.; Garzoni, L.R.; Leal-Ferreira, M.L.; Dutra, H.S.; Chiarini, L.B.; Meirelles, M.N.; Borojevic, R.; Rossi, M.I. Osteoblasts and bone marrow mesenchymal stromal cells control hematopoietic stem cell migration and proliferation in 3D in vitro model. PLoS ONE 2010, 5, e9093. [Google Scholar] [CrossRef] [PubMed]
- Papadimitropoulos, A.; Scherberich, A.; Guven, S.; Theilgaard, N.; Crooijmans, H.J.; Santini, F.; Scheffler, K.; Zallone, A.; Martin, I. A 3D in vitro bone organ model using human progenitor cells. Eur. Cell Mater. 2011, 21, 445–458, discussion 458. [Google Scholar] [CrossRef] [PubMed]
- Wenger, A.; Stahl, A.; Weber, H.; Finkenzeller, G.; Augustin, H.G.; Stark, G.B.; Kneser, U. Modulation of in vitro angiogenesis in a three-dimensional spheroidal coculture model for bone tissue engineering. J. Tissue Eng. 2004, 10, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.R.; Stegemann, J.P. Cell-based approaches to the engineering of vascularized bone tissue. Cytotherapy 2013, 15, 1309–1322. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ye, Y.; Tian, H.; Yang, S.; Jin, X.; Tong, W.; Zhang, Y. In vitro osteogenesis of human adipose-derived stem cells by coculture with human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun. 2011, 412, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Laranjeira, M.S.; Fernandes, M.H.; Monteiro, F.J. Reciprocal induction of human dermal microvascular endothelial cells and human mesenchymal stem cells: Time-dependent profile in a co-culture system. Cell Prolif. 2012, 45, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.; Lampert, F.; Stark, G.B.; Finkenzeller, G. Effects of endothelial cells on proliferation and survival of human mesenchymal stem cells and primary osteoblasts. J. Orthop. Res. 2012, 30, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Leszczynska, J.; Zyzynska-Granica, B.; Koziak, K.; Ruminski, S.; Lewandowska-Szumiel, M. Contribution of endothelial cells to human bone-derived cells expansion in coculture. J. Tissue Eng. Part A 2013, 19, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Guillotin, B.; Bareille, R.; Bourget, C.; Bordenave, L.; Amedee, J. Interaction between human umbilical vein endothelial cells and human osteoprogenitors triggers pleiotropic effect that may support osteoblastic function. Bone 2008, 42, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Xing, Z.; Hellem, S.; Arvidson, K.; Mustafa, K. Endothelial cells influence the osteogenic potential of bone marrow stromal cells. Biomed. Eng. Online 2009, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.Y.; Li, Q.; Bai, N.N.; Dong, H.Z.; Li, D.K. Porous stable poly(lactic acid)/ethyl cellulose/hydroxyapatite composite scaffolds prepared by a combined method for bone regeneration. Carbohydr. Polym. 2018, 180, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.L.; Liu, X.M.; Yeung, K.W.K.; Liu, C.S.; Yang, X.J. Biomimetic porous scaffolds for bone tissue engineering. Mater. Sci. Eng. R 2014, 80, 1–36. [Google Scholar] [CrossRef]
- Liu, Y.C.; Lim, J.; Teoh, S.H. Review: Development of clinically relevant scaffolds for vascularised bone tissue engineering. Biotechnol. Adv. 2013, 31, 688–705. [Google Scholar] [CrossRef] [PubMed]
- Hutmacher, D.W.; Garcia, A.J. Scaffold-based bone engineering by using genetically modified cells. Gene 2005, 347, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Yang, Y.Z.; MohandaS, A.; Stucker, B.; Nguyen, K.T. A review of materials, fabrication to enhance bone regeneration in methods and strategies used engineered bone tissues. J. Biomed. Mater. Res. B 2008, 85, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Muschler, G.E.; Nakamoto, C.; Griffith, L.G. Engineering principles of clinical cell-based tissue engineering. J. Bone Jt. Surg. Am. 2004, 86, 1541–1558. [Google Scholar] [CrossRef]
- Bancroft, G.N.; Sikavitsas, V.I.; van den Dolder, J.; Sheffield, T.L.; Ambrose, C.G.; Jansen, J.A.; Mikos, A.G. Fluid flow increases mineralized matrix deposition in 3D perfusion culture of marrow stromal osteoblasts in a dose-dependent manner. Proc. Natl. Acad. Sci. USA 2002, 99, 12600–12605. [Google Scholar] [CrossRef] [PubMed]
- Planell, J.A.; Best, S.M.; Lacroix, D.; Merolli, A. (Eds.) Bone Repair Biomaterials; Woodhead Publishing: Sawston, UK, 2009; pp. 1–478. [Google Scholar]
- Mitra, D.; Whitehead, J.; Yasui, O.W.; Leach, J.K. Bioreactor culture duration of engineered constructs influences bone formation by mesenchymal stem cells. Biomaterials 2017, 146, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Albrektsson, T.; Johansson, C. Osteoinduction, osteoconduction and osseointegration. Eur. Spine J. 2001, 10, S96–S101. [Google Scholar] [PubMed]
- Li, Z.; Gong, Y.W.; Sun, S.J.; Du, Y.; Lu, D.Y.; Liu, X.F.; Long, M. Differential regulation of stiffness, topography and dimension of substrates in rat mesenchymal stem cells. Biomaterials 2013, 34, 7616–7625. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Roy, M.; Bandyopadhyay, A. Recent advances in bone tissue engineering scaffolds. Trends Biotechnol. 2012, 30, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.M.; Matyas, J.R.; Katzenberg, M.A.; Hallgrimsson, B. Comparison of microcomputed tomographic and microradiographic measurements of cortical bone porosity. Calcif. Tissue Int. 2004, 74, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Mikael, P.E.; Nukavarapu, S.P. Functionalized carbon nanotube composite scaffolds for bone tissue engineering: Prospects and progress. J. Biomater. Tissue Eng. 2011, 1, 76–85. [Google Scholar] [CrossRef]
- Murphy, C.M.; Haugh, M.G.; O’Brien, F.J. The effect of mean pore size on cell attachment, proliferation and migration in collagen–glycosaminoglycan scaffolds for bone tissue engineering. Biomaterials 2010, 31, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Hoshiba, T.; Kawazoe, N.; Koda, I.; Song, M.; Chen, G. Cultured cell-derived extracellular matrix scaffolds for tissue engineering. Biomaterials 2011, 32, 9658–9666. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Shi, Y.; Wei, X.; He, J.; Yang, S.; Dickson, G.; Tang, J.; Xiang, J.; Song, C.; Li, G. Fabrication and repair of cartilage defects with a novel acellular cartilage matrix scaffold. J. Tissue Eng. Part C Methods 2010, 16, 865–876. [Google Scholar] [CrossRef] [PubMed]
- McDuffee, L.A.; Esparza Gonzalez, B.P.; Nino-Fong, R.; Aburto, E. Evaluation of an in vivo heterotopic model of osteogenic differentiation of equine bone marrow and muscle mesenchymal stem cells in fibrin glue scaffold. Cell Tissue Res. 2014, 355, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, J.; Cao, L.; Qian, X.; Xing, W.; Lu, J.; Liu, C. Segmental bone regeneration using rhbmp-2-loaded collagen/chitosan microspheres composite scaffold in a rabbit model. Biomed. Mater. 2012, 7, 035002. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Kamali, A.; Moshiri, A.; Baghaban Eslaminejad, M. Role of mesenchymal stem cells in bone regenerative medicine: What is the evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.X.; He, Y.; Bi, L.; Qu, Z.H.; Zou, J.W.; Pan, Z.; Fan, J.J.; Chen, L.; Dong, X.; Liu, X.N.; et al. Enhancing the bioactivity of poly (lactic-co-glycolic acid) scaffold with a nano-hydroxyapatite coating for the treatment of segmental bone defect in a rabbit model. Int. J. Nanomed. 2013, 8, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zheng, Q.; Guo, X.; Wu, Y.; Wu, B. Polydopamine-assisted BMP-2-derived peptides immobilization on biomimetic copolymer scaffold for enhanced bone induction in vitro and in vivo. Colloids Surf. B Biointerfaces 2016, 142, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Son, S.J.; Son, J.S.; Kang, S.S.; Choi, S.H. Bone-healing capacity of PCL/PLGA/duck beak scaffold in critical bone defects in a rabbit model. Biomed. Res. Int. 2016, 2016, 2136215. [Google Scholar] [CrossRef] [PubMed]
- Almela, T.; Al-Sahaf, S.; Bolt, R.; Brook, I.M.; Moharamzadeh, K. Characterization of multilayered tissue-engineered human alveolar bone and gingival mucosa. Tissue Eng. Part C Methods 2018, 24, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Masaoka, T.; Yoshii, T.; Yuasa, M.; Yamada, T.; Taniyama, T.; Torigoe, I.; Shinomiya, K.; Okawa, A.; Morita, S.; Sotome, S. Bone defect regeneration by a combination of a β-tricalcium phosphate scaffold and bone marrow stromal cells in a non-human primate model. Open Biomed. Eng. J. 2016, 10, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Balla, V.K.; Bodhak, S.; Bose, S.; Bandyopadhyay, A. Porous tantalum structures for bone implants: Fabrication, mechanical and in vitro biological properties. Acta Biomater. 2010, 6, 3349–3359. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Krishna, B.V.; Bandyopadhyay, A.; Bose, S. Processing and biocompatibility evaluation of laser processed porous titanium. Acta Biomater. 2007, 3, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Hashemi, B. Co-existence effect of tricalcium phosphate and bioactive glass on biological and biodegradation characteristic of poly L-lactic acid (PLLA) in trinary composite scaffold form. Biomed. Mater. Eng. 2017, 28, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Todo, M.; Arahira, T. In vitro bone formation by mesenchymal stem cells with 3d collagen/beta-tcp composite scaffold. In Proceedings of the 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Osaka Japan, 3–7 July 2013; pp. 409–412. [Google Scholar]
- Kim, J.M.; Son, J.S.; Kang, S.S.; Kim, G.; Choi, S.H. Bone regeneration of hydroxyapatite/alumina bilayered scaffold with 3 mm passage-like medullary canal in canine tibia model. Biomed. Res. Int. 2015, 2015, 235108. [Google Scholar] [CrossRef] [PubMed]
- García-Gareta, E.; Coathup, M.J.; Blunn, G.W. Osteoinduction of bone grafting materials for bone repair and regeneration. Bone 2015, 81, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Basha, R.Y.; Doble, M. Design of biocomposite materials for bone tissue regeneration. Mater. Sci. Eng. C 2015, 57, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.Y.; Kuhn-Spearing, L.; Zioupos, P. Mechanical properties and the hierarchical structure of bone. Med. Eng. Phys. 1998, 20, 92–102. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Botelho, M.G.; Dorozhkin, S.V. Biphasic calcium phosphates bioceramics (HA/TCP): Concept, physicochemical properties and the impact of standardization of study protocols in biomaterials research. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 71, 1293–1312. [Google Scholar] [CrossRef] [PubMed]
- Samavedi, S.; Whittington, A.R.; Goldstein, A.S. Calcium phosphate ceramics in bone tissue engineering: A review of properties and their influence on cell behaviour. Acta Biomater. 2013, 9, 8037–8045. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Myoui, A. Bone tissue engineering with porous hydroxyapatite ceramics. J. Artif. Organs 2005, 8, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Meskinfam, M.; Bertoldi, S.; Albanese, N.; Cerri, A.; Tanzi, M.C.; Imani, R.; Baheiraei, N.; Farokhi, M.; Fare, S. Polyurethane foam/nano hydroxyapatite composite as a suitable scaffold for bone tissue regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 82, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, S.; Ma, Y.; Xu, W.; Meng, W.; Lin, X.; Wang, W.; Wang, S.; Zhang, J. Innovative biodegradable poly(l-lactide)/collagen/hydroxyapatite composite fibrous scaffolds promote osteoblastic proliferation and differentiation. Int. J. Nanomed. 2017, 12, 7577–7588. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Haque, M.; Kearney, M.T.; Lopez, M.J. Collagen and hydroxyapatite scaffolds activate distinct osteogenesis signalling pathways in adult adipose-derived multipotent stromal cells. Tissue Eng. Part. C Methods 2017, 23, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, R.A.; Mazor, Z.; Miller, R.J.; Krauser, J.; Prasad, H.S.; Rohrer, M.D. Clinical evaluation alveolar ridge preservation with a beta-tricalcium phosphate socket graft. Compend. Contin. Educ. Dent. 2009, 30, 588–590, 592, 594 passim; quiz 604, 606. [Google Scholar] [PubMed]
- Bernhardt, A.; Lode, A.; Peters, F.; Gelinsky, M. Optimization of culture conditions for osteogenically-induced mesenchymal stem cells in β-tricalcium phosphate ceramics with large interconnected channels. J. Tissue Eng. Regen. Med. 2011, 5, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Jiang, C.; Li, C.; Li, T.; Peng, M.; Wang, J.; Dai, K. 3D printed scaffolds of calcium silicate-doped β-tcp synergize with co-cultured endothelial and stromal cells to promote vascularization and bone formation. Sci. Rep. 2017, 7, 5588. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Ren, L.; Yang, Y. Engineering vascularized bone grafts by integrating a biomimetic periosteum and β-TCP scaffold. ACS Appl. Mater. Interfaces 2014, 6, 9622–9633. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.; Ito, T.; Bishop, D.; Otsuka, M.; Ben-Nissan, B.; Milthorpe, B. Controlled release of simvastatin from biomimetic beta-tcp drug delivery system. PLoS ONE 2013, 8, e54676. [Google Scholar]
- Shuai, C.; Cao, Y.; Dan, G.; Gao, C.; Feng, P.; Wu, P. Improvement in degradability of 58s glass scaffolds by zno and beta-tcp modification. Bioengineered 2016, 7, 342–351. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Boukhechba, F.; Balaguer, T.; Michiels, J.F.; Ackermann, K.; Quincey, D.; Bouler, J.M.; Pyerin, W.; Carle, G.F.; Rochet, N. Human primary osteocyte differentiation in a 3D culture system. J. Bone Miner. Res. 2009, 24, 1927–1935. [Google Scholar] [CrossRef] [PubMed]
- Zima, A.; Czechowska, J.; Siek, D.; Olkowski, R.; Noga, M.; Lewandowska-Szumiel, M.; Slosarczyk, A. How calcite and modified hydroxyapatite influence physicochemical properties and cytocompatibility of α-TCP based bone cements. J. Mater. Sci. Mater. Med. 2017, 28, 117. [Google Scholar] [CrossRef] [PubMed]
- Wichterle, O.; Lim, D. Hydrophilic gels for biological use. Nature 1960, 185, 117–118. [Google Scholar] [CrossRef]
- Dong, L.; Wang, S.J.; Zhao, X.R.; Zhu, Y.F.; Yu, J.K. 3D- printed poly(ε-caprolactone) scaffold integrated with cell-laden chitosan hydrogels for bone tissue engineering. Sci. Rep. 2017, 7, 13412. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, S.; Maloney, R.; Chakravarti, A.R.; Whitlow, J.; Basu, S.; Modaresi, S.; Gehrke, S.; Paul, A. Controlling adult stem cell behavior using nanodiamond-reinforced hydrogel: Implication in bone regeneration therapy. Sci. Rep. 2017, 7, 6577. [Google Scholar] [CrossRef] [PubMed]
- Dhivya, S.; Saravanan, S.; Sastry, T.P.; Selvamurugan, N. Nanohydroxyapatite-reinforced chitosan composite hydrogel for bone tissue repair in vitro and in vivo. J. Nanobiotechnol. 2015, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Hoare, T.R.; Kohane, D.S. Hydrogels in drug delivery: Progress and challenges. Polymer 2008, 49, 1993–2007. [Google Scholar] [CrossRef]
- Jhon, M.S.; Andrade, J.D. Water and hydrogels. Journal of biomedical materials research 1973, 7, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Noori, A.; Ashrafi, S.J.; Vaez-Ghaemi, R.; Hatamian-Zaremi, A.; Webster, T.J. A review of fibrin and fibrin composites for bone tissue engineering. Int. J. Nanomed. 2017, 12, 4937–4961. [Google Scholar] [CrossRef] [PubMed]
- Jen, A.C.; Wake, M.C.; Mikos, A.G. Review: Hydrogels for cell immobilization. Biotechnol. Bioeng. 1996, 50, 357–364. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, H.; Moon, S.; Na, K. Bone morphogenic protein-2 (BMP-2) loaded nanoparticles mixed with human mesenchymal stem cell in fibrin hydrogel for bone tissue engineering. J. Biosci. Bioeng. 2009, 108, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Ikada, Y.; Tabata, Y. Controlled release of growth factors based on biodegradation of gelatin hydrogel. J. Biomater. Sci. Polym. Ed. 2001, 12, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kempen, D.H.; Lu, L.; Hefferan, T.E.; Creemers, L.B.; Maran, A.; Classic, K.L.; Dhert, W.J.; Yaszemski, M.J. Retention of in vitro and in vivo BMP-2 bioactivities in sustained delivery vehicles for bone tissue engineering. Biomaterials 2008, 29, 3245–3252. [Google Scholar] [CrossRef] [PubMed]
- Akagawa, Y.; Kubo, T.; Koretake, K.; Hayashi, K.; Doi, K.; Matsuura, A.; Morita, K.; Takeshita, R.; Yuan, Q.; Tabata, Y. Initial bone regeneration around fenestrated implants in beagle dogs using basic fibroblast growth factor-gelatin hydrogel complex with varying biodegradation rates. J. Prosthodont. Res. 2009, 53, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ratanavaraporn, J.; Furuya, H.; Kohara, H.; Tabata, Y. Synergistic effects of the dual release of stromal cell-derived factor-1 and bone morphogenetic protein-2 from hydrogels on bone regeneration. Biomaterials 2011, 32, 2797–2811. [Google Scholar] [CrossRef] [PubMed]
- Laurent, T.C.; Laurent, U.B.; Fraser, J.R. Functions of hyaluronan. Ann. Rheum. Dis. 1995, 54, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Docherty-Skogh, A.C.; Bergman, K.; Waern, M.J.; Ekman, S.; Hultenby, K.; Ossipov, D.; Hilborn, J.; Bowden, T.; Engstrand, T. Bone morphogenetic protein-2 delivered by hyaluronan-based hydrogel induces massive bone formation and healing of cranial defects in minipigs. Plast. Reconstr. Surg. 2010, 125, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, I.S.; Cho, T.H.; Lee, K.B.; Hwang, S.J.; Tae, G.; Noh, I.; Lee, S.H.; Park, Y.; Sun, K. Bone regeneration using hyaluronic acid-based hydrogel with bone morphogenic protein-2 and human mesenchymal stem cells. Biomaterials 2007, 28, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Sanz, E.; Ossipov, D.A.; Hilborn, J.; Larsson, S.; Jonsson, K.B.; Varghese, O.P. Bone reservoir: Injectable hyaluronic acid hydrogel for minimal invasive bone augmentation. J. Control. Release 2011, 152, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Jo, S.; Mikos, A.G. Modulation of marrow stromal osteoblast adhesion on biomimetic oligo[poly(ethylene glycol) fumarate] hydrogels modified with arg–gly–asp peptides and a poly(ethyleneglycol) spacer. J. Biomed. Mater. Res. 2002, 61, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Williams, C.G.; Wang, D.A.; Lee, H.; Manson, P.N.; Elisseeff, J. The effect of incorporating RGD adhesive peptide in polyethylene glycol diacrylate hydrogel on osteogenesis of bone marrow stromal cells. Biomaterials 2005, 26, 5991–5998. [Google Scholar] [CrossRef] [PubMed]
- Peled, E.; Boss, J.; Bejar, J.; Zinman, C.; Seliktar, D. A novel poly(ethylene glycol)-fibrinogen hydrogel for tibial segmental defect repair in a rat model. J. Biomed. Mater. Res. A 2007, 80, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Ni, P.; Wang, B.; Chu, B.; Zheng, L.; Luo, F.; Luo, J.; Qian, Z. Injectable and thermo-sensitive PEG–PCL–PEG copolymer/collagen/n–HA hydrogel composite for guided bone regeneration. Biomaterials 2012, 33, 4801–4809. [Google Scholar] [CrossRef] [PubMed]
- Demirtas, T.T.; Irmak, G.; Gumusderelioglu, M. A bioprintable form of chitosan hydrogel for bone tissue engineering. Biofabrication. 2017, 9, 035003. [Google Scholar] [CrossRef] [PubMed]
- Igwe, J.C.; Mikael, P.E.; Nukavarapu, S.P. Design, fabrication and in vitro evaluation of a novel polymer-hydrogel hybrid scaffold for bone tissue engineering. J. Tissue Eng. Regen. Med. 2014, 8, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Lee, J.H.; Yun, J.W.; Park, J.H.; Park, B.W.; Rho, G.J.; Jang, S.J.; Park, J.S.; Lee, H.C.; Yoon, Y.M.; et al. Development of porous beads to provide regulated BMP-2 stimulation for varying durations: In vitro and in vivo studies for bone regeneration. Biomacromolecules 2016, 17, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, T.; Kabiraj, P.; Narayana, G.H.; Kulanthaivel, S.; Kasiviswanathan, U.; Pal, K.; Giri, S.; Maiti, T.K.; Banerjee, I. Alginate bead based hexagonal close packed 3D implant for bone tissue engineering. ACS Appl. Mater. Interfaces 2016, 8, 32132–32145. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Hoch, A.I.; Decaris, M.L.; Leach, J.K. Alginate hydrogels containing cell-interactive beads for bone formation. FASEB J. 2013, 27, 4844–4852. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.; Wang, C.; Yuan, X.; Liu, Z.; Dziak, R.; Yang, S. Combination of controlled release platelet-rich plasma alginate beads and bone morphogenetic protein-2 genetically modified mesenchymal stem cells for bone regeneration. J. Periodontol. 2016, 87, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Asaoka, T.; Ohtake, S.; Furukawa, K.S.; Tamura, A.; Ushida, T. Development of bioactive porous alpha-tcp/hap beads for bone tissue engineering. J. Biomed. Mater. Res. A 2013, 101, 3295–3300. [Google Scholar] [PubMed]
- Bose, S.; Vahabzadeh, S.; Bandyopadhyay, A. Bone tissue engineering using 3D printing. Mater. Today 2013, 16, 496–504. [Google Scholar] [CrossRef]
- Bergmann, C.; Lindner, M.; Zhang, W.; Koczur, K.; Kirsten, A.; Telle, R.; Fischer, H. 3D printing of bone substitute implants using calcium phosphate and bioactive glasses. J. Eur Ceram. Soc. 2010, 30, 2563–2567. [Google Scholar] [CrossRef]
- Ji, S.; Guvendiren, M. Recent advances in bioink design for 3D bioprinting of tissues and organs. Front. Bioeng. Biotechnol. 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Adepu, S.; Dhiman, N.; Laha, A.; Sharma, C.S.; Ramakrishna, S.; Khandelwal, M. Three-dimensional bioprinting for bone tissue regeneration. Curr. Opin. Biomed. Eng. 2017, 2, 22–28. [Google Scholar] [CrossRef]
- Bendtsen, S.T.; Quinnell, S.P.; Wei, M. Development of a novel alginate-polyvinyl alcohol-hydroxyapatite hydrogel for 3D bioprinting bone tissue engineered scaffolds. J. Biomed. Mater. Res. A 2017, 105, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fu, H.; Wang, Z.Y.; Meng, Q.Y.; Liu, S.M.; Wang, H.R.; Zheng, X.F.; Dai, J.W.; Zhang, Z.J. BMSCs-laden gelatin/sodium alginate/carboxymethyl chitosan hydrogel for 3D bioprinting. RSC Adv. 2016, 6, 108423–108430. [Google Scholar] [CrossRef]
- Park, J.Y.; Shim, J.H.; Choi, S.A.; Jang, J.; Kim, M.; Lee, S.H.; Cho, D.W. 3D printing technology to control bmp-2 and vegf delivery spatially and temporally to promote large-volume bone regeneration. J. Mater. Chem. B 2015, 3, 5415–5425. [Google Scholar] [CrossRef]
- Du, M.C.; Chen, B.; Meng, Q.Y.; Liu, S.M.; Zheng, X.F.; Zhang, C.; Wang, H.R.; Li, H.Y.; Wang, N.; Dai, J.W. 3D bioprinting of BMSC-laden methacrylamide gelatin scaffolds with CBD-BMP2-collagen microfibers. Biofabrication. 2015, 7, 044104. [Google Scholar] [CrossRef] [PubMed]
- Kolesky, D.B.; Homan, K.A.; Skylar-Scott, M.A.; Lewis, J.A. Three-dimensional bioprinting of thick vascularized tissues. Proc. Natl. Acad. Sci. USA. 2016, 113, 3179–3184. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Lee, S.J.; Ko, I.K.; Kengla, C.; Yoo, J.J.; Atala, A. A 3D bioprinting system to produce human-scale tissue constructs with structural integrity. Nat. Biotechnol. 2016, 34, 312. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.H.; Lee, J.S.; Kim, J.Y.; Cho, D.W. Bioprinting of a mechanically enhanced three-dimensional dual cell-laden construct for osteochondral tissue engineering using a multi-head tissue/organ building system. J. Micromech. Microeng. 2012, 22, 085014. [Google Scholar] [CrossRef]
- Mandrycky, C.; Wang, Z.J.; Kim, K.; Kim, D.H. 3D bioprinting for engineering complex tissues. Biotechnol. Adv. 2016, 34, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Catros, S.; Fricain, J.C.; Guillotin, B.; Pippenger, B.; Bareille, R.; Remy, M.; Lebraud, E.; Desbat, B.; Amedee, J.; Guillemot, F. Laser-assisted bioprinting for creating on-demand patterns of human osteoprogenitor cells and nano-hydroxyapatite. Biofabrication. 2011, 3, 025001. [Google Scholar] [CrossRef] [PubMed]
- Keriquel, V.; Oliveira, H.; Remy, M.; Ziane, S.; Delmond, S.; Rousseau, B.; Rey, S.; Catros, S.; Amedee, J.; Guillemot, F.; et al. In situ printing of mesenchymal stromal cells, by laser-assisted bioprinting, for in vivo bone regeneration applications. Sci. Rep.-UK 2017, 7, 1778. [Google Scholar] [CrossRef] [PubMed]
- Kleinhans, C.; Mohan, R.R.; Vacun, G.; Schwarz, T.; Haller, B.; Sun, Y.; Kahlig, A.; Kluger, P.; Finne-Wistrand, A.; Walles, H.; et al. A perfusion bioreactor system efficiently generates cell-loaded bone substitute materials for addressing critical size bone defects. Biotechnol. J. 2015, 10, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Wendt, D.; Marsano, A.; Jakob, M.; Heberer, M.; Martin, I. Oscillating perfusion of cell suspensions through three-dimensional scaffolds enhances cell seeding efficiency and uniformity. Biotechnol. Bioeng. 2003, 84, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Scherberich, A.; Galli, R.; Jaquiery, C.; Farhadi, J.; Martin, I. Three-dimensional perfusion culture of human adipose tissue-derived endothelial and osteoblastic progenitors generates osteogenic constructs with intrinsic vascularization capacity. Stem Cells 2007, 25, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Braccini, A.; Wendt, D.; Jaquiery, C.; Jakob, M.; Heberer, M.; Kenins, L.; Wodnar-Filipowicz, A.; Quarto, R.; Martin, I. Three-dimensional perfusion culture of human bone marrow cells and generation of osteoinductive grafts. Stem Cells 2005, 23, 1066–1072. [Google Scholar] [CrossRef] [PubMed]
- Grayson, W.L.; Frohlich, M.; Yeager, K.; Bhumiratana, S.; Chan, M.E.; Cannizzaro, C.; Wan, L.Q.; Liu, X.S.; Guo, X.E.; Vunjak-Novakovic, G. Engineering anatomically shaped human bone grafts. Proc. Natl. Acad. Sci. USA 2010, 107, 3299–3304. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, E.; Finoli, A.; Nettleship, I.; Gerlach, J.C. Long-term three-dimensional perfusion culture of human adult bone marrow mononuclear cells in bioreactors. Biotechnol. Bioeng. 2015, 112, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; Bacabac, R.G.; Bakker, A.D. Mechanical loading and how it affects bone cells: The role of the osteocyte cytoskeleton in maintaining our skeleton. Eur. Cell Mater. 2012, 24, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Matziolis, G.; Tuischer, J.; Kasper, G.; Thompson, M.; Bartmeyer, B.; Krocker, D.; Perka, C.; Duda, G. Simulation of cell differentiation in fracture healing: Mechanically loaded composite scaffolds in a novel bioreactor system. Tissue Eng. 2006, 12, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Matziolis, D.; Tuischer, J.; Matziolis, G.; Kasper, G.; Duda, G.; Perka, C. Osteogenic predifferentiation of human bone marrow-derived stem cells by short-term mechanical stimulation. Open Orthop. J. 2011, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ramani-Mohan, R.K.; Schwedhelm, I.; Finne-Wistrand, A.; Krug, M.; Schwarz, T.; Jakob, F.; Walles, H.; Hansmann, J. Deformation strain is the main physical driver for skeletal precursors to undergo osteogenesis in earlier stages of osteogenic cell maturation. J. Tissue Eng. Regen. Med. 2018, 12, e1474–e1479. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W.; Feliciano, S.; Martin, I.; de Wild, M.; Wendt, D. Novel perfused compression bioreactor system as an in vitro model to investigate fracture healing. Front. Bioeng. Biotechnol. 2015, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Joly, P.; Bergmann, C.; Korus, G.; Duda, G.N. The impact of substrate stiffness and mechanical loading on fibroblast-induced scaffold remodeling. Tissue Eng. Part A 2012, 18, 1804–1817. [Google Scholar] [CrossRef] [PubMed]
- Jagodzinski, M.; Breitbart, A.; Wehmeier, M.; Hesse, E.; Haasper, C.; Krettek, C.; Zeichen, J.; Hankemeier, S. Influence of perfusion and cyclic compression on proliferation and differentiation of bone marrow stromal cells in 3-dimensional culture. J. Biomech. 2008, 41, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Kopf, J.; Petersen, A.; Duda, G.N.; Knaus, P. Bmp2 and mechanical loading cooperatively regulate immediate early signalling events in the bmp pathway. BMC Biol. 2012, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.G.; Simpson, A.E.; Jaehn, K.; Furlong, P.I.; Stoddart, M.J. Establishing a 3D ex vivo culture system for investigations of bone metabolism and biomaterial interactions. ALTEX 2007, 24, 56–59. [Google Scholar] [PubMed]
- Jones, D.B.; Broeckmann, E.; Pohl, T.; Smith, E.L. Development of a mechanical testing and loading system for trabecular bone studies for long term culture. Eur. Cell Mater. 2003, 5, 48–59, discussion 59–60. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.; Cartmell, S.H.; Keramane, A.; El Haj, A.J. Principles and design of a novel magnetic force mechanical conditioning bioreactor for tissue engineering, stem cell conditioning and dynamic in vitro screening. IEEE Trans. Nanobiosci. 2006, 5, 173–177. [Google Scholar] [CrossRef]
- Cartmell, S.H.; Dobson, J.; Verschueren, S.B.; El Haj, A.J. Development of magnetic particle techniques for long-term culture of bone cells with intermittent mechanical activation. IEEE Trans. Nanobiosci. 2002, 1, 92–97. [Google Scholar] [CrossRef]
- Schulze, F.; Dienelt, A.; Geissler, S.; Zaslansky, P.; Schoon, J.; Henzler, K.; Guttmann, P.; Gramoun, A.; Crowe, L.A.; Maurizi, L.; et al. Amino-polyvinyl alcohol coated superparamagnetic iron oxide nanoparticles are suitable for monitoring of human mesenchymal stromal cells in vivo. Small 2014, 10, 4340–4351. [Google Scholar] [CrossRef] [PubMed]
- Kodzius, R.; Schulze, F.; Gao, X.; Schneider, M.R. Organ-on-chip technology: Current state and future developments. Genes (Basel) 2017, 8, 266. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Sackmann, E.K.; Fulton, A.L.; Beebe, D.J. The present and future role of microfluidics in biomedical research. Nature 2014, 507, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Selimovic, S.; Gauvin, R.; Bae, H. Organ-on-a-chip: Development and clinical prospects toward toxicity assessment with an emphasis on bone marrow. Drug Saf. 2015, 38, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Torisawa, Y.S.; Spina, C.S.; Mammoto, T.; Mammoto, A.; Weaver, J.C.; Tat, T.; Collins, J.J.; Ingber, D.E. Bone marrow-on-a-chip replicates hematopoietic niche physiology in vitro. Nat. Methods 2014, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Sieber, S.; Wirth, L.; Cavak, N.; Koenigsmark, M.; Marx, U.; Lauster, R.; Rosowski, M. Bone marrow-on-a-chip: Long-term culture of human haematopoietic stem cells in a three-dimensional microfluidic environment. J. Tissue Eng. Regen. Med. 2017, 12, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Jusoh, N.; Oh, S.; Kim, S.; Kim, J.; Jeon, N.L. Microfluidic vascularized bone tissue model with hydroxyapatite-incorporated extracellular matrix. Lab Chip 2015, 15, 3984–3988. [Google Scholar] [CrossRef] [PubMed]
- Bersini, S.; Gilardi, M.; Arrigoni, C.; Talo, G.; Zamai, M.; Zagra, L.; Caiolfa, V.; Moretti, M. Human in vitro 3D co-culture model to engineer vascularized bone-mimicking tissues combining computational tools and statistical experimental approach. Biomaterials 2016, 76, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.L.; Choudhary, S.; Mannion, C.; Kissin, Y.; Zilberberg, J.; Lee, W.Y. Ex vivo replication of phenotypic functions of osteocytes through biomimetic 3D bone tissue construction. Bone 2018, 106, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Sim, W.Y.; Min, B.H.; Yang, S.S.; Khademhosseini, A.; Kaplan, D.L. Chip-based comparison of the osteogenesis of human bone marrow- and adipose tissue-derived mesenchymal stem cells under mechanical stimulation. PLoS ONE 2012, 7, e46689. [Google Scholar] [CrossRef] [PubMed]
- Torisawa, Y.S.; Mammoto, T.; Jiang, E.; Jiang, A.; Mammoto, A.; Watters, A.L.; Bahinski, A.; Ingber, D.E. Modeling hematopoiesis and responses to radiation countermeasures in a bone marrow-on-a-chip. Tissue Eng. Part C Methods 2016, 22, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Walasek, M.A.; van Os, R.; de Haan, G. Hematopoietic stem cell expansion: Challenges and opportunities. Ann. N. Y. Acad. Sci. 2012, 1266, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Scotti, C.; Tonnarelli, B.; Papadimitropoulos, A.; Scherberich, A.; Schaeren, S.; Schauerte, A.; Lopez-Rios, J.; Zeller, R.; Barbero, A.; Martin, I. Recapitulation of endochondral bone formation using human adult mesenchymal stem cells as a paradigm for developmental engineering. Proc. Natl. Acad. Sci. USA 2010, 107, 7251–7256. [Google Scholar] [CrossRef] [PubMed]
- Kostenuik, P.; Mirza, F.M. Fracture healing physiology and the quest for therapies for delayed healing and nonunion. J. Orthop. Res. 2017, 35, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Rakow, A.; Schoon, J.; Dienelt, A.; John, T.; Textor, M.; Duda, G.; Perka, C.; Schulze, F.; Ode, A. Influence of particulate and dissociated metal-on-metal hip endoprosthesis wear on mesenchymal stromal cells in vivo and in vitro. Biomaterials 2016, 98, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Schoon, J.; Geissler, S.; Traeger, J.; Luch, A.; Tentschert, J.; Perino, G.; Schulze, F.; Duda, G.N.; Perka, C.; Rakow, A. Multi-elemental nanoparticle exposure after tantalum component failure in hip arthroplasty: In-depth analysis of a single case. Nanomedicine 2017, 13, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Blaber, E.; Marcal, H.; Burns, B.P. Bioastronautics: The influence of microgravity on astronaut health. Astrobiology 2010, 10, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.T. What do we know about the effects of spaceflight on bone? J. Appl. Physiol. 2000, 89, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Sibonga, J.D.; Spector, E.R.; Johnston, S.L.; Tarver, W.J. Evaluating bone loss in iss astronauts. Aerosp. Med. Hum. Perf. 2015, 86, A38–A44. [Google Scholar] [CrossRef] [PubMed]
- Orwoll, E.S.; Adler, R.A.; Amin, S.; Binkley, N.; Lewiecki, E.M.; Petak, S.M.; Shapses, S.A.; Sinaki, M.; Watts, N.B.; Sibonga, J.D. Skeletal health in long-duration astronauts: Nature, assessment and management recommendations from the nasa bone summit. J. Bone Miner. Res. 2013, 28, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, P.R.; Licata, A.A.; Rice, A.J. Exercise and pharmacological countermeasures for bone loss during long-duration space flight. Gravit. Space Biol. Bull. 2005, 18, 39–58. [Google Scholar] [PubMed]
- NASA. Osteocytes and Mechano-Transduction (osteo-4). Available online: https://www.nasa.gov/mission_pages/station/research/experiments/1276.html (accessed on 30 January 2018).
- Bouet, G.; Cruel, M.; Laurent, C.; Vico, L.; Malaval, L.; Marchat, D. Validation of an in vitro 3D bone culture model with perfused and mechanically stressed ceramic scaffold. Eur. Cell Mater. 2015, 29, 250–266, discussion 266–257. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Haniu, H.; Kamanaka, T.; Takizawa, T.; Sobajima, A.; Yoshida, K.; Aoki, K.; Okamoto, M.; Kato, H.; Saito, N. Physico-chemical, in vitro, and in vivo evaluation of a 3D unidirectional porous hydroxyapatite scaffold for bone regeneration. Materials (Basel) 2017, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Baykan, E.; Koc, A.; Eser Elcin, A.; Murat Elcin, Y. Evaluation of a biomimetic poly(ε-caprolactone)/β-tricalcium phosphate multispiral scaffold for bone tissue engineering: In vitro and in vivo studies. Biointerphases 2014, 9, 029011. [Google Scholar] [CrossRef] [PubMed]
- Berner, A.; Henkel, J.; Woodruff, M.A.; Saifzadeh, S.; Kirby, G.; Zaiss, S.; Gohlke, J.; Reichert, J.C.; Nerlich, M.; Schuetz, M.A.; et al. Scaffold-cell bone engineering in a validated preclinical animal model: Precursors vs differentiated cell source. J. Tissue Eng. Regen. Med. 2017, 11, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, D.; Kiamehr, M.; Yang, X.; Su, B. Fabrication and in vitro evaluation of a sponge-like bioactive-glass/gelatin composite scaffold for bone tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2013, 33, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Cao, H.; Wang, X.; Chen, S.; Zhang, M.; Wang, N.; Yao, Z.; Dai, Y.; Xie, X.; Zhang, P.; et al. Porous composite scaffold incorporating osteogenic phytomolecule icariin for promoting skeletal regeneration in challenging osteonecrotic bone in rabbits. Biomaterials 2018, 153, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dhivya, S.; Keshav Narayan, A.; Logith Kumar, R.; Viji Chandran, S.; Vairamani, M.; Selvamurugan, N. Proliferation and differentiation of mesenchymal stem cells on scaffolds containing chitosan, calcium polyphosphate and pigeonite for bone tissue engineering. Cell Prolif. 2017. [Google Scholar] [CrossRef] [PubMed]
- Barba, A.; Diez-Escudero, A.; Maazouz, Y.; Rappe, K.; Espanol, M.; Montufar, E.B.; Bonany, M.; Sadowska, J.M.; Guillem-Marti, J.; Ohman-Magi, C.; et al. Osteoinduction by foamed and 3D-printed calcium phosphate scaffolds: Effect of nanostructure and pore architecture. ACS Appl. Mater. Interfaces 2017, 9, 41722–41736. [Google Scholar] [CrossRef] [PubMed]
- Burdette, A.J.; Guda, T.; Thompson, M.E.; Banas, R.; Sheppard, F. A novel secretome biotherapeutic influences regeneration in critical size bone defects. J. Craniofac. Surg. 2018, 29, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhang, N.; Marsano, A.; Vunjak-Novakovic, G.; Zhang, Y.; Lopez, M.J. In vitro mesenchymal trilineage differentiation and extracellular matrix production by adipose and bone marrow derived adult equine multipotent stromal cells on a collagen scaffold. Stem Cell Rev. 2013, 9, 858–872. [Google Scholar] [CrossRef] [PubMed]
- Dinescu, S.; Ionita, M.; Pandele, A.M.; Galateanu, B.; Iovu, H.; Ardelean, A.; Costache, M.; Hermenean, A. In vitro cytocompatibility evaluation of chitosan/graphene oxide 3D scaffold composites designed for bone tissue engineering. Biomed. Mater. Eng. 2014, 24, 2249–2256. [Google Scholar] [PubMed]
- Li, Q.; Yu, X.; Zhou, G.; Wu, Y.W.; Hu, H.C.; Wang, T.; Tang, Z.H. [synthesis and in vitro characterization of chitosan microspheres/ceramic bovine bone composite scaffold]. Beijing Da Xue Xue Bao 2016, 48, 1043–1048. [Google Scholar] [PubMed]
- Oryan, A.; Alidadi, S.; Bigham-Sadegh, A.; Moshiri, A. Effectiveness of tissue engineered based platelet gel embedded chitosan scaffold on experimentally induced critical sized segmental bone defect model in rat. Injury 2017, 48, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, Y.; Tian, X.; Cui, G.; Zhao, Y.; Yang, Q.; Yu, S.; Xing, G.; Zhang, B. Segmental bone regeneration using an rhBMP-2-loaded gelatin/nanohydroxyapatite/fibrin scaffold in a rabbit model. Biomaterials 2009, 30, 6276–6285. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Lin, Z.; Chen, J.; Fan, Y.; Davey, T.; Cake, M.; Day, R.; Dai, K.; Xu, J.; Zheng, M. Natural bone collagen scaffold combined with autologous enriched bone marrow cells for induction of osteogenesis in an ovine spinal fusion model. Tissue Eng. Part A 2009, 15, 3547–3558. [Google Scholar] [CrossRef] [PubMed]
- Gholipourmalekabadi, M.; Mozafari, M.; Bandehpour, M.; Salehi, M.; Sameni, M.; Caicedo, H.H.; Mehdipour, A.; Hamidabadi, H.G.; Samadikuchaksaraei, A.; Ghanbarian, H. Optimization of nanofibrous silk fibroin scaffold as a delivery system for bone marrow adherent cells: In vitro and in vivo studies. Biotechnol. Appl. Biochem. 2015, 62, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.L.; Sánchez-Abarca, L.I.; Muntión, S.; Preciado, S.; Puig, N.; López-Ruano, G.; Hernández-Hernández, Á.; Redondo, A.; Ortega, R.; Rodríguez, C.; et al. MSC surface markers (CD44, CD73, and CD90) can identify human msc-derived extracellular vesicles by conventional flow cytometry. J. Cell Commun. Signal. 2016, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, S.; Cakmak, A.S.; Gumusderelioglu, M. RGD-bearing peptide-amphiphile-hydroxyapatite nanocomposite bone scaffold: an in vitro study. Biomed. Mater. 2013, 8, 045014. [Google Scholar] [CrossRef] [PubMed]
- Koc, A.; Finkenzeller, G.; Elcin, A.E.; Stark, G.B.; Elcin, Y.M. Evaluation of adenoviral vascular endothelial growth factor-activated chitosan/hydroxyapatite scaffold for engineering vascularized bone tissue using human osteoblasts: In vitro and in vivo studies. J. Biomater. Appl. 2014, 29, 748–760. [Google Scholar] [CrossRef] [PubMed]
- Przekora, A.; Ginalska, G. In vitro evaluation of the risk of inflammatory response after chitosan/HA and chitosan/β-1,3-glucan/HA bone scaffold implantation. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 61, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Przekora, A.; Vandrovcova, M.; Travnickova, M.; Pajorova, J.; Molitor, M.; Ginalska, G.; Bacakova, L. Evaluation of the potential of chitosan/β-1,3-glucan/hydroxyapatite material as a scaffold for living bone graft production in vitro by comparison of ADSC and BMDSC behaviour on its surface. Biomed. Mater. 2017, 12, 015030. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Yuan, F.; Zhimin, Z.; Anchun, M. Dynamic perfusion bioreactor system for 3D culture of rat bone marrow mesenchymal stem cells on nanohydroxyapatite/polyamide 66 scaffold in vitro. J. Biomed. Mater. Res. Part B Appl. Biomater. 2013, 101, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Xu, D.P.; Liu, Z.M.; Du, Y.; Wang, X.K. Synthesis of and in vitro and in vivo evaluation of a novel TGF-β1-SF-CS three-dimensional scaffold for bone tissue engineering. Int. J. Mol. Med. 2016, 38, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Yoon, D.; Kang, B.J.; Kim, Y.; Lee, S.H.; Rhew, D.; Kim, W.H.; Kweon, O.K. Effect of serum-derived albumin scaffold and canine adipose tissue-derived mesenchymal stem cells on osteogenesis in canine segmental bone defect model. J. Vet. Sci. 2015, 16, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Gerlach, J.C.; Schmelzer, E.; Nettleship, I. Effect of calcium-infiltrated hydroxyapatite scaffolds on the hematopoietic fate of human umbilical vein endothelial cells. J. Vasc. Res. 2017, 54, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Bae, J.H.; Park, S.A.; Kim, W.D.; Park, M.S.; Ko, Y.J.; Jang, H.S.; Park, J.H. Combination therapy with BMP-2 and BMSCS enhances bone healing efficacy of PCL scaffold fabricated using the 3D plotting system in a large segmental defect model. Biotechnol. Lett. 2012, 34, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Rahman, C.V.; Ben-David, D.; Dhillon, A.; Kuhn, G.; Gould, T.W.; Muller, R.; Rose, F.R.; Shakesheff, K.M.; Livne, E. Controlled release of BMP-2 from a sintered polymer scaffold enhances bone repair in a mouse calvarial defect model. J. Tissue Eng. Regen. Med. 2014, 8, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Requicha, J.F.; Moura, T.; Leonor, I.B.; Martins, T.; Munoz, F.; Reis, R.L.; Gomes, M.E.; Viegas, C.A. Evaluation of a starch-based double layer scaffold for bone regeneration in a rat model. J. Orthop. Res. 2014, 32, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Wang, K.; Shimer, A.L.; Li, X.; Balian, G.; Shen, F.H. Use of a bioactive scaffold for the repair of bone defects in a novel reproducible vertebral body defect model. Bone 2010, 47, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, X.; Zeng, X.; Zhu, J.; Pi, Y.; Zhou, C.; Ao, Y. One-step articular cartilage repair: Combination of in situ bone marrow stem cells with cell-free Poly(l-lactic-co-glycolic acid) scaffold in a rabbit model. Orthopedics 2012, 35, e665–671. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Maiti, T.K.; Bhattacharya, D.; Nandi, S.K. Effect of different mineralization processes on in vitro and in vivo bone regeneration and osteoblast-macrophage cross-talk in co-culture system using dual growth factor mediated non-mulberry silk fibroin grafted poly (capital je, ukrainian-caprolactone) nanofibrous scaffold. Colloids Surf. B Biointerfaces 2017, 156, 270–281. [Google Scholar] [PubMed]
- Suliman, S.; Xing, Z.; Wu, X.; Xue, Y.; Pedersen, T.O.; Sun, Y.; Doskeland, A.P.; Nickel, J.; Waag, T.; Lygre, H.; et al. Release and bioactivity of bone morphogenetic protein-2 are affected by scaffold binding techniques in vitro and in vivo. J. Control. Release 2015, 197, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Vergroesen, P.P.; Kroeze, R.J.; Helder, M.N.; Smit, T.H. The use of Poly(l-lactide-co-caprolactone) as a scaffold for adipose stem cells in bone tissue engineering: Application in a spinal fusion model. Macromol. Biosci. 2011, 11, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Giavaresi, G.; Guarino, V.; Raucci, M.G.; Sandri, M.; Tampieri, A.; Ambrosio, L.; Fini, M. Bioactivity and bone healing properties of biomimetic porous composite scaffold: In vitro and in vivo studies. J. Biomed. Mater. Res. A 2015, 103, 2932–2941. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.; Tang, H.; Wu, B.; Ding, X.; Lu, Z.; Li, W.; Xu, Z. A specific groove design for individualized healing in a canine partial sternal defect model by a polycaprolactone/hydroxyapatite scaffold coated with bone marrow stromal cells. J. Biomed. Mater. Res. A 2014, 102, 3401–3408. [Google Scholar] [CrossRef] [PubMed]
- Lasher, R.A.; Wolchok, J.C.; Parikh, M.K.; Kennedy, J.P.; Hitchcock, R.W. Design and characterization of a modified T-flask bioreactor for continuous monitoring of engineered tissue stiffness. Biotechnol. Prog. 2010, 26, 857–864. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Polymer Origin | Advantages | Disadvantages |
|---|---|---|
| Natural | Biocompatibility and their degradation is facilitated by enzymes present in vivo | Inconsistent hydration and elastic properties |
| Synthetic | Improved consistency and ability to modify properties (degradation, cell binding) | Weak mechanical strength and inability to sequester growth factors, resulting in burst release |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheinpflug, J.; Pfeiffenberger, M.; Damerau, A.; Schwarz, F.; Textor, M.; Lang, A.; Schulze, F. Journey into Bone Models: A Review. Genes 2018, 9, 247. https://doi.org/10.3390/genes9050247
Scheinpflug J, Pfeiffenberger M, Damerau A, Schwarz F, Textor M, Lang A, Schulze F. Journey into Bone Models: A Review. Genes. 2018; 9(5):247. https://doi.org/10.3390/genes9050247
Chicago/Turabian StyleScheinpflug, Julia, Moritz Pfeiffenberger, Alexandra Damerau, Franziska Schwarz, Martin Textor, Annemarie Lang, and Frank Schulze. 2018. "Journey into Bone Models: A Review" Genes 9, no. 5: 247. https://doi.org/10.3390/genes9050247
APA StyleScheinpflug, J., Pfeiffenberger, M., Damerau, A., Schwarz, F., Textor, M., Lang, A., & Schulze, F. (2018). Journey into Bone Models: A Review. Genes, 9(5), 247. https://doi.org/10.3390/genes9050247