The Alteration of Nasopharyngeal and Oropharyngeal Microbiota in Children with MPP and Non-MPP

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Diagnosis, Sampling and Pathogen Examination
2.3. DNA Extraction, Library Construction, and Sequencing
2.4. Bioinformatics Analysis and Visualization
3. Results
3.1. Sample Information, Data Output, and Confounder Analysis
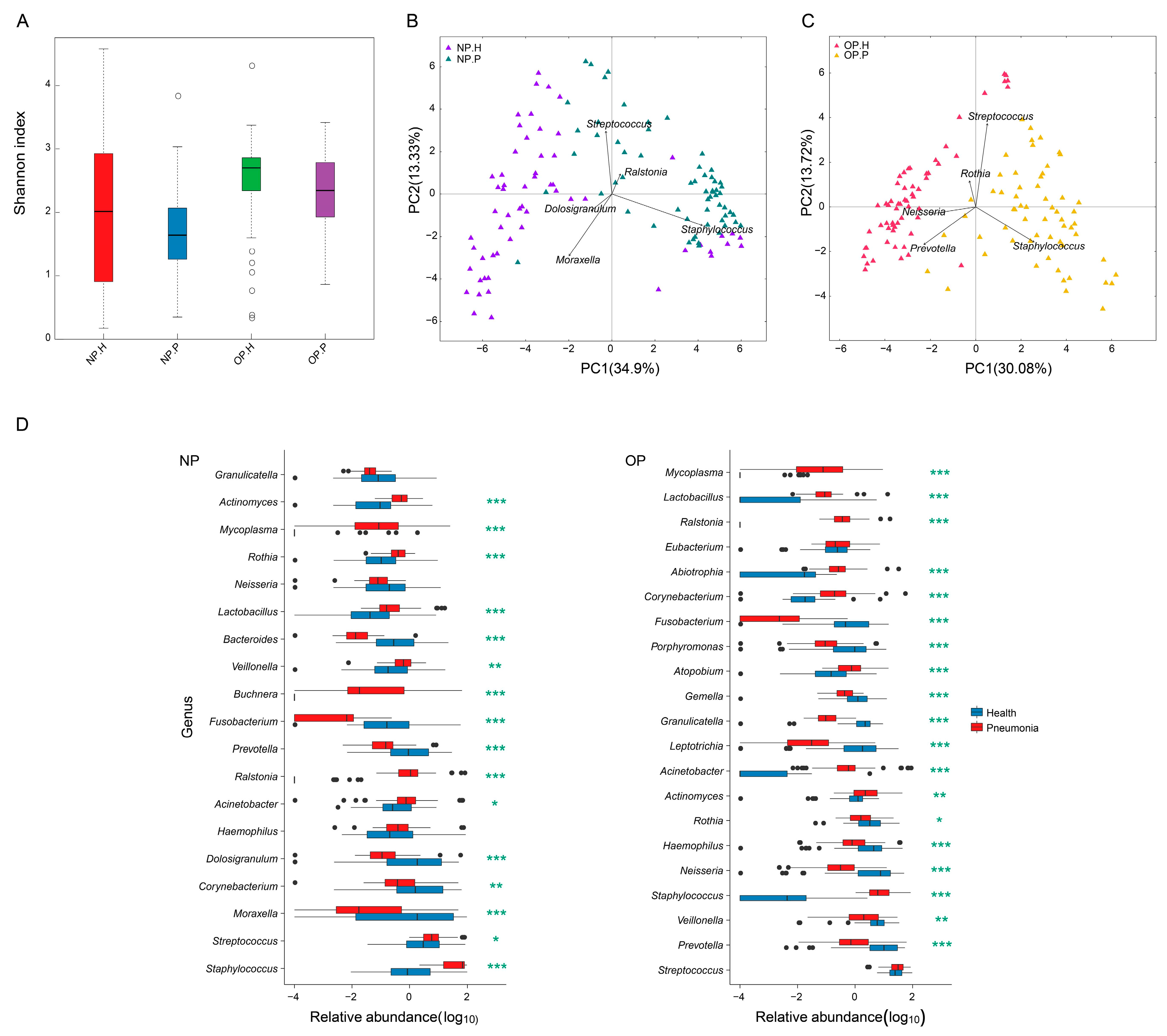
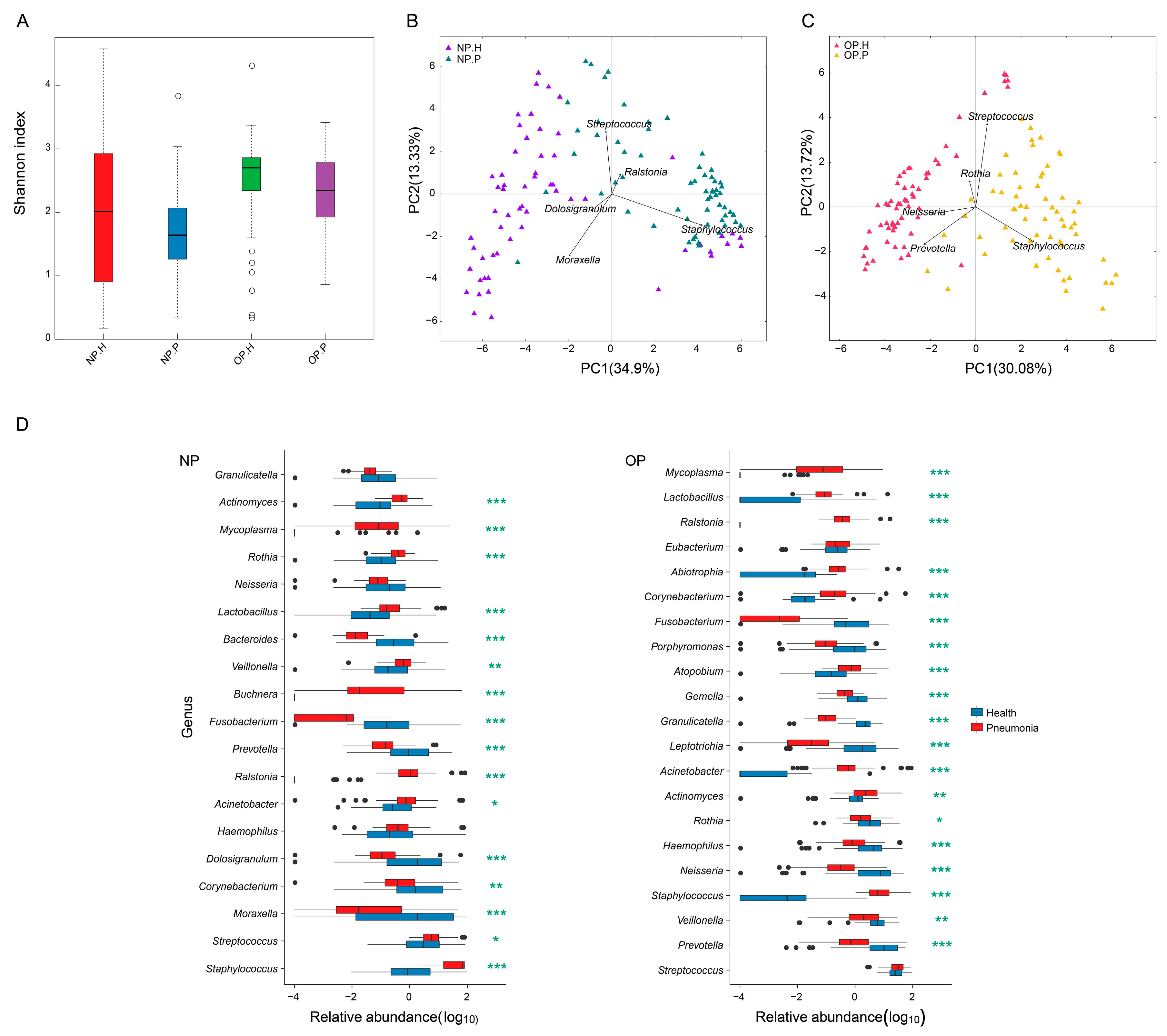
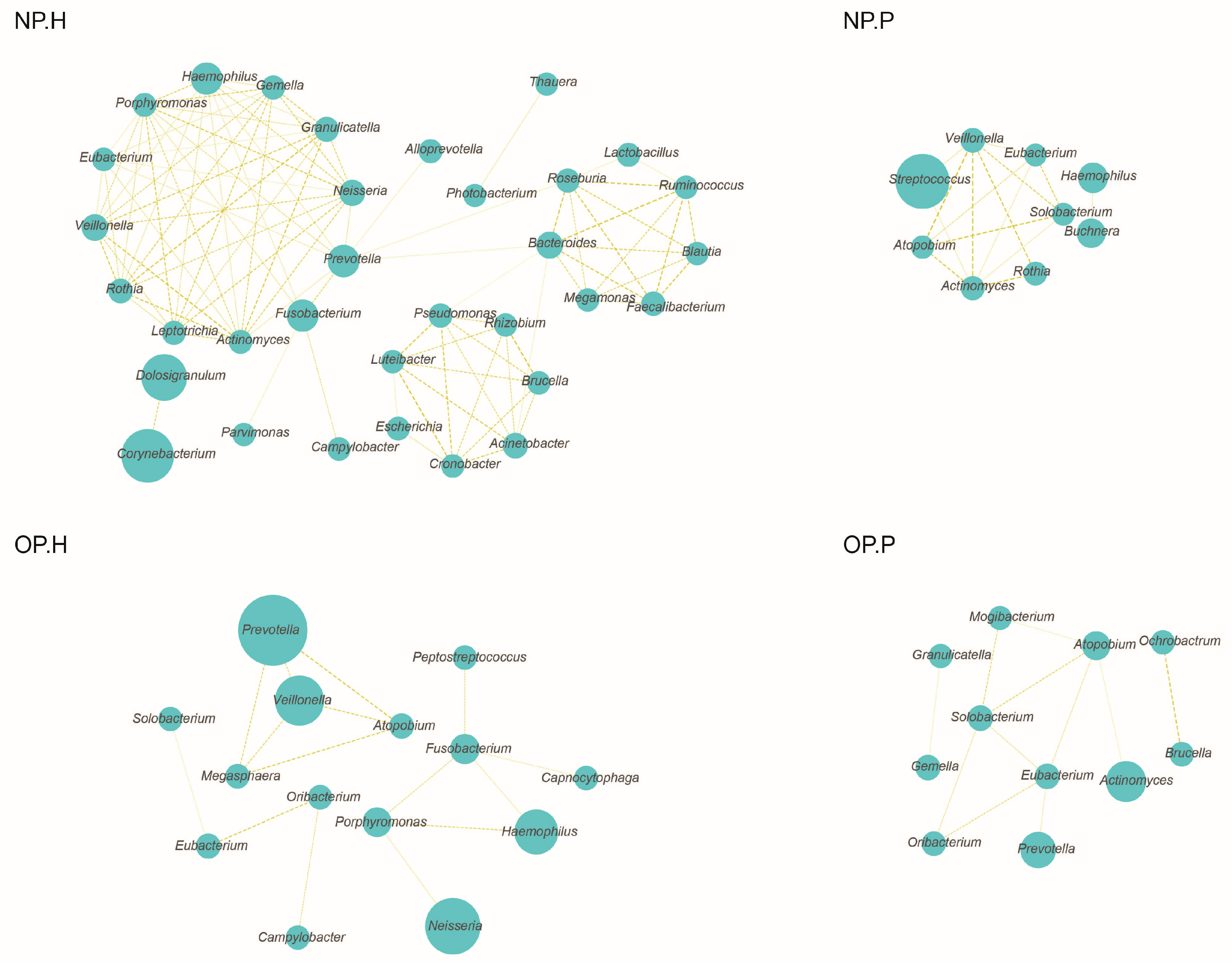
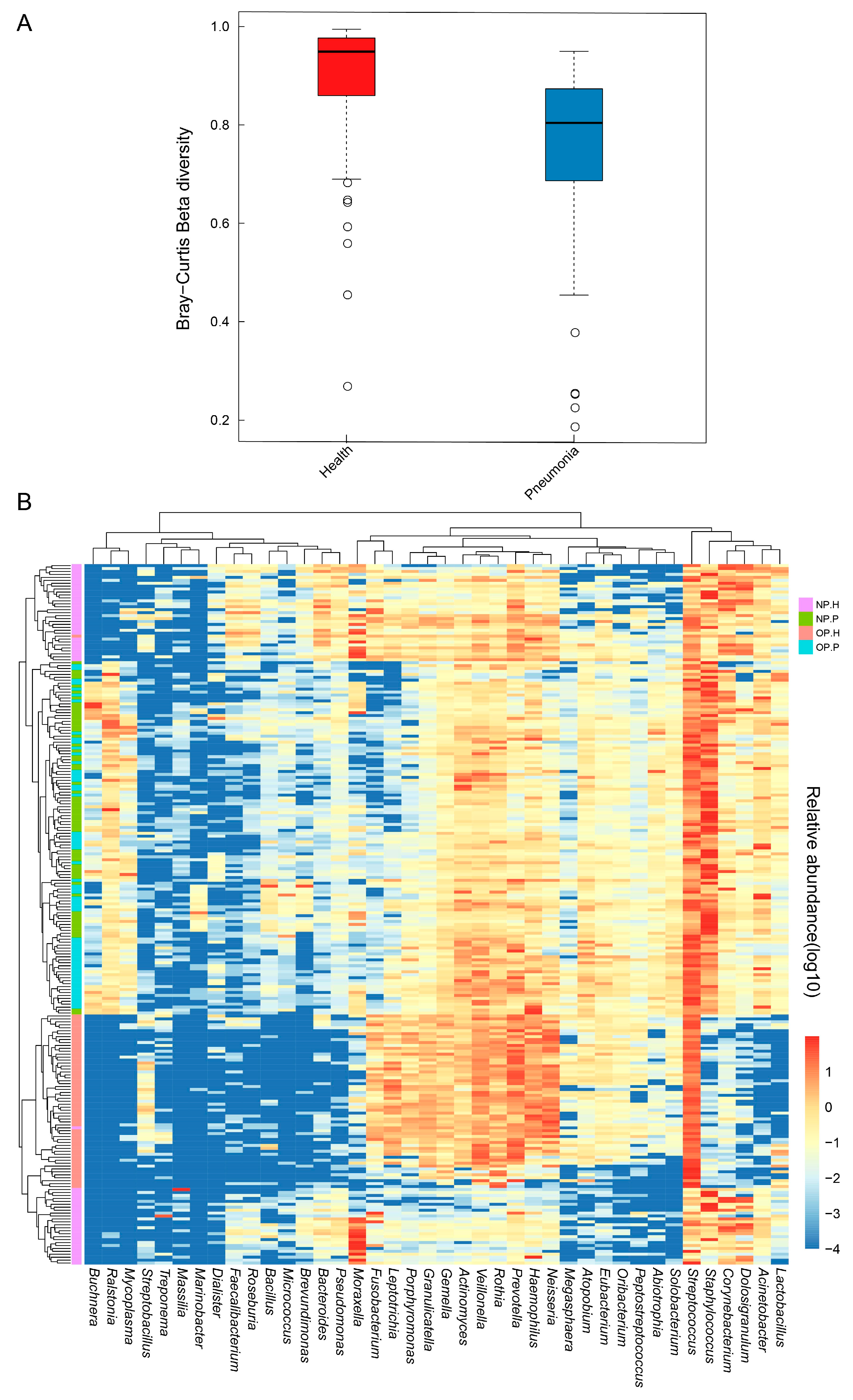
3.2. OP and NP Microbiota of Patients Differ Significantly from that of Healthy Children
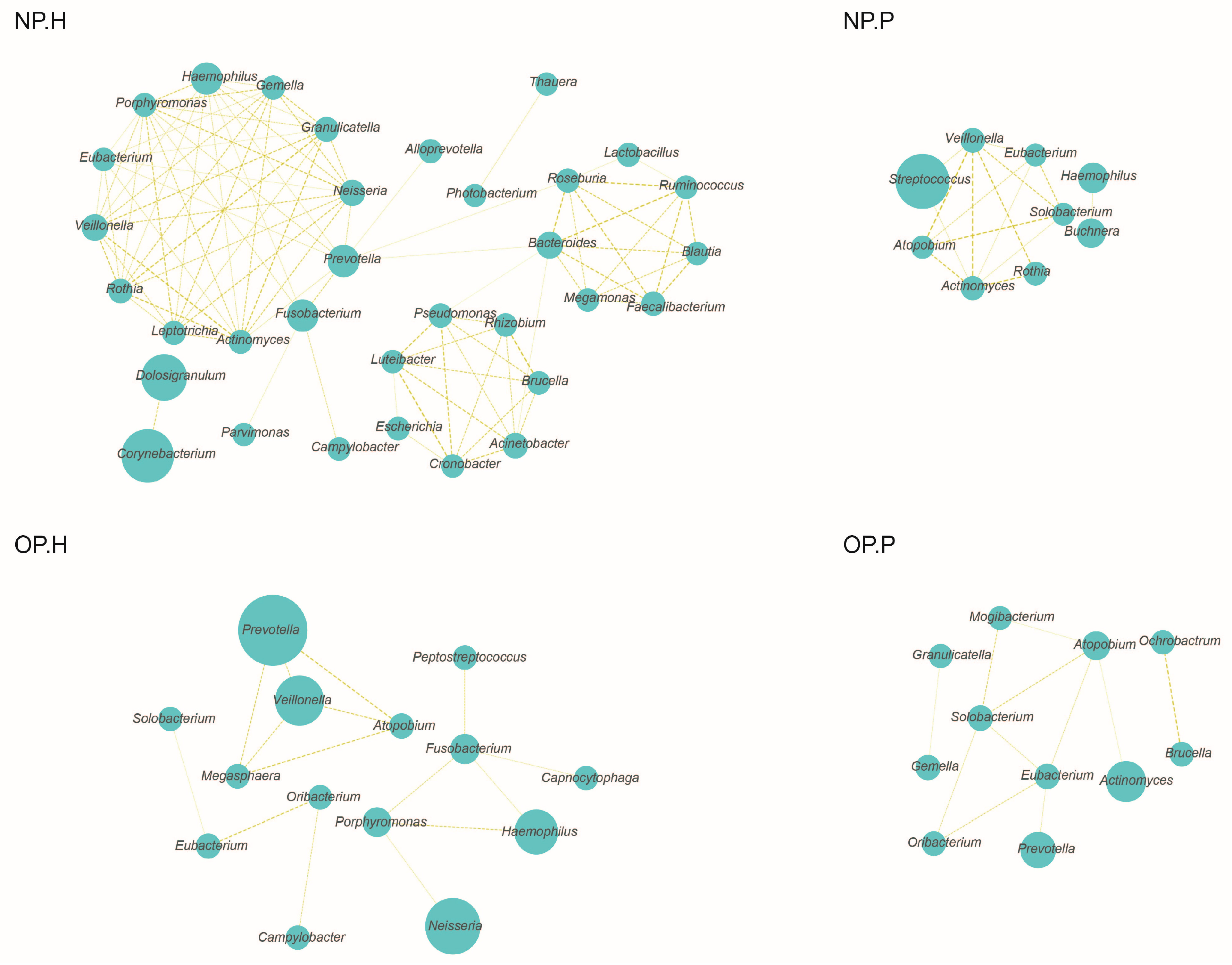
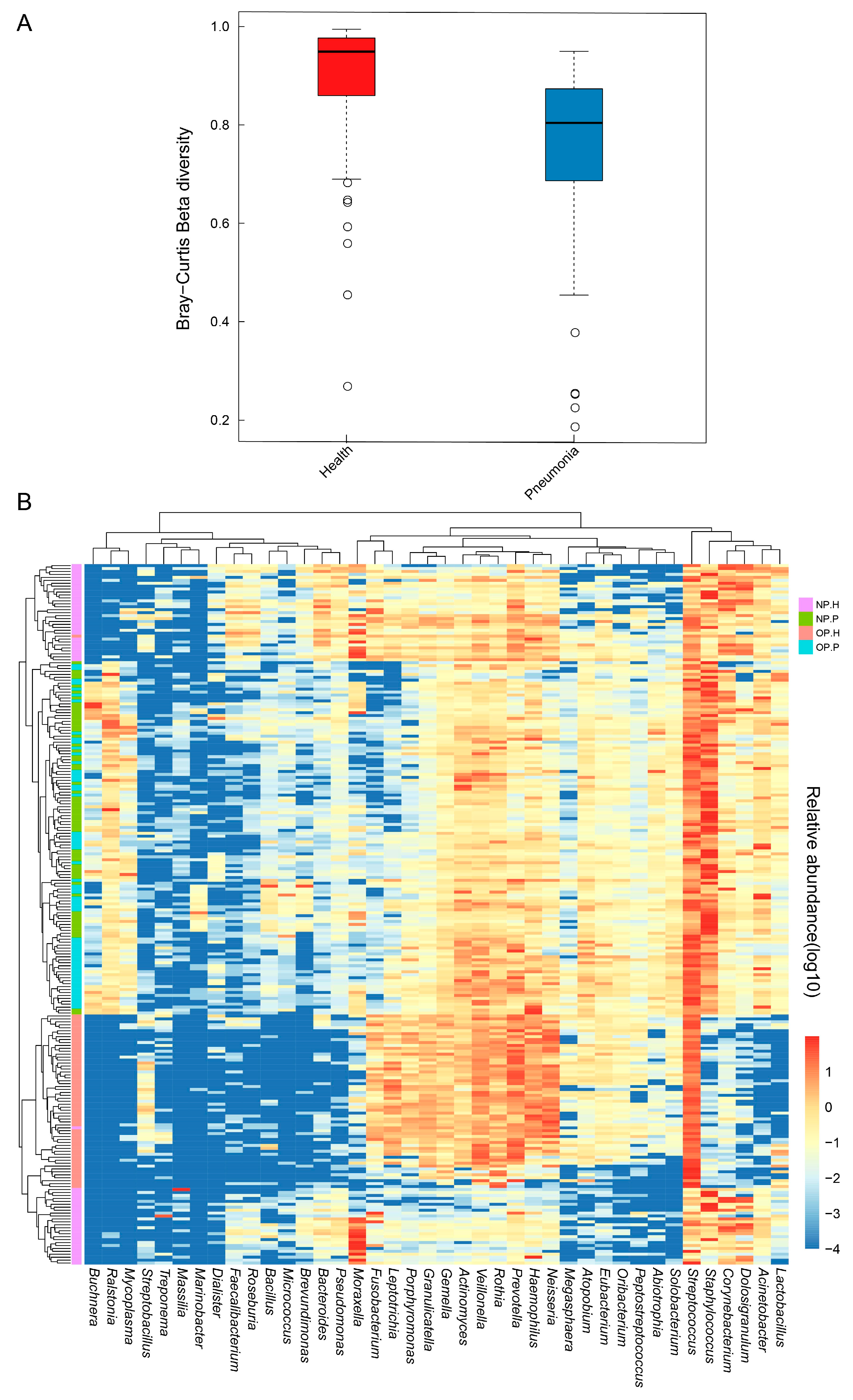
3.3. NP and OP Microbiota Pattern of Inpatients Have an Increased Similarity to Each Other
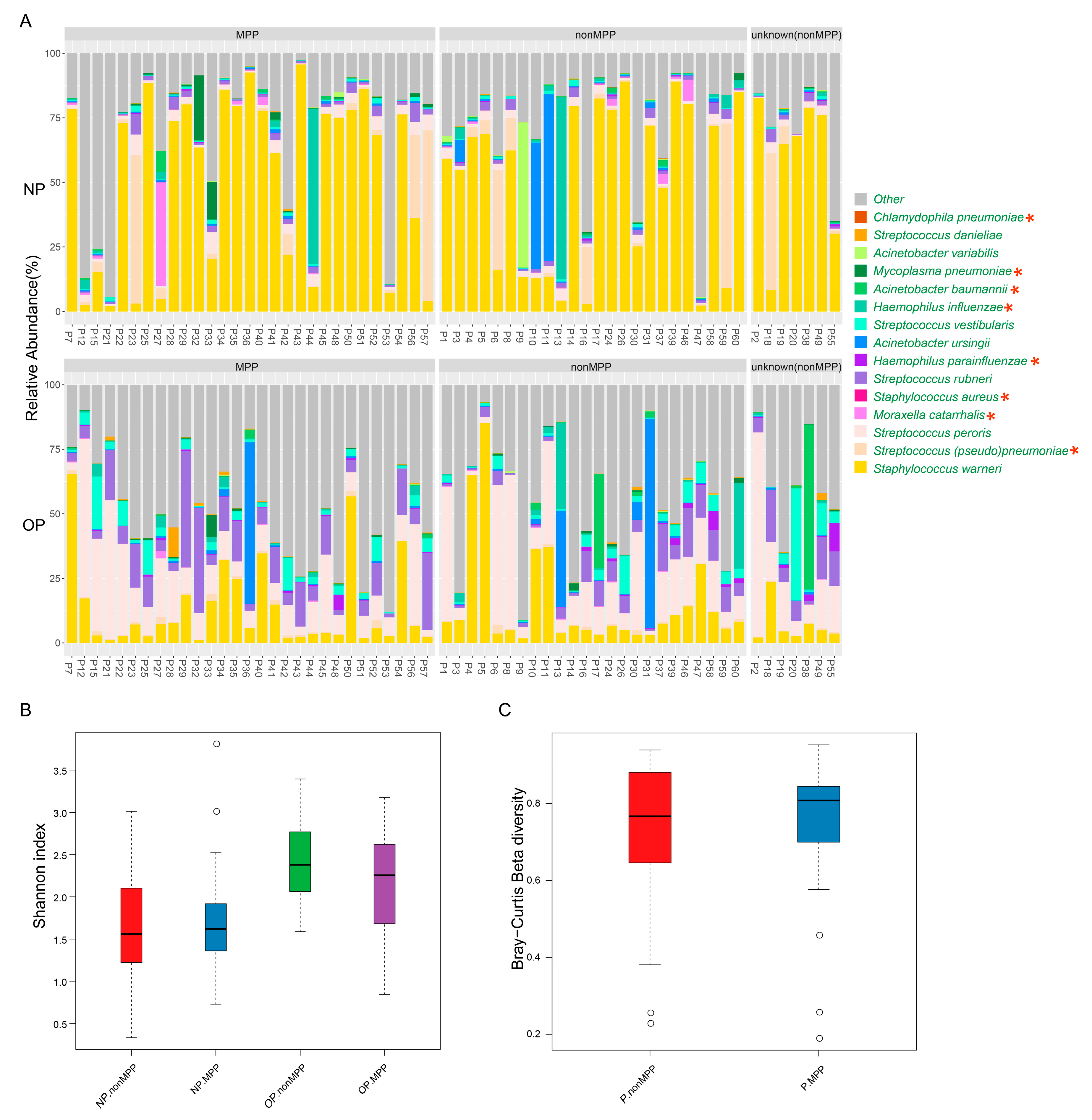
3.4. Microbiota Analysis Can Be Complementary to Clinical Detection of Respiratory Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rudan, I.; O’Brien, K.L.; Nair, H.; Liu, L.; Theodoratou, E.; Qazi, S.; Luksic, I.; Fischer Walker, C.L.; Black, R.E.; Campbell, H.; et al. Epidemiology and etiology of childhood pneumonia in 2010: Estimates of incidence, severe morbidity, mortality, underlying risk factors and causative pathogens for 192 countries. J. Glob. Health 2013, 3, 10401. [Google Scholar] [CrossRef]
- Sakwinska, O.; Bastic Schmid, V.; Berger, B.; Bruttin, A.; Keitel, K.; Lepage, M.; Moine, D.; Ngom Bru, C.; Brussow, H.; Gervaix, A. Nasopharyngeal microbiota in healthy children and pneumonia patients. J. Clin. Microbiol. 2014, 52, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, M.L.; Calvo, C.; Pozo, F.; Villadangos, P.A.; Perez-Brena, P.; Casas, I. Spectrum of respiratory viruses in children with community-acquired pneumonia. Pediatr. Infect. Dis. J. 2012, 31, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Xu, B.; Liu, X.; Shen, K. Status of Mycoplasma Pneumoniae pneumonia in chinese children: A systematic review. Adv. Microbiol. 2014, 4, 704–711. [Google Scholar] [CrossRef]
- De Steenhuijsen Piters, W.A.; Huijskens, E.G.; Wyllie, A.L.; Biesbroek, G.; van den Bergh, M.R.; Veenhoven, R.H.; Wang, X.; Trzcinski, K.; Bonten, M.J.; Rossen, J.W.; et al. Dysbiosis of upper respiratory tract microbiota in elderly pneumonia patients. ISME J. 2016, 10, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Xiao, F.; Wang, C.; Wang, Z. The impact of different methods of DNA extraction on microbial community measures of balf samples based on metagenomic data. Am. J. Transl. Res. 2016, 8, 1412–1425. [Google Scholar] [PubMed]
- Murdoch, D.R.; O’Brien, K.L.; Scott, J.A.; Karron, R.A.; Bhat, N.; Driscoll, A.J.; Knoll, M.D.; Levine, O.S. Breathing new life into pneumonia diagnostics. J. Clin. Microbiol. 2009, 47, 3405–3408. [Google Scholar] [CrossRef] [PubMed]
- Vissing, N.H.; Chawes, B.L.; Bisgaard, H. Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 2013, 188, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, N.; Leonard, E.; Martin, J.M. Prevalence of streptococcal pharyngitis and streptococcal carriage in children: A meta-analysis. Pediatrics 2010, 126, e557–e564. [Google Scholar] [CrossRef] [PubMed]
- Bousbia, S.; Papazian, L.; Saux, P.; Forel, J.M.; Auffray, J.P.; Martin, C.; Raoult, D.; La Scola, B. Repertoire of intensive care unit pneumonia microbiota. PLoS ONE 2012, 7, e32486. [Google Scholar] [CrossRef] [PubMed]
- De Steenhuijsen Piters, W.A.; Heinonen, S.; Hasrat, R.; Bunsow, E.; Smith, B.; Suarez-Arrabal, M.C.; Chaussabel, D.; Cohen, D.M.; Sanders, E.A.; Ramilo, O.; et al. Nasopharyngeal microbiota, host transcriptome, and disease severity in children with respiratory syncytial virus infection. Am. J. Respir. Crit. Care Med. 2016, 194, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.A.; de Steenhuijsen Piters, W.A.; van Houten, M.A.; Chu, M.; Biesbroek, G.; Kool, J.; Pernet, P.; de Groot, P.C.M.; Eijkemans, M.J.C.; Keijser, B.J.F.; et al. Maturation of the infant respiratory microbiota, environmental drivers and health consequences: A prospective cohort study. Am. J. Respir. Crit. Care Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, M.M.; Gent, J.F.; Kong, Y.; Wade, M.; Gansebom, S.; Bramley, A.M.; Jain, S.; Arnold, S.L.; McCullers, J.A. Association of sputum microbiota profiles with severity of community-acquired pneumonia in children. BMC Infect. Dis. 2016, 16, 317. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dai, W.; Qiu, C.; Li, S.; Wang, W.; Xu, J.; Li, Z.; Wang, H.; Li, Y.; Yang, Z.; et al. Mycoplasma Pneumoniae and streptococcus pneumoniae caused different microbial structure and correlation network in lung microbiota. J. Thorac. Dis. 2016, 8, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.J.; Imai, I.; Bittinger, K.; Laughlin, A.; Fuchs, B.D.; Bushman, F.D.; Collman, R.G. Composition and dynamics of the respiratory tract microbiome in intubated patients. Microbiome 2016, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The silva and “all-species living tree project (ltp)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed]
- Biesbroek, G.; Bosch, A.A.; Wang, X.; Keijser, B.J.; Veenhoven, R.H.; Sanders, E.A.; Bogaert, D. The impact of breastfeeding on nasopharyngeal microbial communities in infants. Am. J. Respir. Crit. Care Med. 2014, 190, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.I.; Ernst, R.K.; Bader, M.W. Lps, tlr4 and infectious disease diversity. Nat. Rev. Microbiol. 2005, 3, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Lother, S.A.; Demczuk, W.; Martin, I.; Mulvey, M.; Dufault, B.; Lagace-Wiens, P.; Keynan, Y. Clonal clusters and virulence factors of group c and g streptococcus causing severe infections, Manitoba, Canada, 2012–2014. Emerg. Infect. Dis. 2017, 23, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Bayer-Santos, E.; Durkin, C.H.; Rigano, L.A.; Kupz, A.; Alix, E.; Cerny, O.; Jennings, E.; Liu, M.; Ryan, A.S.; Lapaque, N.; et al. The salmonella effector sted mediates march8-dependent ubiquitination of mhc ii molecules and inhibits t cell activation. Cell Host Microbe 2016, 20, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.G.; Rosa, A.C.; Ferreira, A.S.; Moreira, L.M.; Proenca, D.N.; Morais, P.V.; Cardoso, O. Virulence factors and infection ability of pseudomonas aeruginosa isolates from a hydropathic facility and respiratory infections. J. Appl. Microbiol. 2014, 116, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.J. Antigenic variation with a twist—The borrelia story. Mol. Microbiol. 2006, 60, 1319–1322. [Google Scholar] [CrossRef] [PubMed]
- Reddick, L.E.; Alto, N.M. Bacteria fighting back: How pathogens target and subvert the host innate immune system. Mol. Cell 2014, 54, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Dai, W.; Zhou, Q.; Fu, D.; Zheng, Y.; Wang, W.; Liu, Y.; Yang, Q.; Dai, D.; Liu, S.; et al. Dynamic oropharyngeal and faecal microbiota during treatment in infants hospitalized for bronchiolitis compared with age-matched healthy subjects. Sci. Rep. 2017, 7, 11266. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Bassis, C.M.; Erb-Downward, J.R.; Dickson, R.P.; Freeman, C.M.; Schmidt, T.M.; Young, V.B.; Beck, J.M.; Curtis, J.L.; Huffnagle, G.B. Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. MBio 2015, 6, e00037-15. [Google Scholar] [CrossRef] [PubMed]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.D.; Collman, R.G. Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Sahin-Yilmaz, A.; Naclerio, R.M. Anatomy and physiology of the upper airway. Proc. Am. Thorac. Soc. 2011, 8, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Al Masalma, M.; Armougom, F.; Scheld, W.M.; Dufour, H.; Roche, P.H.; Drancourt, M.; Raoult, D. The expansion of the microbiological spectrum of brain abscesses with use of multiple 16s ribosomal DNA sequencing. Clin. Infect. Dis. 2009, 48, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Uehara, S.; Sunakawa, K.; Eguchi, H.; Ouchi, K.; Okada, K.; Kurosaki, T.; Suzuki, H.; Tsutsumi, H.; Haruta, T.; Mitsuda, T.; et al. Japanese guidelines for the management of respiratory infectious diseases in children 2007 with focus on pneumonia. Pediatr. Int. 2011, 53, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Musher, D.M.; Thorner, A.R. Community-acquired pneumonia. N. Engl. J. Med. 2014, 371, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Self, W.H.; Wunderink, R.G.; Fakhran, S.; Balk, R.; Bramley, A.M.; Grijalva, C.G.; Anderson, E.J.; Courtney, D.M.; Chappell, J.D.; et al. Community-acquired pneumonia requiring ospitalization among U.S. Adults. N. Engl. J. Med. 2015, 373, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Langelier, C.; Zinter, M.S.; Kalantar, K.; Yanik, G.A.; Christenson, S.; O’Donovan, B.; White, C.; Wilson, M.; Sapru, A.; Dvorak, C.C.; et al. Metagenomic sequencing detects respiratory pathogens in hematopoietic cellular transplant patients. Am. J. Respir. Crit. Care Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Yatera, K.; Kawanami, T.; Yamasaki, K.; Fukuda, K.; Naito, K.; Akata, K.; Nagata, S.; Ishimoto, H.; Taniguchi, H.; et al. Pneumonia and empyema caused by streptococcus intermedius that shows the diagnostic importance of evaluating the microbiota in the lower respiratory tract. Intern. Med. 2014, 53, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, K.M.; Erb-Downward, J.R.; Bao, Y.; Branton, W.R.; Falkowski, N.R.; Newton, D.W.; Huffnagle, G.B.; Dickson, R.P. Rapid pathogen identification in bacterial pneumonia using real-time metagenomics. Am. J. Respir. Crit. Care Med. 2017. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Children (n = 59) | Pneumonia Patients (n = 60) | |
|---|---|---|
| Characteristics | ||
| Gender | ||
| Female | 33 | 19 |
| Male | 26 | 41 |
| Age (years) | 2.8 (0.1–9.9) | 2.8 (0.2–12.7) |
| Delivery Mode | ||
| Caesarean section | 20 | 22 |
| Vaginally born | 39 | 38 |
| Feed Pattern | ||
| Breast feed | 18 | 40 |
| Breast feed + Milk feed | 31 | 6 |
| Milk feed | 10 | 14 |
| Family history of allergy | - | 1 |
| History of pneumonia | - | 12 |
| Asthma | - | - |
| Clinical records | ||
| Lung consolidation, atelectasis, infiltration | NA | 60 |
| Hospitalization time (days) | - | 9 (2–37) |
| Fever | - | 26 |
| Cough | - | 57 |
| Wheezing | - | 16 |
| CRP (<0.499 mg/L) | NA | 21 |
| PCT (<0.5 ng/mL) | NA | 60 |
| Eosinophil (0.5–5%) | NA | 33 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Dai, W.; Liu, Y.; Zhou, Q.; Wang, H.; Li, D.; Yang, Z.; Li, Y.; Xie, G.; Li, S.; et al. The Alteration of Nasopharyngeal and Oropharyngeal Microbiota in Children with MPP and Non-MPP. Genes 2017, 8, 380. https://doi.org/10.3390/genes8120380
Lu Z, Dai W, Liu Y, Zhou Q, Wang H, Li D, Yang Z, Li Y, Xie G, Li S, et al. The Alteration of Nasopharyngeal and Oropharyngeal Microbiota in Children with MPP and Non-MPP. Genes. 2017; 8(12):380. https://doi.org/10.3390/genes8120380
Chicago/Turabian StyleLu, Zhiwei, Wenkui Dai, Yanhong Liu, Qian Zhou, Heping Wang, Dongfang Li, Zhenyu Yang, Yinhu Li, Gan Xie, Shuaicheng Li, and et al. 2017. "The Alteration of Nasopharyngeal and Oropharyngeal Microbiota in Children with MPP and Non-MPP" Genes 8, no. 12: 380. https://doi.org/10.3390/genes8120380
APA StyleLu, Z., Dai, W., Liu, Y., Zhou, Q., Wang, H., Li, D., Yang, Z., Li, Y., Xie, G., Li, S., & Zheng, Y. (2017). The Alteration of Nasopharyngeal and Oropharyngeal Microbiota in Children with MPP and Non-MPP. Genes, 8(12), 380. https://doi.org/10.3390/genes8120380