A-to-I RNA Editing Affects lncRNAs Expression after Heat Shock


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
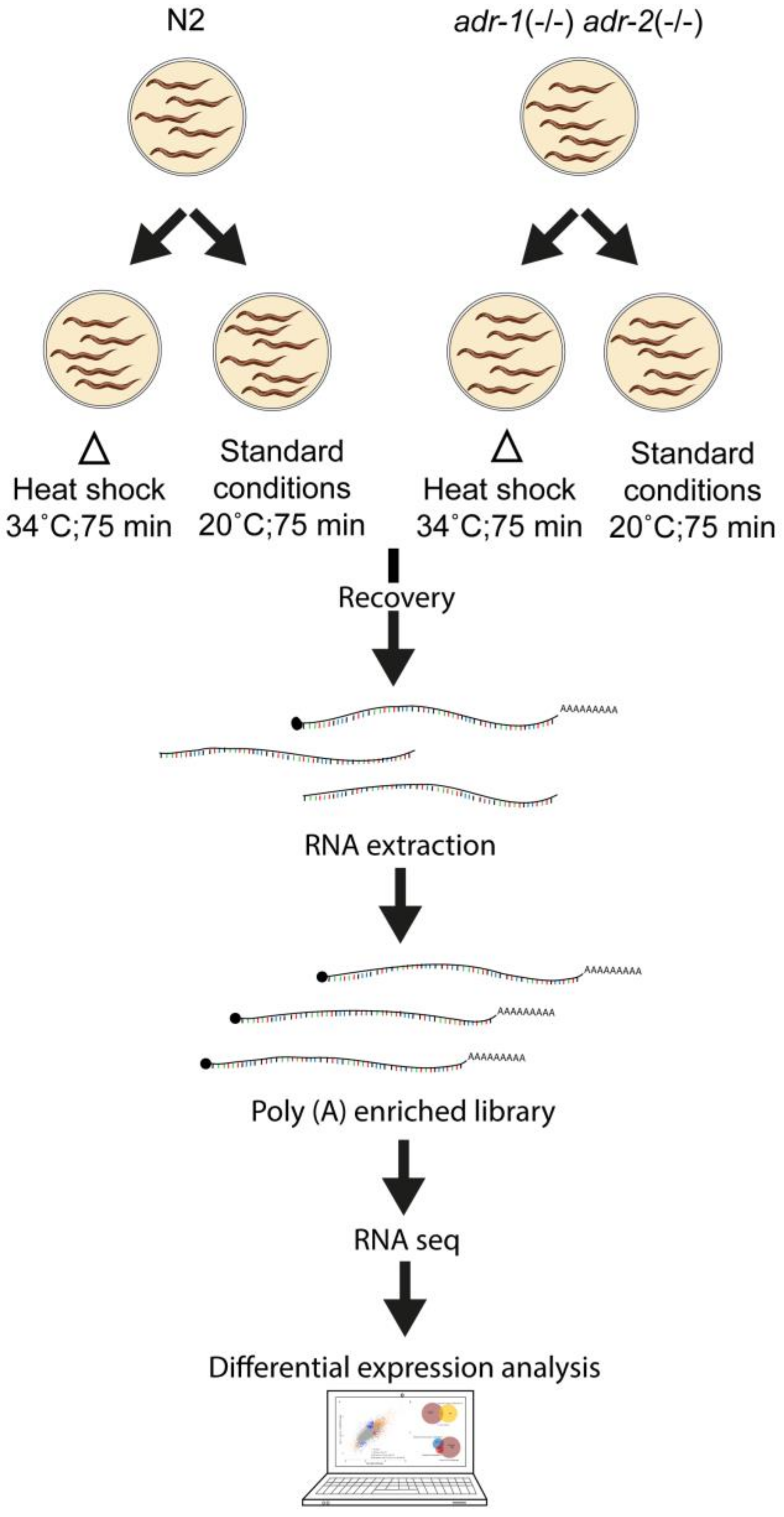
2. Materials and Methods
2.1. Maintenance and Growth of Caenorhabditis elegans Strains
2.2. Heat Shock Treatment
2.3. RNA Extraction and mRNA Library Preparation
2.4. RNA-Seq and Gene Expression Analysis
2.5. Data Availability
3. Results
3.1. Gene Expression Under Standard Conditions and upon Heat-Shock
3.2. Expression of Heat-Shock Genes upon Heat Stress Is Not Affected by Adenosine-Deaminases (ADAR) Function
3.3. Pseudogenes and lncRNAs Are Differentially Expressed upon Heat Shock in Adenosine-Deaminases (ADAR) Mutants
3.4. No Substantial Enrichment of Gene OntologyTerms in Adenosine-Deaminases (ADAR) Specifically Expressed Genes after Heat Shock
4. Discussion
4.1. Adenosine to Inosine RNA Editing Does Not Directly Affect Heat Shock Related Genes upon Heat Stress
4.2. Adenosine to Inosine RNA Editing Regulates the Heat Stress Response
4.3. Elevating Temperatures May Destabilize dsRNA Structures and Protect Them from RNAi Degradation, Despite the Lack of Adenosine-Deaminases (ADAR) Function
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morimoto, R.I. Regulation of the heat shock transcriptional response: Cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev. 1998, 12, 3788–3796. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, A.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Prahlad, V.; Cornelius, T.; Morimoto, R.I. Regulation of the cellular heat shock response in Caenorhabditis elegans by thermosensory neurons. Science 2008, 320, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Buchumenski, I.; Bartok, O.; Ashwal-Fluss, R.; Pandey, V.; Porath, H.T.; Levanon, E.Y.; Kadener, S. Dynamic hyper-editing underlies temperature adaptation in Drosophila. PLOS Genet. 2017, 13, e1006931. [Google Scholar] [CrossRef] [PubMed]
- Garrett, S.C.; Rosenthal, J.J.C. A Role for A-to-I RNA editing in temperature adaptation. Physiology 2012, 27, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Savva, Y.A.; Rieder, L.E.; Reenan, R.A. The ADAR protein family. Genome Boil. 2012, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat. Rev. Mol. Cell Boil. 2015, 17, 83. [Google Scholar] [CrossRef]
- Bazak, L.; Haviv, A.; Barak, M.; Jacob-Hirsch, J.; Deng, P.; Zhang, R.; Isaacs, F.J.; Rechavi, G.; Li, J.B.; Eisenberg, E.; et al. A-to-I RNA editing occurs at over a hundred million genomic sites, located in a majority of human genes. Genome Res. 2014, 24, 365–376. [Google Scholar] [CrossRef]
- Garrett, S.; Rosenthal, J.J. RNA editing underlies temperature adaptation in K+ channels from polar octopuses. Science 2012, 335, 848–851. [Google Scholar] [CrossRef]
- Higuchi, M.; Maas, S.; Single, F.N.; Hartner, J.; Rozov, A.; Burnashev, N.; Feldmeyer, D.; Sprengel, R.; Seeburg, P.H. Point mutation in an AMPA receptor gene rescues lethality in mice deficient in the RNA-editing enzyme ADAR2. Nature 2000, 406, 78–81. [Google Scholar] [CrossRef]
- Rueter, S.M.; Dawson, T.R.; Emeson, R.B. Regulation of alternative splicing by RNA editing. Nature 1999, 399, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Schmauss, C.; Howe, J.R. RNA editing of neurotransmitter receptors in the mammalian brain. Sci. STKE 2002, 2002. [Google Scholar] [CrossRef]
- Hoopengardner, B.; Bhalla, T.; Staber, C.; Reenan, R. Nervous system targets of RNA editing identified by comparative genomics. Science 2003, 301, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Liddicoat, B.J.; Piskol, R.; Chalk, A.M.; Ramaswami, G.; Higuchi, M.; Hartner, J.C.; Li, J.B.; Seeburg, P.H.; Walkley, C.R. RNA editing by ADAR1 prevents MDA5 sensing of endogenous dsRNA as nonself. Science 2015, 349, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Ganem, N.S.; Lamm, A.T. A-to-I RNA editing—Thinking beyond the single nucleotide. RNA Biol. 2017, 14, 1690–1694. [Google Scholar] [CrossRef] [PubMed]
- Reich, D.P.; Tyc, K.M.; Bass, B.L.C. C. elegans ADARs antagonize silencing of cellular dsRNAs by the antiviral RNAi pathway. Genes Dev. 2018, 32, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.; Agranat-Tamir, L.; Light, D.; Ben-Naim Zgayer, O.; Fishman, A.; Lamm, A.T. A-to-I RNA editing promotes developmental stage-specific gene and lncRNA expression. Genome Res. 2017, 27, 462–470. [Google Scholar] [CrossRef]
- Wu, D.; Lamm, A.T.; Fire, A.Z. Competition between ADAR and RNAi pathways for an extensive class of RNA targets. Nat. Struct. Mol. Biol. 2011, 18, 1094. [Google Scholar] [CrossRef]
- Zhao, H.Q.; Zhang, P.; Gao, H.; He, X.; Dou, Y.; Huang, A.Y.; Liu, X.M.; Ye, A.Y.; Dong, M.Q.; Wei, L. Profiling the RNA editomes of wild-type C. elegans and ADAR mutants. Genome Res. 2015, 25, 66–75. [Google Scholar] [CrossRef]
- Whipple, J.M.; Youssef, O.A.; Aruscavage, P.J.; Nix, D.A.; Hong, C.; Johnson, W.E.; Bass, B.L. Genome-wide profiling of the C. elegans dsRNAome. RNA 2015, 21, 786–800. [Google Scholar] [CrossRef]
- Sebastiani, P.; Montano, M.; Puca, A.; Solovieff, N.; Kojima, T.; Wang, M.C.; Melista, E.; Meltzer, M.; Fischer, S.E.; Andersen, S.; et al. RNA editing genes associated with extreme old age in humans and with lifespan in C. elegans. PLoS ONE 2009, 4, e8210. [Google Scholar] [CrossRef]
- Tonkin, L.A.; Saccomanno, L.; Morse, D.P.; Brodigan, T.; Krause, M.; Bass, B.L. RNA editing by ADARs is important for normal behavior in Caenorhabditis elegans. EMBO J. 2002, 21, 6025–6035. [Google Scholar] [CrossRef] [PubMed]
- Savva, Y.A.; Jepson, J.E.C.; Sahin, A.; Sugden, A.U.; Dorsky, J.S.; Alpert, L.; Lawrence, C.; Reenan, R.A. Auto-regulatory RNA editing fine-tunes mRNA re-coding and complex behaviour in Drosophila. Nat. Commun. 2012, 3, 790. [Google Scholar] [CrossRef] [PubMed]
- Stocker, J.; Huang, H.W.; Wang, H.M.; Chang, H.W.; Chiu, C.C.; Cho, C.L.; Tseng, C.N. Reduction of RNA A-to-I editing in Drosophila acclimated to heat shock. Kaohsiung J. Med. Sci. 2013, 29, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Rieder, L.E.; Savva, Y.A.; Reyna, M.A.; Chang, Y.J.; Dorsky, J.S.; Rezaei, A.; Reenan, R.A. Dynamic response of RNA editing to temperature in Drosophila. BMC Biol. 2015, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Palladino, M.J.; Keegan, L.P.; O’Connell, M.A.; Reenan, R.A. A-to-I Pre-mRNA Editing in Drosophila is primarily involved in adult nervous system function and integrity. Cell 2000, 102, 437–449. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Hundley, H.A.; Krauchuk, A.A.; Bass, B.L.C. C. elegans and H. sapiens mRNAs with edited 3’ UTRs are present on polysomes. RNA 2008, 14, 2050–2060. [Google Scholar] [CrossRef]
- Koudande, O.D.; Iraqi, F.; Thomson, P.C.; Teale, A.J.; van Arendonk, J.A. Strategies to optimize marker-assisted introgression of multiple unlinked QTL. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2000, 11, 145–150. [Google Scholar] [CrossRef]
- WormBase. Available online: http://www.wormbase.org (accessed on 29 October 2017).
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Brunquell, J.; Morris, S.; Lu, Y.; Cheng, F.; Westerheide, S.D. The genome-wide role of HSF-1 in the regulation of gene expression in Caenorhabditis elegans. BMC Genom. 2016, 17, 559. [Google Scholar] [CrossRef] [PubMed]
- Eden, E.; Lipson, D.; Yogev, S.; Yakhini, Z. Discovering motifs in ranked lists of DNA sequences. PLoS Comput. Biol. 2007, 3, e39. [Google Scholar] [CrossRef] [PubMed]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Zevian, S.C.; Yanowitz, J.L. Methodological considerations for heat shock of the nematode Caenorhabditis elegans. Methods 2014, 68, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.T.; Tanaka, Y.; Kodama, T.S.; Kyogoku, Y.; Yanagi, H.; Yura, T. Translational induction of heat shock transcription factor sigma32: Evidence for a built-in RNA thermosensor. Genes Dev. 1999, 13, 655–665. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Rich, A.; Maas, S. Widespread A-to-I RNA editing of Alu-containing mRNAs in the human transcriptome. PLoS Biol. 2004, 2, e391. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haas, R.; Ganem, N.S.; Keshet, A.; Orlov, A.; Fishman, A.; Lamm, A.T. A-to-I RNA Editing Affects lncRNAs Expression after Heat Shock. Genes 2018, 9, 627. https://doi.org/10.3390/genes9120627
Haas R, Ganem NS, Keshet A, Orlov A, Fishman A, Lamm AT. A-to-I RNA Editing Affects lncRNAs Expression after Heat Shock. Genes. 2018; 9(12):627. https://doi.org/10.3390/genes9120627
Chicago/Turabian StyleHaas, Roni, Nabeel S. Ganem, Ayya Keshet, Angela Orlov, Alla Fishman, and Ayelet T. Lamm. 2018. "A-to-I RNA Editing Affects lncRNAs Expression after Heat Shock" Genes 9, no. 12: 627. https://doi.org/10.3390/genes9120627
APA StyleHaas, R., Ganem, N. S., Keshet, A., Orlov, A., Fishman, A., & Lamm, A. T. (2018). A-to-I RNA Editing Affects lncRNAs Expression after Heat Shock. Genes, 9(12), 627. https://doi.org/10.3390/genes9120627