
Telomere and Telomerase Therapeutics in Cancer

Abstract
:1. Introduction
2. Telomere Biology
3. Telomerase Biology
4. Telomerase as a Cancer Biomarker
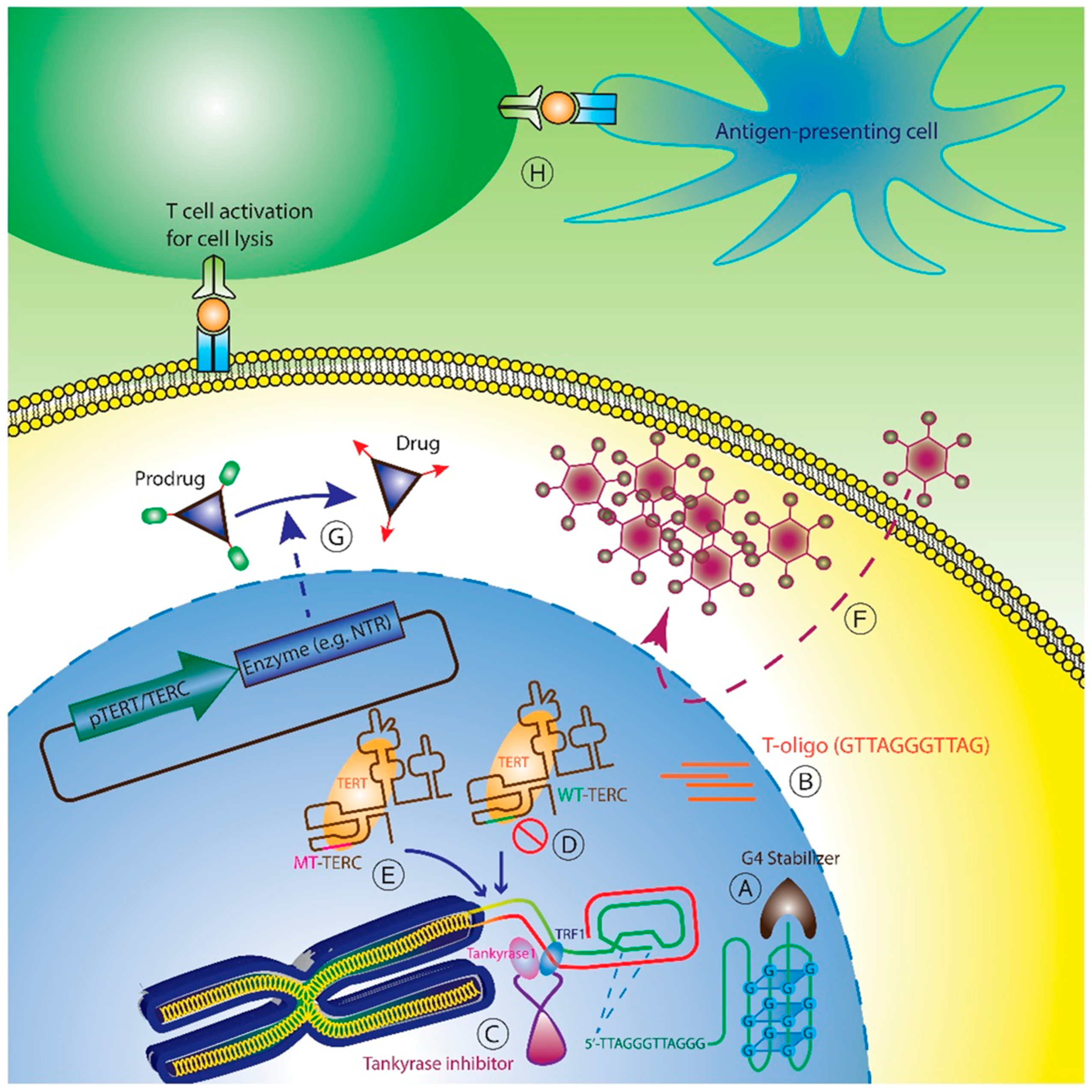
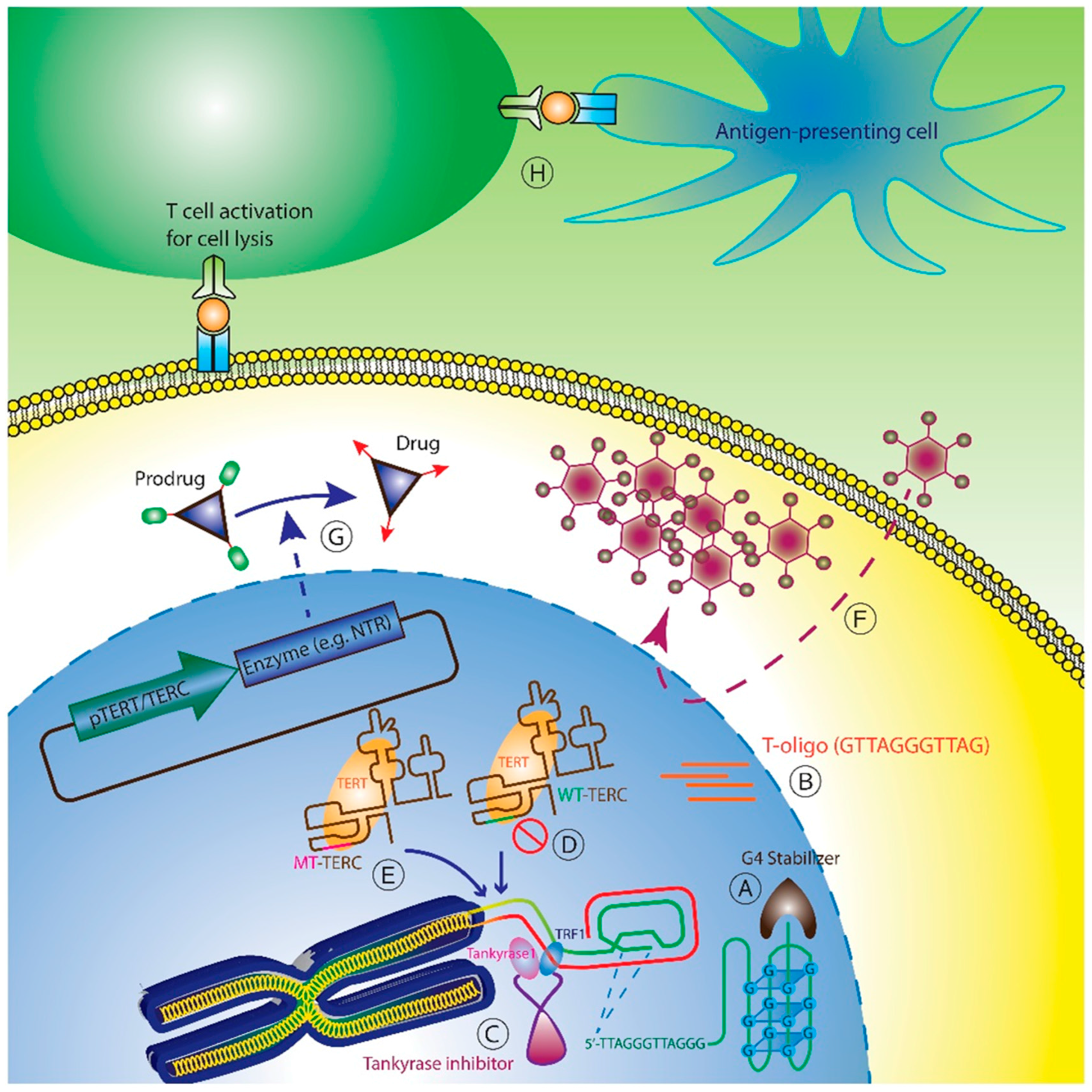
5. Telomere and Telomerase Therapeutics
5.1. Telomere-Based Strategies
5.2. Direct Telomerase Inhibition
5.3. Telomerase Interference
5.4. TERT or TERC Promoter Driven Therapy
5.5. Telomerase Immunotherapy
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; de Lange, T. Mammalian telomeres end in a large duplex loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Broccoli, D.; Smogorzewska, A.; Chong, L.; de Lange, T. Human telomeres contain two distinct Myb-related proteins, TRF1 and TRF2. Nat. Genet. 1997, 17, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.; Stansel, R.M.; Fairall, L.; Griffith, J.D.; Rhodes, D.; de Lange, T. TRF1 binds a bipartite telomeric site with extreme spatial flexibility. EMBO J. 1999, 18, 5735–5744. [Google Scholar] [CrossRef] [PubMed]
- Loayza, D.; de Lange, T. POT1 as a terminal transducer of TRF1 telomere length control. Nature 2003, 423, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Oestreich, S.; de Lange, T. Identification of human Rap1: Implications for telomere evolution. Cell 2000, 101, 471–483. [Google Scholar] [CrossRef]
- Arat, N.Ö.; Griffith, J.D. Human Rap1 interacts directly with telomeric DNA and regulates TRF2 localization at the telomere. J. Biol. Chem. 2012, 287, 41583–41594. [Google Scholar] [CrossRef] [PubMed]
- Janoušková, E.; Nečasová, I.; Pavloušková, J.; Zimmermann, M.; Hluchý, M.; Marini, V.; Nováková, M.; Hofr, C. Human Rap1 modulates TRF2 attraction to telomeric DNA. Nucleic acids Res. 2015, 43, 2691–2700. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Z.; Donigian, J.R.; van Overbeek, M.; Loayza, D.; Luo, Y.; Krutchinsky, A.N.; Chait, B.T.; de Lange, T. Tin2 binds trf1 and TRF2 simultaneously and stabilizes the TRF2 complex on telomeres. J. Biol. Chem. 2004, 279, 47264–47271. [Google Scholar] [CrossRef] [PubMed]
- Frescas, D.; de Lange, T. Binding of TPP1 protein to TIN2 protein is required for POT1a, b protein-mediated telomere protection. J. Biol. Chem. 2014, 289, 24180–24187. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Safari, A.; O’Connor, M.S.; Chan, D.W.; Laegeler, A.; Qin, J.; Songyang, Z. Ptop interacts with POT1 and regulates its localization to telomeres. Nat. Cell Boil. 2004, 6, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.D. Origin of concatemeric T7 DNA. Nat. New Boil. 1972, 239, 197–201. [Google Scholar] [CrossRef]
- Xu, Y.; He, K.; Goldkorn, A. Telomerase targeted therapy in cancer and cancer stem cells. Clin. Adv. Hematol. Oncol. 2011, 9, 442–455. [Google Scholar] [PubMed]
- Hahn, W.C.; Weinberg, R.A. Rules for making human tumor cells. N. Engl. J. Med. 2002, 347, 1593–1603. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Balk, B.; Maicher, A.; Dees, M.; Klermund, J.; Luke-Glaser, S.; Bender, K.; Luke, B. Telomeric RNA-DNA hybrids affect telomere-length dynamics and senescence. Nat. Struct. Mol. Boil. 2013, 20, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Porro, A.; Feuerhahn, S.; Delafontaine, J.; Riethman, H.; Rougemont, J.; Lingner, J. Functional characterization of the TERRA transcriptome at damaged telomeres. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Redon, S.; Reichenbach, P.; Lingner, J. The non-coding RNA TERRA is a natural ligand and direct inhibitor of human telomerase. Nucleic Acids Res. 2010, 38, 5797–5806. [Google Scholar] [CrossRef] [PubMed]
- Morin, G.B. The human telomere terminal transferase enzyme is a ribonucleoprotein that synthesizes TTAGGG repeats. Cell 1989, 59, 521–529. [Google Scholar] [CrossRef]
- Venteicher, A.S.; Abreu, E.B.; Meng, Z.; McCann, K.E.; Terns, R.M.; Veenstra, T.D.; Terns, M.P.; Artandi, S.E. A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis. Science 2009, 323, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.R.; Wood, E.; Collins, K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature 1999, 402, 551–555. [Google Scholar] [PubMed]
- Fu, D.; Collins, K. Purification of human telomerase complexes identifies factors involved in telomerase biogenesis and telomere length regulation. Mol. Cell 2007, 28, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Venteicher, A.S.; Meng, Z.; Mason, P.J.; Veenstra, T.D.; Artandi, S.E. Identification of atpases pontin and reptin as telomerase components essential for holoenzyme assembly. Cell 2008, 132, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Savage, S.A.; Shkreli, M.; Giri, N.; Jessop, L.; Myers, T.; Chen, R.; Alter, B.P.; Artandi, S.E. Disruption of telomerase trafficking by TCAB1 mutation causes dyskeratosis congenita. Genes Dev. 2011, 25, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, T.; Beswick, R.; Kirwan, M.; Marrone, A.; Digweed, M.; Walne, A.; Dokal, I. Mutations in the telomerase component NHP2 cause the premature ageing syndrome dyskeratosis congenita. Proc. Natl. Acad. Sci. USA 2008, 105, 8073–8078. [Google Scholar] [CrossRef] [PubMed]
- Walne, A.J.; Vulliamy, T.; Marrone, A.; Beswick, R.; Kirwan, M.; Masunari, Y.; Al-Qurashi, F.h.; Aljurf, M.; Dokal, I. Genetic heterogeneity in autosomal recessive dyskeratosis congenita with one subtype due to mutations in the telomerase-associated protein NOP10. Hum. Mol. Genet. 2007, 16, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Almeida, A.; Pópulo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Oishi, T.; Kigawa, J.; Minagawa, Y.; Shimada, M.; Takahashi, M.; Terakawa, N. Alteration of telomerase activity associated with development and extension of epithelial ovarian cancer. Obstet. Gynecol. 1998, 91, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.A.; Kim, N.W.; Goodman, S.; Marks, J.; Henderson, G.; Umbricht, C.B.; Dome, J.S.; Dooley, W.; Amshey, S.R.; Sukumar, S. Telomerase activity and prognosis in primary breast cancers. J. Clin. Oncol. 1999, 17, 3075–3081. [Google Scholar] [PubMed]
- Okayasu, I.; Mitomi, H.; Yamashita, K.; Mikami, T.; Fujiwara, M.; Kato, M.; Oshimura, M. Telomerase activity significantly correlates with cell differentiation, proliferation and lymph node metastasis in colorectal carcinomas. J. Cancer Res. Clin. Oncol. 1998, 124, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Pirker, C.; Holzmann, K.; Spiegl-Kreinecker, S.; Elbling, L.; Thallinger, C.; Pehamberger, H.; Micksche, M.; Berger, W. Chromosomal imbalances in primary and metastatic melanomas: Over-representation of essential telomerase genes. Melanoma Res. 2003, 13, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Lu, B.; Tai, Y.C.; Goldkorn, A. A cancer detection platform which measures telomerase activity from live circulating tumor cells captured on a microfilter. Cancer Res. 2010, 70, 6420–6426. [Google Scholar] [CrossRef] [PubMed]
- Goldkorn, A.; Ely, B.; Quinn, D.I.; Tangen, C.M.; Fink, L.M.; Xu, T.; Twardowski, P.; Van Veldhuizen, P.J.; Agarwal, N.; Carducci, M.A.; et al. Circulating tumor cell counts are prognostic of overall survival in swog s0421: A phase III trial of docetaxel with or without atrasentan for metastatic castration-resistant prostate cancer. J. Clin. Oncol. 2014, 32, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhang, C.; Zhu, G.; Irwin, M.; Risch, H.; Menato, G.; Mitidieri, M.; Katsaros, D.; Yu, H. Telomerase expression and telomere length in breast cancer and their associations with adjuvant treatment and disease outcome. Breast Cancer Res. 2011. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, P.F.; Geng, M.; Cao, Y.C.; Yin, Y.C. Correlation between chemosensitivity to anticancer drugs and telomerase reverse transcriptase mRNA expression in gastric cancer. Diagn. Pathol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wesbuer, S.; Lanvers-Kaminsky, C.; Duran-Seuberth, I.; Bolling, T.; Schafer, K.L.; Braun, Y.; Willich, N.; Greve, B. Association of telomerase activity with radio- and chemosensitivity of neuroblastomas. Radiat. Oncol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Deville, L.; Hillion, J.; Pendino, F.; Samy, M.; Nguyen, E.; Segal-Bendirdjian, E. hTERT promotes imatinib resistance in chronic myeloid leukemia cells: Therapeutic implications. Mol. Cancer Ther. 2011, 10, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, E.M.; Erickson, P.A.; Gentry, E.R.; Gryaznov, S.M.; Herbert, B.S. Lipid-conjugated telomerase template antagonists sensitize resistant HER2-positive breast cancer cells to trastuzumab. Breast Cancer Res. Treat. 2009, 118, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Ma, N.N.; Zhou, F.G.; Zhang, L.; Bu, X.X.; Sun, K.; Song, J.R.; Li, R.; Zhang, B.H.; Wu, M.C.; et al. Up-regulation of htert expression by low-dose cisplatin contributes to chemotherapy resistance in human hepatocellular cancer cells. Oncol. Rep. 2009, 22, 549–556. [Google Scholar] [PubMed]
- Mukherjee, S.; Bhattacharya, R.K.; Roy, M. Targeting protein kinase C (PKC) and telomerase by phenethyl isothiocyanate (PEITC) sensitizes PC-3 cells towards chemotherapeutic drug-induced apoptosis. J. Environ. Pathol. Toxicol. Oncol. 2009, 28, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Paranjape, A.N.; Mandal, T.; Mukherjee, G.; Kumar, M.V.; Sengupta, K.; Rangarajan, A. Introduction of SV40ER and hTERT into mammospheres generates breast cancer cells with stem cell properties. Oncogene 2012, 31, 1896–1909. [Google Scholar] [CrossRef] [PubMed]
- Joseph, I.; Tressler, R.; Bassett, E.; Harley, C.; Buseman, C.M.; Pattamatta, P.; Wright, W.E.; Shay, J.W.; Go, N.F. The telomerase inhibitor imetelstat depletes cancer stem cells in breast and pancreatic cancer cell lines. Cancer Res. 2010, 70, 9494–9504. [Google Scholar] [CrossRef] [PubMed]
- Marian, C.O.; Cho, S.K.; McEllin, B.M.; Maher, E.A.; Hatanpaa, K.J.; Madden, C.J.; Mickey, B.E.; Wright, W.E.; Shay, J.W.; Bachoo, R.M. The telomerase antagonist, imetelstat, efficiently targets glioblastoma tumor-initiating cells leading to decreased proliferation and tumor growth. Clin. Cancer Res. 2010, 16, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Marian, C.O.; Wright, W.E.; Shay, J.W. The effects of telomerase inhibition on prostate tumor-initiating cells. Int. J. Cancer 2010, 127, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Serrano, D.; Bleau, A.M.; Fernandez-Garcia, I.; Fernandez-Marcelo, T.; Iniesta, P.; Ortiz-de-Solorzano, C.; Calvo, A. Inhibition of telomerase activity preferentially targets aldehyde dehydrogenase-positive cancer stem-like cells in lung cancer. Mol. Cancer 2011. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; He, K.; Wang, L.; Goldkorn, A. Prostate tumor cells with cancer progenitor properties have high telomerase activity and are rapidly killed by telomerase interference. Prostate 2011, 71, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tergaonkar, V. Noncanonical functions of telomerase: Implications in telomerase-targeted cancer therapies. Cancer Res. 2014, 74, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Saginc, G.; Leow, S.C.; Khattar, E.; Shin, E.M.; Yan, T.D.; Wong, M.; Zhang, Z.; Li, G.; Sung, W.K. Telomerase directly regulates NF-κB-dependent transcription. Nat. Cell Boil. 2012, 14, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Xi, P.; Zhou, J.; Wang, M.; Cong, Y.S. Human telomerase reverse transcriptase regulates MMP expression independently of telomerase activity via NF-κB-dependent transcription. FASEB J. 2013, 27, 4375–4383. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.A.; Hahn, W.C.; O’Connor, B.F.; Banner, E.N.; Lundberg, A.S.; Modha, P.; Mizuno, H.; Brooks, M.W.; Fleming, M.; Zimonjic, D.B. Telomerase contributes to tumorigenesis by a telomere length-independent mechanism. Proc. Natl. Acad. Sci. USA 2002, 99, 12606–12611. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Firpo, E.J.; Wang, Y.; Roberts, J.M. Separation of telomerase functions by reverse genetics. Proc. Natl. Acad. Sci. USA 2011, 108, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.L.; Coller, H.A.; Roberts, J.M. Telomerase modulates expression of growth-controlling genes and enhances cell proliferation. Nat. Cell Boil. 2003, 5, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Flores, I.; Cayuela, M.L.; Blasco, M.A. Effects of telomerase and telomere length on epidermal stem cell behavior. Science 2005, 309, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Beck, S.; Jin, X.; Sohn, Y.W.; Kim, J.K.; Kim, S.H.; Yin, J.; Pian, X.; Kim, S.C.; Nam, D.H.; Choi, Y.J. Telomerase activity-independent function of TERT allows glioma cells to attain cancer stem cell characteristics by inducing EGFR expression. Mol. Cells 2011, 31, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Q.; Li, K.; Chen, L.; Li, W.; Hou, M.; Liu, T.; Yang, J.; Lindvall, C.; Björkholm, M. Telomerase reverse transcriptase promotes epithelial-mesenchymal transition and stem cell-like traits in cancer cells. Oncogene 2013, 32, 4203–4213. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesh, S.; Banerjee, P.P. Telomerase reverse transcriptase regulates the expression of a key cell cycle regulator, cyclin D1. Biochem. Biophys. Res. Commun. 2006, 347, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.M.; Khattar, E.; Leow, S.C.; Liu, C.Y.; Muller, J.; Ang, W.X.; Li, Y.; Franzoso, G.; Li, S.; Guccione, E.; et al. Telomerase regulates MYC-driven oncogenesis independent of its reverse transcriptase activity. J. Clin. Invest. 2015, 125, 2109–2122. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Nelson, A.D.; Shippen, D.E. A transposable element within the non-canonical telomerase RNA of arabidopsis thaliana modulates telomerase in response to DNA damage. PLoS Genet. 2015. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz-Pérez, F.; García-Castillo, J.; García-Moreno, D.; López-Muñoz, A.; Anchelin, M.; Angosto, D.; Zon, L.I.; Mulero, V.; Cayuela, M.L. A non-canonical function of telomerase RNA in the regulation of developmental myelopoiesis in zebrafish. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wright, W.E. Telemerase activity in human germiine and embryanie tissues and cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Forsyth, N.R.; Wright, W.E.; Shay, J.W. Telomerase and differentiation in multicellular organisms: Turn it off, turn it on, and turn it off again. Differentiation 2002, 69, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Poremba, C.; Heine, B.; Diallo, R.; Heinecke, A.; Wai, D.; Schaefer, K.L.; Braun, Y.; Schuck, A.; Lanvers, C.; Bankfalvi, A. Telomerase as a prognostic marker in breast cancer: High-throughput tissue microarray analysis of hTERT and HTR. J. Pathol. 2002, 198, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Kiyozuka, Y.; Ichiyoshi, H.; Senzaki, H.; Takada, H.; Hioki, K.; Tsubura, A. Change in telomerase activity during human colorectal carcinogenesis. Anticancer Res. 1998, 19, 2167–2172. [Google Scholar]
- Miyoshi, Y.; Tsukinoki, K.; Imaizumi, T.; Yamada, Y.; Ishizaki, T.; Watanabe, Y.; Sasakura, Y.; Lin, Y.; Hosaka, M.; Kubota, Y. Telomerase activity in oral cancer. Oral Oncol. 1999, 35, 283–289. [Google Scholar] [CrossRef]
- Mori, K.; Sato, S.; Kodama, M.; Habu, M.; Takahashi, O.; Nishihara, T.; Tominaga, K.; Takenaka, S. Oral cancer diagnosis via a ferrocenylnaphthalene diimide-based electrochemical telomerase assay. Clin. Chem. 2013, 59, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Ishida, M.; Motoi, F.; Yamaguchi, T.; Naitoh, T.; Katayose, Y.; Egawa, S.; Unno, M. Telomerase activity in pancreatic juice differentiates pancreatic cancer from chronic pancreatitis: A meta-analysis. Pancreatology 2016. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Nayak, S.; Kakar, R.; Chaudhari, U.K.; Joshi, D.; Vundinti, B.R.; Fernandes, G.; Barai, R.S.; Kholkute, S.D.; Sachdeva, G. A triad of telomerase, androgen receptor and early growth response 1 in prostate cancer cells. Cancer Biol. Ther. 2016, 17, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, R.; Seto, M.; Tsumuki, H.; Iida, K.; Yamazaki, T.; Sonoda, J.; Matsumine, A.; Uchida, A. Telomerase activity and human telomerase reverse transcriptase mRNA expression are correlated with clinical aggressiveness in soft tissue tumors. Cancer 2002, 95, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Broccoli, D.; Young, J.W.; de Lange, T. Telomerase activity in normal and malignant hematopoietic cells. Proc. Natl. Acad. Sci. USA 1995, 92, 9082–9086. [Google Scholar] [CrossRef] [PubMed]
- Soria, J.C.; Gauthier, L.R.; Raymond, E.; Granotier, C.; Morat, L.; Armand, J.P.; Boussin, F.D.; Sabatier, L. Molecular detection of telomerase-positive circulating epithelial cells in metastatic breast cancer patients. Clin. Cancer Res. 1999, 5, 971–975. [Google Scholar] [PubMed]
- Sapi, E.; Okpokwasili, N.I.; Rutherford, T. Detection of telomerase-positive circulating epithelial cells in ovarian cancer patients. Cancer Detect. Prev. 2002, 26, 158–167. [Google Scholar] [CrossRef]
- Goldkorn, A.; Ely, B.; Tangen, C.M.; Tai, Y.C.; Xu, T.; Li, H.; Twardowski, P.; Veldhuizen, P.J.; Agarwal, N.; Carducci, M.A.; et al. Circulating tumor cell telomerase activity as a prognostic marker for overall survival in swog 0421: A phase III metastatic castration resistant prostate cancer trial. Int. J. Cancer 2015, 136, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Kyo, S.; Kanaya, T.; Takakura, M.; Inoue, M.; Namiki, M. Expression of human telomerase subunits and correlation with telomerase activity in urothelial cancer. Clin. Cancer Res. 1998, 4, 1603–1608. [Google Scholar] [PubMed]
- Hara, H.; Yamashita, K.; Shinada, J.; Yoshimura, H.; Kameya, T. Clinicopathologic significance of telomerase activity and hTERT mRNA expression in non-small cell lung cancer. Lung Cancer 2001, 34, 219–226. [Google Scholar] [CrossRef]
- Kirkpatrick, K.L.; Clark, G.; Ghilchick, M.; Newbold, R.F.; Mokbel, K. hTERT mRNA expression correlates with telomerase activity in human breast cancer. Eur. J. Surg. Oncol. 2003, 29, 321–326. [Google Scholar] [CrossRef] [PubMed]
- March-Villalba, J.A.; Martinez-Jabaloyas, J.M.; Herrero, M.J.; Santamaria, J.; Alino, S.F.; Dasi, F. Cell-free circulating plasma hTERT mRNA is a useful marker for prostate cancer diagnosis and is associated with poor prognosis tumor characteristics. PLoS ONE 2012, 7, e43470. [Google Scholar] [CrossRef] [PubMed]
- Ping, B.; Tsuno, S.; Wang, X.; Ishihara, Y.; Yamashita, T.; Miura, K.; Miyoshi, F.; Shinohara, Y.; Matsuki, T.; Tanabe, Y.; et al. Comparative study of (1)(8)F-FDG-PET/CT imaging and serum hTERT mRNA quantification in cancer diagnosis. Cancer Med. 2015, 4, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Avilion, A.A.; Piatyszek, M.A.; Gupta, J.; Shay, J.W.; Bacchetti, S.; Greider, C.W. Human telomerase RNA and telomerase activity in immortal cell lines and tumor tissues. Cancer Res. 1996, 56, 645–650. [Google Scholar] [PubMed]
- Sallinen, P.; Miettinen, H.; Sallinen, S.L.; Haapasalo, H.; Helin, H.; Kononen, J. Increased expression of telomerase RNA component is associated with increased cell proliferation in human astrocytomas. Am. J. Pathol. 1997, 150, 1159–1164. [Google Scholar] [PubMed]
- Heine, B.; Hummel, M.; Demel, G.; Stein, H. Demonstration of constant upregulation of the telomerase RNA component in human gastric carcinomas using in situ hybridization. J. Pathol. 1998, 185, 139–144. [Google Scholar] [CrossRef]
- Hiyama, T.; Yokozaki, H.; Kitadai, Y.; Haruma, K.; Yasui, W.; Kajiyama, G.; Tahara, E. Overexpression of human telomerase RNA is an early event in oesophageal carcinogenesis. Virchows Arch. 1999, 434, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Bonnefoi, H.; Pelte, M.F.; Lyautey, J.; Lederrey, C.; Movarekhi, S.; Schaeffer, P.; Mulcahy, H.E.; Meyer, P.; Stroun, M.; et al. Telomerase RNA as a detection marker in the serum of breast cancer patients. Clin. Cancer Res. 2000, 6, 3823–3826. [Google Scholar] [PubMed]
- Bialkowska-Hobrzanska, H.; Bowles, L.; Bukala, B.; Joseph, M.G.; Fletcher, R.; Razvi, H. Comparison of human telomerase reverse transcriptase messenger RNA and telomerase activity as urine markers for diagnosis of bladder carcinoma. Mol. Diagn. 2000, 5, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Krause, H.; Heicappell, R.; Tischendorf, J.; Shay, J.W.; Miller, K. Comparison of human telomerase RNA and telomerase activity in urine for diagnosis of bladder cancer. Clin. Cancer Res. 1998, 4, 1949–1954. [Google Scholar] [PubMed]
- Eissa, S.; Swellam, M.; Ali-Labib, R.; Mansour, A.; El-Malt, O.; Tash, F.M. Detection of telomerase in urine by 3 methods: Evaluation of diagnostic accuracy for bladder cancer. J. Urol. 2007, 178, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative Lengthening of Telomeres in Mammalian Cells; Landes Bioscience: Austin, TX, USA, 2000. [Google Scholar]
- Mocellin, S.; Pooley, K.A.; Nitti, D. Telomerase and the search for the end of cancer. Trends Mol. Med. 2013, 19, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.W.; Feigon, J. Quadruplex structure of oxytricha telomeric DNA oligonucleotides. Nature 1992, 356, 164–168. [Google Scholar] [PubMed]
- Bochman, M.L.; Paeschke, K.; Zakian, V.A. DNA secondary structures: Stability and function of G-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Moye, A.L.; Porter, K.C.; Cohen, S.B.; Phan, T.; Zyner, K.G.; Sasaki, N.; Lovrecz, G.O.; Beck, J.L.; Bryan, T.M. Telomeric g-quadruplexes are a substrate and site of localization for human telomerase. Nat. Commun. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, J.Q.; Chen, Z.; Zheng, K.W.; Chen, C.Y.; Hao, Y.H.; Tan, Z. G-quadruplex formation at the 3' end of telomere DNA inhibits its extension by telomerase, polymerase and unwinding by helicase. Nucleic Acids Res. 2011, 39, 6229–6237. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Tang, J.; Kan, Z.Y.; Hao, Y.H. Telomere G-quadruplex as a potential target to accelerate telomere shortening by expanding the incomplete end-replication of telomere DNA. Curr. Top. Med. Chem. 2015, 15, 1940–1946. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, V.; Salvati, E.; Alvino, A.; Bianco, A.; Ciammaichella, A.; D’Angelo, C.; Ginnari-Satriani, L.; Serrilli, A.M.; Iachettini, S.; Leonetti, C. N-cyclic bay-substituted perylene G-quadruplex ligands have selective antiproliferative effects on cancer cells and induce telomere damage. J. Med. Chem. 2011, 54, 1140–1156. [Google Scholar] [CrossRef] [PubMed]
- Leonetti, C.; Amodei, S.; D’Angelo, C.; Rizzo, A.; Benassi, B.; Antonelli, A.; Elli, R.; Stevens, M.F.; D’Incalci, M.; Zupi, G. Biological activity of the G-quadruplex ligand RHPS4 (3, 11-difluoro-6, 8, 13-trimethyl-8h-quino [4, 3, 2-kl] acridinium methosulfate) is associated with telomere capping alteration. Mol. Pharmacol. 2004, 66, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Phatak, P.; Cookson, J.; Dai, F.; Smith, V.; Gartenhaus, R.; Stevens, M.; Burger, A. Telomere uncapping by the G-quadruplex ligand RHPS4 inhibits clonogenic tumour cell growth in vitro and in vivo consistent with a cancer stem cell targeting mechanism. Br. J. Cancer 2007, 96, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Mikami-Terao, Y.; Akiyama, M.; Yuza, Y.; Yanagisawa, T.; Yamada, O.; Yamada, H. Antitumor activity of G-quadruplex-interactive agent TMPyP4 in K562 leukemic cells. Cancer Lett. 2008, 261, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, J.; Matsuo, T.; Shimose, S.; Kubo, T.; Ishikawa, M.; Yasunaga, Y.; Ochi, M. Antitumor effects of telomerase inhibitor TMPyP4 in osteosarcoma cell lines. J. Orthop. Res. 2011, 29, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Shammas, M.A.; Shmookler Reis, R.J.; Akiyama, M.; Koley, H.; Chauhan, D.; Hideshima, T.; Goyal, R.K.; Hurley, L.H.; Anderson, K.C.; Munshi, N.C. Telomerase inhibition and cell growth arrest by G-quadruplex interactive agent in multiple myeloma. Mol. Cancer Ther. 2003, 2, 825–833. [Google Scholar] [PubMed]
- Leonetti, C.; Scarsella, M.; Riggio, G.; Rizzo, A.; Salvati, E.; D’Incalci, M.; Staszewsky, L.; Frapolli, R.; Stevens, M.F.; Stoppacciaro, A.; et al. G-quadruplex ligand RHPS4 potentiates the antitumor activity of camptothecins in preclinical models of solid tumors. Clin. Cancer Res. 2008, 14, 7284–7291. [Google Scholar] [CrossRef] [PubMed]
- Berardinelli, F.; Siteni, S.; Tanzarella, C.; Stevens, M.F.; Sgura, A.; Antoccia, A. The G-quadruplex-stabilising agent RHPS4 induces telomeric dysfunction and enhances radiosensitivity in glioblastoma cells. DNA Repair 2015, 25, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Burger, A.M.; Dai, F.; Schultes, C.M.; Reszka, A.P.; Moore, M.J.; Double, J.A.; Neidle, S. The G-quadruplex-interactive molecule BRACO-19 inhibits tumor growth, consistent with telomere targeting and interference with telomerase function. Cancer Res. 2005, 65, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Gowan, S.M.; Harrison, J.R.; Patterson, L.; Valenti, M.; Read, M.A.; Neidle, S.; Kelland, L.R. A G-quadruplex-interactive potent small-molecule inhibitor of telomerase exhibiting in vitro and in vivo antitumor activity. Mol. Pharmacol. 2002, 61, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, X.; Li, Y.; Xu, S.; Ma, C.; Wu, X.; Cheng, Y.; Yu, Z.; Zhao, G.; Chen, Y. Telomere targeting with a novel G-quadruplex-interactive ligand BRACO-19 induces T-loop disassembly and telomerase displacement in human glioblastoma cells. Oncotarget 2016. [Google Scholar] [CrossRef]
- Waki, K.; Anno, K.; Ono, T.; Ide, T.; Chayama, K.; Tahara, H. Establishment of functional telomerase immortalized human hepatocytes and a hepatic stellate cell line for telomere-targeting anticancer drug development. Cancer Sci. 2010, 101, 1678–1685. [Google Scholar] [CrossRef] [PubMed]
- Doi, T.; Shibata, K.; Yoshida, M.; Takagi, M.; Tera, M.; Nagasawa, K.; Shin-ya, K.; Takahashi, T. (S)-stereoisomer of telomestatin as a potent G-quadruplex binder and telomerase inhibitor. Org. Biomol. Chem. 2011, 9, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Vankayalapati, H.; Shin-Ya, K.; Wierzba, K.; Hurley, L.H. Telomestatin, a potent telomerase inhibitor that interacts quite specifically with the human telomeric intramolecular G-quadruplex. J. Am. Chem. Soc. 2002, 124, 2098–2099. [Google Scholar] [CrossRef] [PubMed]
- Tauchi, T.; Shin-Ya, K.; Sashida, G.; Sumi, M.; Nakajima, A.; Shimamoto, T.; Ohyashiki, J.H.; Ohyashiki, K. Activity of a novel G-quadruplex-interactive telomerase inhibitor, telomestatin (sot-095), against human leukemia cells: Involvement of atm-dependent DNA damage response pathways. Oncogene 2003, 22, 5338–5347. [Google Scholar] [PubMed]
- Hasegawa, D.; Okabe, S.; Okamoto, K.; Nakano, I.; Shin-Ya, K.; Seimiya, H. G-quadruplex ligand-induced DNA damage response coupled with telomere dysfunction and replication stress in glioma stem cells. Biochem. Biophys. Res. Commun. 2016, 471, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Drygin, D.; Siddiqui-Jain, A.; O'Brien, S.; Schwaebe, M.; Lin, A.; Bliesath, J.; Ho, C.B.; Proffitt, C.; Trent, K.; Whitten, J.P.; et al. Anticancer activity of CX-3543: A direct inhibitor of rRNA biogenesis. Cancer Res. 2009, 69, 7653–7661. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Eller, M.S.; Panova, I.; Kubera, J.; Wee, L.H.; Cowan, K.H.; Gilchrest, B.A. Telomeric DNA induces apoptosis and senescence of human breast carcinoma cells. Breast Cancer Res. 2007. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Iwado, E.; Eller, M.S.; Kondo, Y.; Fujiwara, K.; Li, G.-Z.; Hess, K.R.; Siwak, D.R.; Sawaya, R.; Mills, G.B. Telomere 3′ overhang-specific DNA oligonucleotides induce autophagy in malignant glioma cells. FASEB J. 2007, 21, 2918–2930. [Google Scholar] [CrossRef] [PubMed]
- Puri, N.; Eller, M.S.; Byers, H.R.; Dykstra, S.; Kubera, J.; Gilchrest, B.A. Telomere-based DNA damage responses: A new approach to melanoma. FASEB J. 2004, 18, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tsolou, A.; Passos, J.F.; Nelson, G.; Arai, Y.; von Zglinicki, T. ssDNA fragments induce cell senescence by telomere uncapping. Exp. Gerontol. 2008, 43, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Faller, D.V. T-oligos inhibit growth and induce apoptosis in human ovarian cancer cells. Oligonucleotides 2011, 21, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Pitman, R.T.; Wojdyla, L.; Puri, N. Mechanism of DNA damage responses induced by exposure to an oligonucleotide homologous to the telomere overhang in melanoma. Oncotarget 2013, 4, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Giriat, I.; Schmitt, A.; de Lange, T. Tankyrase, a poly(adp-ribose) polymerase at human telomeres. Science 1998, 282, 1484–1487. [Google Scholar] [CrossRef] [PubMed]
- Smogorzewska, A.; de Lange, T. Regulation of telomerase by telomeric proteins. Annu. Rev. Biochem. 2004, 73, 177–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Perrem, K.; Harper, J.W.; Lu, K.P.; Zhou, X.Z. The F-box protein FBX4 targets PIN2/TRF1 for ubiquitin-mediated degradation and regulates telomere maintenance. J. Boil. Chem. 2006, 281, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Her, Y.R.; Chung, I.K. Ubiquitin ligase RLIM modulates telomere length homeostasis through a proteolysis of TRF1. J. Boil. Chem. 2009, 284, 8557–8566. [Google Scholar] [CrossRef] [PubMed]
- Kulak, O.; Chen, H.; Holohan, B.; Wu, X.; He, H.; Borek, D.; Otwinowski, Z.; Yamaguchi, K.; Garofalo, L.A.; Ma, Z.; et al. Disruption of wnt/beta-catenin signaling and telomeric shortening are inextricable consequences of tankyrase inhibition in human cells. Mol. Cell. Biol. 2015, 35, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Donigian, J.R.; de Lange, T. The role of the poly(ADP-ribose) polymerase tankyrase1 in telomere length control by the TRF1 component of the shelterin complex. J. Boil. Chem. 2007, 282, 22662–22667. [Google Scholar] [CrossRef] [PubMed]
- Seimiya, H.; Muramatsu, Y.; Ohishi, T.; Tsuruo, T. Tankyrase 1 as a target for telomere-directed molecular cancer therapeutics. Cancer Cell 2005, 7, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Riffell, J.L.; Lord, C.J.; Ashworth, A. Tankyrase-targeted therapeutics: Expanding opportunities in the parp family. Nat. Rev. Drug Discov. 2012, 11, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Oshima, Y.; Yamamoto, Y.; Uochi, T.A.; Kusaka, H.; Akinaga, S.; Yamashita, Y.; Pongracz, K.; Pruzan, R.; Wunder, E.; et al. A novel telomerase template antagonist (GRN163) as a potential anticancer agent. Cancer Res. 2003, 63, 3931–3939. [Google Scholar] [PubMed]
- Armanios, M.; Greider, C.W. Treating myeloproliferation—On target or off? N. Engl. J. Med. 2015, 373, 965–966. [Google Scholar] [CrossRef] [PubMed]
- Chiappori, A.A.; Kolevska, T.; Spigel, D.R.; Hager, S.; Rarick, M.; Gadgeel, S.; Blais, N.; Von Pawel, J.; Hart, L.; Reck, M.; et al. A randomized phase II study of the telomerase inhibitor imetelstat as maintenance therapy for advanced non-small-cell lung cancer. Ann. Oncol. 2015, 26, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Baerlocher, G.M.; Oppliger Leibundgut, E.; Ottmann, O.G.; Spitzer, G.; Odenike, O.; McDevitt, M.A.; Roth, A.; Daskalakis, M.; Burington, B.; Stuart, M.; et al. Telomerase inhibitor imetelstat in patients with essential thrombocythemia. N. Engl. J. Med. 2015, 373, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Tefferi, A.; Lasho, T.L.; Begna, K.H.; Patnaik, M.M.; Zblewski, D.L.; Finke, C.M.; Laborde, R.R.; Wassie, E.; Schimek, L.; Hanson, C.A.; et al. A pilot study of the telomerase inhibitor imetelstat for myelofibrosis. N. Engl. J. Med. 2015, 373, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Ratain, M.; Benedetti, F.; Janisch, L.; Khor, S.; Schilsky, R.; Cohen, E.; Maitland, M.; Elias, L.; LoRusso, P. A phase I trial of GRN163L (GRN), a first-in-class telomerase inhibitor, in advanced solid tumors. In Proceedings of ASCO Annual Meeting, Chicago, IL, USA, May 2008; p. 3581.
- Kozloff, M.; Sledge, G.; Benedetti, F.; Starr, A.; Wallace, J.; Stuart, M.; Gruver, D.; Miller, K. Phase I study of imetelstat (GRN163L) in combination with paclitaxel (P) and bevacizumab (B) in patients (pts) with locally recurrent or metastatic breast cancer (MBC). In Proceedings of 2010 ASCO Annual Meeting, Chicago, IL, USA, June 2010; p. 2598.
- Begna, K.; Laborde, R.R.; Patnaik, M.M.; Lasho, T.L.; Zblewski, D.; Finke, C.; Schimek, L.; LaPlant, B.R.; Hanson, C.A.; Stuart, M.J. Imetelstat, a telomerase inhibitor, induces morphologic and molecular remissions in myelofibrosis and reversal of bone marrow fibrosis. Blood 2013, 122, 662–662. [Google Scholar]
- Tefferi, A.; LaPlant, B.R.; Begna, K.; Patnaik, M.M.; Lasho, T.L.; Zblewski, D.; Finke, C.; Schimek, L.; Pettinger, A.; Hanson, C.A. Imetelstat, a telomerase inhibitor, therapy for myelofibrosis: A pilot study. Blood 2014, 124, 634–634. [Google Scholar]
- Nemunaitis, J.; Tong, A.W.; Nemunaitis, M.; Senzer, N.; Phadke, A.P.; Bedell, C.; Adams, N.; Zhang, Y.A.; Maples, P.B.; Chen, S.; et al. A phase I study of telomerase-specific replication competent oncolytic adenovirus (telomelysin) for various solid tumors. Mol. Ther. 2010, 18, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Greten, T.F.; Forner, A.; Korangy, F.; N’Kontchou, G.; Barget, N.; Ayuso, C.; Ormandy, L.A.; Manns, M.P.; Beaugrand, M.; Bruix, J. A phase II open label trial evaluating safety and efficacy of a telomerase peptide vaccination in patients with advanced hepatocellular carcinoma. BMC Cancer 2010. [Google Scholar] [CrossRef] [PubMed]
- Brunsvig, P.F.; Aamdal, S.; Gjertsen, M.K.; Kvalheim, G.; Markowski-Grimsrud, C.J.; Sve, I.; Dyrhaug, M.; Trachsel, S.; Møller, M.; Eriksen, J.A. Telomerase peptide vaccination: A phase I/II study in patients with non-small cell lung cancer. Cancer Immunol. Immunother. 2006, 55, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Aamdal, S.; Dueland, S.; Engebraaten, O.; Owre, K.; Dyrhaug, M.; Trachsel, S.; Gaudernack, G. A phase I/II study of telomerase peptide vaccination in combination with chemotherapy in patients with stage IV malignant melanoma. In Proceedings of ASCO Annual Meeting, San Francisco, CA, USA, January 2006; p. 8031.
- Inderberg-Suso, E.M.; Trachsel, S.; Lislerud, K.; Rasmussen, A.M.; Gaudernack, G. Widespread CD4+ T-cell reactivity to novel hTERT epitopes following vaccination of cancer patients with a single hTERT peptide gv1001. Oncoimmunology 2012, 1, 670–686. [Google Scholar] [CrossRef] [PubMed]
- Buanes, T.; Maurel, J.; Liauw, W.; Hebbar, M.; Nemunaitis, J. A randomized phase III study of gemcitabine (G) versus GV1001 in sequential combination with G in patients with unresectable and metastatic pancreatic cancer (PC). In Proceedings of ASCO Annual Meeting, Orlando, FL, USA, May 2009; p. 4601.
- Georgoulias, V.; Douillard, J.Y.; Khayat, D.; Manegold, C.; Rosell, R.; Rossi, A.; Menez-Jamet, J.; Iche, M.; Kosmatopoulos, K.; Gridelli, C. A multicenter randomized phase IIB efficacy study of VX-001, a peptide-based cancer vaccine as maintenance treatment in advanced non-small-cell lung cancer: Treatment rationale and protocol dynamics. Clin. Lung Cancer 2013, 14, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Parkhurst, M.R.; Riley, J.P.; Igarashi, T.; Li, Y.; Robbins, P.F.; Rosenberg, S.A. Immunization of patients with the hTERT: 540–548 peptide induces peptide-reactive T lymphocytes that do not recognize tumors endogenously expressing telomerase. Clin. Cancer Res. 2004, 10, 4688–4698. [Google Scholar] [CrossRef] [PubMed]
- Domchek, S.M.; Recio, A.; Mick, R.; Clark, C.E.; Carpenter, E.L.; Fox, K.R.; DeMichele, A.; Schuchter, L.M.; Leibowitz, M.S.; Wexler, M.H.; et al. Telomerase-specific T-cell immunity in breast cancer: Effect of vaccination on tumor immunosurveillance. Cancer Res. 2007, 67, 10546–10555. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.P.; Aqui, N.A.; Stadtmauer, E.A.; Vogl, D.T.; Fang, H.B.; Cai, L.; Janofsky, S.; Chew, A.; Storek, J.; Akpek, G. Combination immunotherapy using adoptive T-cell transfer and tumor antigen vaccination on the basis of hTERT and survivin after asct for myeloma. Blood 2011, 117, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Rech, A.; Mick, R.; Recio, A.; DeMichele, A.; Tweed, C.; Fox, K.; Domchek, S.; Vonderheide, R. Phase I study of anti-CD25 mab daclizumab to deplete regulatory T cells prior to telomerase/survivin peptide vaccination in patients (pts) with metastatic breast cancer (MBC). In Proceedings of ASCO Annual Meeting, Chicago, IL, USA, June 2010; p. 2508.
- Khoury, H.J.; Collins, R.; Blum, W.G.; Stiff, P.J.; Lebkowski, J.; Wirth, E.; Nishimoto, K.; DiPersio, J.F. Long-term follow-up of patients with acute myelogenous leukemia receiving an autologous telomerase-based dendritic cell vaccine. In Proceedings of 2015 ASCO Annual Meeting, Chicago, IL, USA, May 2015.
- Vonderheide, R.H.; Domchek, S.M.; Schultze, J.L.; George, D.J.; Hoar, K.M.; Chen, D.Y.; Stephans, K.F.; Masutomi, K.; Loda, M.; Xia, Z. Vaccination of cancer patients against telomerase induces functional antitumor CD8+ T lymphocytes. Clin. Cancer Res. 2004, 10, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Trepiakas, R.; Berntsen, A.; Hadrup, S.R.; Bjørn, J.; Geertsen, P.F.; Straten, P.T.; Andersen, M.H.; Pedersen, A.E.; Soleimani, A.; Lorentzen, T. Vaccination with autologous dendritic cells pulsed with multiple tumor antigens for treatment of patients with malignant melanoma: Results from a phase I/II trial. Cytotherapy 2010, 12, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Berntsen, A.; Geertsen, P.; Trepiakas, R.; Wenandy, L.; Andersen, M.; Straten, P.; Lorentzen, T.; Johansen, J.; Johnsen, H.; Svane, I. Dendritic cell based vaccination in combination with IL-2 as a treatment for advanced renal cell carcinoma patients: Results from a phase I/II trial. In Proceedings of ASCO Annual Meeting, San Francisco, CA, USA, January 2006; p. 2575.
- Su, Z.; Dannull, J.; Yang, B.K.; Dahm, P.; Coleman, D.; Yancey, D.; Sichi, S.; Niedzwiecki, D.; Boczkowski, D.; Gilboa, E. Telomerase mrna-transfected dendritic cells stimulate antigen-specific CD8+ and CD4+ T cell responses in patients with metastatic prostate cancer. J. Immunol. 2005, 174, 3798–3807. [Google Scholar] [CrossRef] [PubMed]
- Millard, F.; Gerloni, M.; Darrah, D.; Farness, P.; Zanetti, M. Phase I study of transgenic B lymphocyte immunization (TLI) against telomerase in androgen-independent prostate cancer (PC). In Proceedings of ASCO Annual Meeting, New Orleans, LA, USA, June 2004; p. 2519.
- Yu, G.L.; Bradley, J.D.; Attardi, L.D.; Blackburn, E.H. In vivo alteration of telomere sequences and senescence caused by mutated tetrahymena telomerase RNAs. Nature 1990, 344, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Rosenberg, J.E.; Donjacour, A.A.; Botchkina, I.L.; Hom, Y.K.; Cunha, G.R.; Blackburn, E.H. Rapid inhibition of cancer cell growth induced by lentiviral delivery and expression of mutant-template telomerase RNA and anti-telomerase short-interfering RNA. Cancer Res. 2004, 64, 4833–4840. [Google Scholar] [CrossRef] [PubMed]
- Goldkorn, A.; Blackburn, E.H. Assembly of mutant-template telomerase RNA into catalytically active telomerase ribonucleoprotein that can act on telomeres is required for apoptosis and cell cycle arrest in human cancer cells. Cancer Res. 2006, 66, 5763–5771. [Google Scholar] [CrossRef] [PubMed]
- Stohr, B.A.; Blackburn, E.H. ATM mediates cytotoxicity of a mutant telomerase RNA in human cancer cells. Cancer Res. 2008, 68, 5309–5317. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Tay, L.L.; Tan, W.H.; Chai, J.H.; Wang, X. Mutant telomerase RNAS induce DNA damage and apoptosis via the TRF2-ATM pathway in telomerase-overexpressing primary fibroblasts. FEBS J. 2011, 278, 3724–3738. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Xu, Y.; Liao, C.P.; Lau, R.; Goldkorn, A. Reprogramming murine telomerase rapidly inhibits the growth of mouse cancer cells in vitro and in vivo. Mol. Cancer Ther. 2010, 9, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.Y. Non-canonical NF-κB signalling and ets1/2 cooperatively drive c250t mutant tert promoter activation. Nat. Cell Boil. 2015, 17, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Kagawa, S.; Kobayashi, N.; Shirakiya, Y.; Umeoka, T.; Teraishi, F.; Taki, M.; Kyo, S.; Tanaka, N.; Fujiwara, T. Telomerase-specific replication-selective virotherapy for human cancer. Clin. Cancer Res. 2004, 10, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hioki, M.; Fujiwara, T.; Nishizaki, M.; Kagawa, S.; Taki, M.; Kishimoto, H.; Endo, Y.; Urata, Y.; Tanaka, N.; et al. Histone deacetylase inhibitor FR901228 enhances the antitumor effect of telomerase-specific replication-selective adenoviral agent OBP-301 in human lung cancer cells. Exp. Cell Res. 2006, 312, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Tazawa, H.; Hashimoto, Y.; Shirakawa, Y.; Kuroda, S.; Nishizaki, M.; Kishimoto, H.; Uno, F.; Nagasaka, T.; Urata, Y.; et al. A genetically engineered oncolytic adenovirus decoys and lethally traps quiescent cancer stem-like cells in S/G2/M phases. Clin. Cancer Res. 2013, 19, 6495–6505. [Google Scholar] [CrossRef] [PubMed]
- Sakai, R.; Kagawa, S.; Yamasaki, Y.; Kojima, T.; Uno, F.; Hashimoto, Y.; Watanabe, Y.; Urata, Y.; Tanaka, N.; Fujiwara, T. Preclinical evaluation of differentially targeting dual virotherapy for human solid cancer. Mol. Cancer Ther. 2010, 9, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Bilsland, A.E.; Anderson, C.J.; Fletcher-Monaghan, A.J.; McGregor, F.; Evans, T.R.; Ganly, I.; Knox, R.J.; Plumb, J.A.; Keith, W.N. Selective ablation of human cancer cells by telomerase-specific adenoviral suicide gene therapy vectors expressing bacterial nitroreductase. Oncogene 2003, 22, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.A.; Bilsland, A.; Kakani, R.; Zhao, J.; Glasspool, R.M.; Knox, R.J.; Evans, T.R.; Keith, W.N. Telomerase-specific suicide gene therapy vectors expressing bacterial nitroreductase sensitize human cancer cells to the pro-drug CB1954. Oncogene 2001, 20, 7797–7803. [Google Scholar] [CrossRef] [PubMed]
- Schepelmann, S.; Ogilvie, L.M.; Hedley, D.; Friedlos, F.; Martin, J.; Scanlon, I.; Chen, P.; Marais, R.; Springer, C.J. Suicide gene therapy of human colon carcinoma xenografts using an armed oncolytic adenovirus expressing carboxypeptidase G2. Cancer Res. 2007, 67, 4949–4955. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.T.; Li, C.; Lu, M.H.; Liang, G.P.; Li, N.; Tang, X.D.; Wu, Y.Y.; Shi, C.M.; Chen, L.; Li, C.Z.; et al. Noninvasive and real-time monitoring of the therapeutic response of tumors in vivo with an optimized hTERT promoter. Cancer 2012, 118, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Low, K.C.; Liu, C.Y.; Yan, T.D.; Oji, A.; Ikawa, M.; Li, S.; Tergaonkar, V. Quantitative assessment of telomerase components in cancer cell lines. FEBS Lett. 2015, 589, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Vonderheide, R.H.; Hahn, W.C.; Schultze, J.L.; Nadler, L.M. The telomerase catalytic subunit is a widely expressed tumor-associated antigen recognized by cytotoxic T lymphocytes. Immunity 1999, 10, 673–679. [Google Scholar] [CrossRef]
- Kokhaei, P.; Palma, M.; Hansson, L.; Osterborg, A.; Mellstedt, H.; Choudhury, A. Telomerase (hTERT 611–626) serves as a tumor antigen in B-cell chronic lymphocytic leukemia and generates spontaneously antileukemic, cytotoxic T cells. Exp. Hematol. 2007, 35, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Brunsvig, P.F.; Kyte, J.A.; Kersten, C.; Sundstrom, S.; Moller, M.; Nyakas, M.; Hansen, G.L.; Gaudernack, G.; Aamdal, S. Telomerase peptide vaccination in nsclc: A phase II trial in stage III patients vaccinated after chemoradiotherapy and an 8-year update on a phase I/II trial. Clin. Cancer Res. 2011, 17, 6847–6857. [Google Scholar] [CrossRef] [PubMed]
- Vetsika, E.K.; Papadimitraki, E.; Aggouraki, D.; Konsolakis, G.; Mela, M.E.; Kotsakis, A.; Christou, S.; Patramani, S.; Alefantinou, M.; Kaskara, A.; et al. Sequential administration of the native TERT572 cryptic peptide enhances the immune response initiated by its optimized variant TERT(572Y) in cancer patients. J. Immunother. 2011, 34, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, D.; Bolonakis, I.; Cornet, S.; Myllaki, G.; Kanellou, P.; Kotsakis, A.; Galanis, A.; Nikoloudi, I.; Spyropoulou, M.; Menez, J.; et al. A phase I study of the optimized cryptic peptide TERT(572Y) in patients with advanced malignancies. Oncology 2006, 70, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Bolonaki, I.; Kotsakis, A.; Papadimitraki, E.; Aggouraki, D.; Konsolakis, G.; Vagia, A.; Christophylakis, C.; Nikoloudi, I.; Magganas, E.; Galanis, A.; et al. Vaccination of patients with advanced non-small-cell lung cancer with an optimized cryptic human telomerase reverse transcriptase peptide. J. Clin. Oncol. 2007, 25, 2727–2734. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, D.; Parodi, A.; Lavieri, R.; Kalli, F.; Ferrera, F.; Tagliamacco, A.; Guastalla, A.; Lamperti, M.G.; Giacomini, M.; Filaci, G. Immunogenicity of GX301 cancer vaccine: Four (telomerase peptides) are better than one. Hum. Vaccine. Immunother. 2015, 11, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, D.; Traverso, P.; Parodi, A.; Tomasello, L.; Negrini, S.; Kalli, F.; Battaglia, F.; Ferrera, F.; Sciallero, S.; Murdaca, G.; et al. A multi-peptide, dual-adjuvant telomerase vaccine (GX301) is highly immunogenic in patients with prostate and renal cancer. Cancer Immunol. Immunother. 2013, 62, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Maker, A.V.; Attia, P.; Rosenberg, S.A. Analysis of the cellular mechanism of antitumor responses and autoimmunity in patients treated with CTLA-4 blockade. J. Immunol. 2005, 175, 7746–7754. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.C.; Rucki, A.A.; Wu, A.A.; Olino, K.; Xiao, Q.; Chai, Y.; Wamwea, A.; Bigelow, E.; Lutz, E.; Liu, L.; et al. PD-1/PD-L1 blockade together with vaccine therapy facilitates effector T-cell infiltration into pancreatic tumors. J. Immunol. 2015, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Perrem, K.; Colgin, L.M.; Neumann, A.A.; Yeager, T.R.; Reddel, R.R. Coexistence of alternative lengthening of telomeres and telomerase in hTERT-transfected GM847 cells. Mol. Cell. Boil. 2001, 21, 3862–3875. [Google Scholar] [CrossRef] [PubMed]
- Cerone, M.A.; Londono-Vallejo, J.A.; Bacchetti, S. Telomere maintenance by telomerase and by recombination can coexist in human cells. Hum. Mol. Genet. 2001, 10, 1945–1952. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Agent | NCT Identifier | Trial | Status | Results |
|---|---|---|---|---|
| Telomerase Inhibitor | ||||
| Imetelstat | NCT01256762 | Ph II: Breast cancer | Completed | N/A |
| Imetelstat [129] | NCT01137968 | Ph II: NSCLC | Completed | Failed to improve PFS; Grade 3/4 neutropenia and thrombocytopenia |
| Imetelstat | NCT00124189 | Ph I: CLD | Completed | N/A |
| Imetelstat | NCT00510445 | Ph I: Lung cancer | Completed | N/A |
| Imetelstat | NCT00718601 | Ph I: Melanoma | Completed | N/A |
| Imetelstat [132] | NCT00310895 | Ph I: Solid tumor | Completed | Thrombocytopenia at doses >3.2 mg/kg/wk |
| Imetelstat | NCT01265927 | Ph I: Breast neoplasms | Completed | N/A |
| Imetelstat [133] | NCT00732056 | Ph I: Breast cancer | Completed | No DLTs, Cytopenias |
| Imetelstat [130] | NCT01243073 | Ph II: ET | Completed | Rapid and durable hematologic and molecular responses |
| Imetelstat | NCT00594126 | Ph I: Myeloma | Completed | N/A |
| Imetelstat | NCT01242930 | Ph II: Myeloma | Completed | N/A |
| Imetelstat | NCT01273090 | Ph I: Solid tumor or Lymphoma | Completed | N/A |
| Imetelstat | NCT01836549 | Ph II: Brain tumor | Active, not recruiting | N/A |
| Imetelstat [131,134,135] | NCT01731951 | Ph II: Myelofibrosis | Active, not recruiting | Complete or partial remissions with various toxicity |
| Imetelstat | NCT02598661 | Ph III: MDS | Recruiting | N/A |
| Imetelstat | NCT02426086 | Ph II: Myelofibrosis | Recruiting | N/A |
| Oncolytic Virus | ||||
| Telomelysin/OBP-301 [136] | NCT02293850 | Ph I/II: HCC | Recruiting | Pain at injection site, fevers, chills; detected viral DNA in 13/16 pts |
| Telomerase Vaccine | ||||
| GV1001 | NCT01223209 | Ph I: Carcinoma | Recruiting | N/A |
| GV1001 [137] | NCT00444782 | Ph II: HCC | Completed | Well tolerated, mild injection site reaction; No antitumor immune response |
| GV1001 [138] | NCT00509457 | Ph I/II: NSCLC | Completed | Minor side effects, no bone marrow toxicity; Immune response in 13/24 pts |
| GV1001 [139,140] | NCT01247623 | Ph I/II: Melanoma | Completed | Well tolerated with neutropenia in 1/14 pts; Immune response in 17/21 pts; highly diverse TERT-specific T-cell response |
| GV1001 [141] | NCT00358566 | Ph III: Pancreatic cancer | Terminated | No survival benefit |
| GV1001 | NCT00425360 | Ph III: Pancreatic cancer | Completed | N/A |
| GV1001 | NCT01342224 | Ph I: Pancreatic adenocarcinoma | Active, not recruiting | N/A |
| GV1001 | NCT01579188 | Ph III: NSCLC | Not yet recruiting | N/A |
| VX-001 [142] | NCT01935154 | Ph II: NSCLC | Active, not recruiting | |
| GX-301 | NCT02293707 | Ph II: Prostate cancer | Recruiting | N/A |
| hTERT 540–548 peptide [143] | NCT00021164 | Ph II: Melanoma, Solid tumor | Completed | No immune response against hTERT + tumor |
| hTERT 540–548 peptide | NCT00069940 | Ph I: Brain tumor, Sarcoma | Completed | N/A |
| hTERT 540–548 peptide [144] | NCT00079157 | Ph I: Breast cancer | N/A | Injection site related erythema, pain, pruritus, or induration; Prolonged survival |
| hTERT multi peptide [145] | NCT00499577 | Ph I/II: Myloma, plasma cell neoplasm | Completed | Mild to moderate chills and rigors; Antitumor immunity in 10/28 pts |
| hTERT multi peptide [146] | NCT00573495 | Ph I: Breast cancer | Completed | Well tolerated; Immune response in 80% pts |
| hTERT multi peptide | NCT00834665 | Ph I/II: Myeloma | N/A | N/A |
| GRNVAC1/AST-VAC1 [147] | NCT00510133 | Ph II: AML | Completed | TERT specific T cell response in 11/19 pts; 57% pts in complete remission at follow-up 52 months |
| AST-VAC2 | N/A | Ph I/II: NSCLC | N/A | N/A |
| DC pulsed with hTERT 540–548 peptide [148] | N/A | Ph I: Breast and Prostate cancer | N/A | Well tolerated, no changes in B cells number; Immune response in 4/7 pts |
| DC pulsed with Telomerase peptide or tumor lysates [149] | NCT00197912 | Ph I/II: Melanoma | Completed | Feasible and safe: Prolonged survival |
| DC pulsed with Telomerase peptide or tumor lysates [150] | NCT00197860 | Ph I/II: RCC | Completed | Well tolerated without severe toxicities; Disease stabilized in half pts |
| DC pulsed with hTERT mRNA [151] | NCT01153113 | Ph I/II: Prostate cancer | Withdrawn | Fatigue or flu-like symptoms, erythema/induration; Immune response in 19/20 pts |
| TLI (hTERT DNA fragment) [152] | NCT00061035 | Ph I: Prostate cancer | Completed | Feasible and safe: Immune response by single-dose TLI |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Goldkorn, A. Telomere and Telomerase Therapeutics in Cancer. Genes 2016, 7, 22. https://doi.org/10.3390/genes7060022
Xu Y, Goldkorn A. Telomere and Telomerase Therapeutics in Cancer. Genes. 2016; 7(6):22. https://doi.org/10.3390/genes7060022
Chicago/Turabian StyleXu, Yucheng, and Amir Goldkorn. 2016. "Telomere and Telomerase Therapeutics in Cancer" Genes 7, no. 6: 22. https://doi.org/10.3390/genes7060022
APA StyleXu, Y., & Goldkorn, A. (2016). Telomere and Telomerase Therapeutics in Cancer. Genes, 7(6), 22. https://doi.org/10.3390/genes7060022