Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications—A Twin Study

 ,
,
Abstract
:1. Introduction
2. Experimental

{kind=link}
| All | MZ | DZ | P | |
|---|---|---|---|---|
| n (men/women) | 244 (110/134) | 96 (51/44) | 148 (59/90) | - |
| n (in pairs/single twins) | 156/88 | 66/30 | 90/58 | - |
| Age (years) | 73.4 ± 5.2 | 73.8 ± 4.9 | 73.2 ± 5.4 | 0.36 |
| BMI (kg/m2) | 26.0 ± 3.7 | 26.0 ± 3.2 | 26.1 ± 4.0 | 0.97 |
| Birth weight (g) | 2641.0 ± 446.7 | 2608.6 ± 469.7 | 2659.8 ± 431.9 | 0.50 |
| 2 h OGTT glucose (mmol/L) | 8.3 ± 3.8 | 8.0 ± 3.5 | 8.5 ± 4.0 | 0.37 |
| Triglycerides (mmol/L) | 1.3 ± 0.6 | 1.4 ± 0.7 | 1.2 ± 0.5 | 0.02 |
| HOMA-IR | 1.8 ± 1.3 | 1.9 ± 1.7 | 1.8 ± 1.1 | 0.53 |
| HbA1c (%) | 5.8 ± 0.7 | 5.8 ± 0.6 | 5.8 ± 0.7 | 0.89 |
| miR-103 (RQ) | 2.1 ± 1.8 | 1.8 ± 1.7 | 2.3 ± 1.9 | 0.03 |
| miR-143 (RQ) | 3.2 ± 2.1 | 3.4 ± 2.3 | 3.2 ± 2.0 | 0.54 |
| miR-483-3p (RQ) | 2.5 ± 1.9 | 2.5 ± 1.8 | 2.5 ± 1.9 | 0.88 |
| n (diabetes) | 31 (13%) | 9 (9%) | 22 (15%) | - |
| n (IGT) | 72 (30%) | 32 (33%) | 41 (28%) | - |
| n (NGT) | 139 (57%) | 54 (56%) | 85 (57%) | - |
2.1. Clinical Examination and Tissue Sampling
2.2. miRNA Expression Analysis
2.3. Statistical Methods
3. Results and Discussion
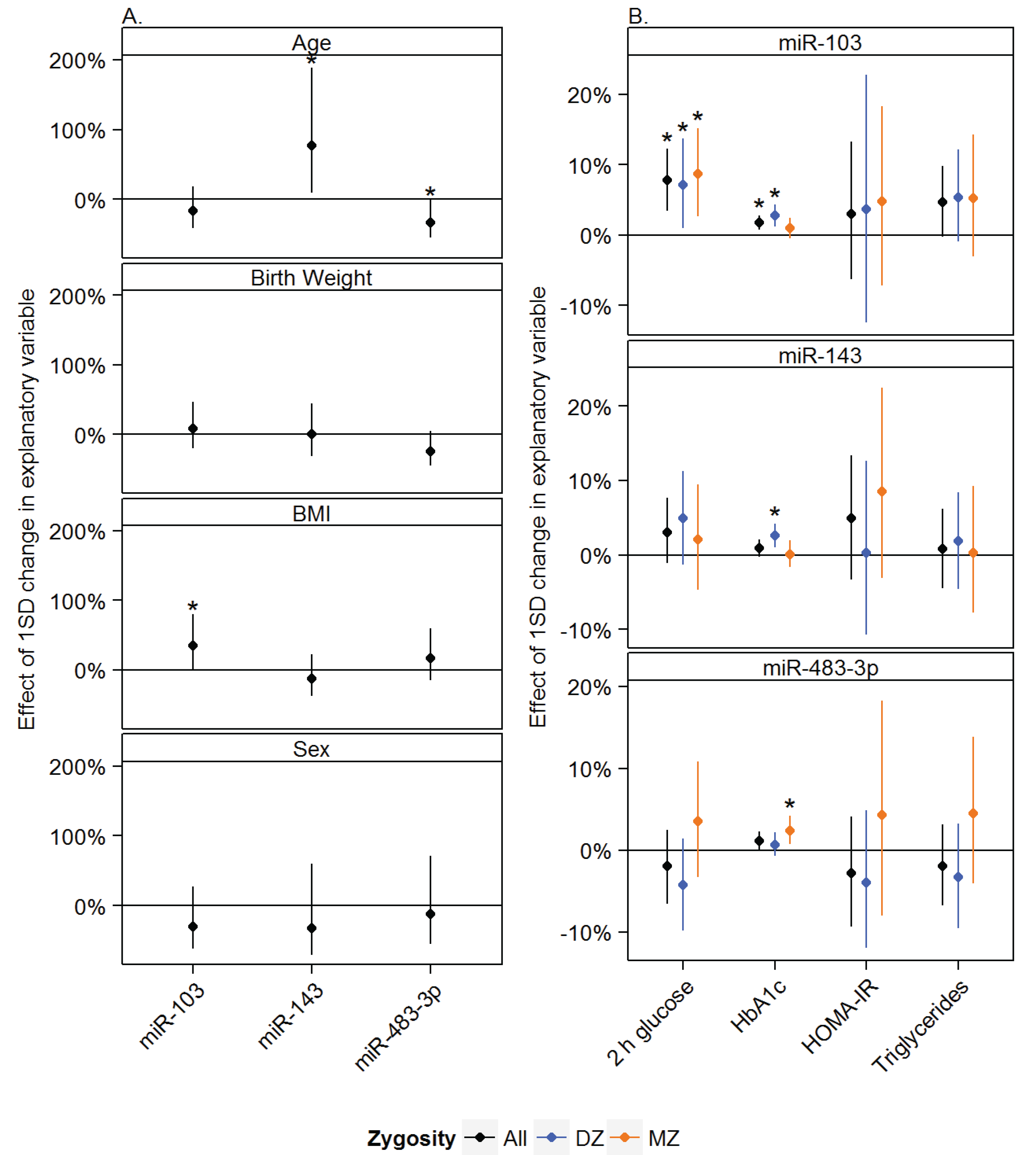
3.1. Age, Sex and Anthropometric Measurements Associated with miRNA Expression in Adipose Tissue
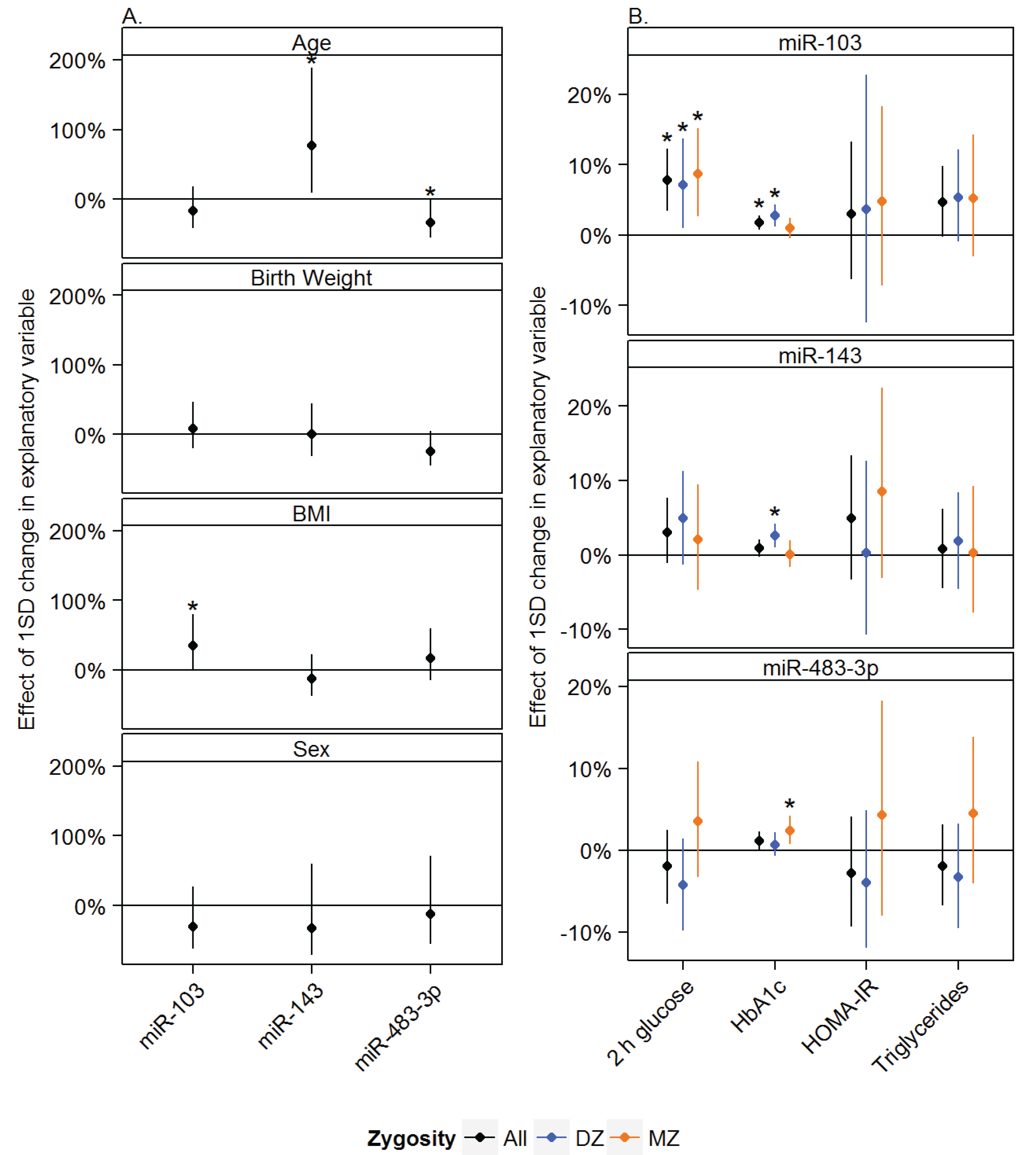
3.2. Associations between miRNAs and Measures of Glucose and Lipid Metabolism
3.3. Effects of Zygosity
3.4. Heritability Analysis
3.5. Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar]
- Xie, H.; Lim, B.; Lodish, H.F. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef]
- Takanabe, R.; Ono, K.; Abe, Y.; Takaya, T.; Horie, T.; Wada, H.; Kita, T.; Satoh, N.; Shimatsu, A.; Hasegawa, K. Up-regulated expression of microRNA-143 in association with obesity in adipose tissue of mice fed high-fat diet. Biochem. Biophys. Res. Commun. 2008, 376, 728–732. [Google Scholar] [CrossRef]
- Jordan, S.D.; Kruger, M.; Willmes, D.M.; Redemann, N.; Wunderlich, F.T.; Bronneke, H.S.; Merkwirth, C.; Kashkar, H.; Olkkonen, V.M.; Bottger, T.; et al. Obesity-induced overexpression of miRNA-143 inhibits insulin-stimulated AKT activation and impairs glucose metabolism. Nat. Cell Biol. 2011, 13, 434–446. [Google Scholar]
- Duan, R.; Pak, C.; Jin, P. Single nucleotide polymorphism associated with mature miR-125a alters the processing of pri-miRNA. Hum. Mol. Genet. 2007, 16, 1124–1131. [Google Scholar] [CrossRef]
- Sun, T.; Fu, M.; Bookout, A.L.; Kliewer, S.A.; Mangelsdorf, D.J. MicroRNA let-7 regulates 3T3-L1 adipogenesis. Mol. Endocrinol. 2009, 23, 925–931. [Google Scholar] [CrossRef]
- Ferland-McCollough, D.; Fernandez-Twinn, D.S.; Cannell, I.G.; David, H.; Warner, M.; Vaag, A.A.; Bork-Jensen, J.; Brons, C.; Gant, T.W.; Willis, A.E.; et al. Programming of adipose tissue miR-483–3p and GDF-3 expression by maternal diet in type 2 diabetes. Cell Death Differ. 2012, 19, 1003–1012. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar]
- Vaag, A.; Jensen, C.B.; Poulsen, P.; Brons, C.; Pilgaard, K.; Grunnet, L.; Vielwerth, S.; Alibegovic, A. Metabolic aspects of insulin resistance in individuals born small for gestational age. Horm. Res. Paediatr. 2006, 65, 137–143. [Google Scholar] [CrossRef]
- Poulsen, P.; Ohm Kyvik, K.; Vaag, A.; Beck-Nielsen, H. Heritability of type II (non-insulin-dependent) diabetes mellitus and abnormal glucose tolerance—A population-based twin study. Diabetologia 1999, 42, 139–145. [Google Scholar] [CrossRef]
- Grunnet, L.G.; Nilsson, E.; Ling, C.; Hansen, T.; Pedersen, O.; Groop, L.; Vaag, A.; Poulsen, P. Regulation and function of FTO mRNA expression in human skeletal muscle and subcutaneous adipose tissue. Diabetes 2009, 58, 2402–2408. [Google Scholar] [CrossRef]
- Poulsen, P.; Grunnet, L.G.; Pilgaard, K.; Storgaard, H.; Alibegovic, A.; Sonne, M.P.; Carstensen, B.; Beck-Nielsen, H.; Vaag, A. Increased risk of type 2 diabetes in elderly twins. Diabetes 2009, 58, 1350–1355. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Bring, J.; Wernroth, L. Inefficient analysis of twin data: Is there an association between diabetes and birth weight? Diabetologia 1999, 42, 898–899. [Google Scholar] [CrossRef]
- Poulsen, P.; Vaag, A.; Beck-Nielsen, H. The influence of zygosity status on blood pressure and on lipid profiles in male and female twins. J. Hypertens. 2002, 20, 654–659. [Google Scholar]
- Ryan, B.M.; Robles, A.I.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389–402. [Google Scholar] [CrossRef]
- Mohan, S.; Farley, J.R.; Baylink, D.J. Age-related changes in IGFBP-4 and IGFBP-5 levels in human serum and bone: Implications for bone loss with aging. Prog. Growth Factor Res. 1995, 6, 465–473. [Google Scholar] [CrossRef]
- Garite, T.J.; Clark, R.H.; Elliott, J.P.; Thorp, J.A. The Pediatrix/Obstetrix Perinatal Research Group. Twins and triplets: The effect of plurality and growth on neonatal outcome compared with singleton infants. Am. J. Obstet. Gynecol. 2004, 191, 700–707. [Google Scholar]
- Ortega, F.J.; Moreno-Navarrete, J.M.; Pardo, G.; Sabater, M.; Hummel, M.; Ferrer, A.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. MiRNA expression profile of human subcutaneous adipose and during adipocyte differentiation. PLoS One 2010, 5, e9022. [Google Scholar]
- Klöting, N.; Berthold, S.; Kovacs, P.; Schön, M.R.; Fasshauer, M.; Ruschke, K.; Stumvoll, M.; Blüher, M. MicroRNA expression in human omental and subcutaneous adipose tissue. PLoS One 2009, 4, e4699. [Google Scholar]
- Yan, D.; Mäyränpää, M.I.; Wong, J.; Perttilä, J.; Lehto, M.; Jauhiainen, M.; Kovanen, P.T.; Ehnholm, C.; Brown, A.J.; Olkkonen, V.M.; et al. OSBP-related protein 8 (ORP8) suppresses ABCA1 expression and cholesterol efflux from macrophages. J. Biol. Chem. 2008, 283, 332–340. [Google Scholar] [CrossRef]
- Poulsen, P.; Levin, K.; Beck-Nielsen, H.; Vaag, A. Age-dependent impact of zygosity and birth weight on insulin secretion and insulin action in twins. Diabetologia 2002, 45, 1649–1657. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bork-Jensen, J.; Thuesen, A.C.B.; Bang-Bertelsen, C.H.; Grunnet, L.G.; Pociot, F.; Beck-Nielsen, H.; Ozanne, S.E.; Poulsen, P.; Vaag, A. Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications—A Twin Study. Genes 2014, 5, 508-517. https://doi.org/10.3390/genes5030508
Bork-Jensen J, Thuesen ACB, Bang-Bertelsen CH, Grunnet LG, Pociot F, Beck-Nielsen H, Ozanne SE, Poulsen P, Vaag A. Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications—A Twin Study. Genes. 2014; 5(3):508-517. https://doi.org/10.3390/genes5030508
Chicago/Turabian StyleBork-Jensen, Jette, Anne Cathrine Baun Thuesen, Claus Heiner Bang-Bertelsen, Louise Groth Grunnet, Flemming Pociot, Henning Beck-Nielsen, Susan E. Ozanne, Pernille Poulsen, and Allan Vaag. 2014. "Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications—A Twin Study" Genes 5, no. 3: 508-517. https://doi.org/10.3390/genes5030508
APA StyleBork-Jensen, J., Thuesen, A. C. B., Bang-Bertelsen, C. H., Grunnet, L. G., Pociot, F., Beck-Nielsen, H., Ozanne, S. E., Poulsen, P., & Vaag, A. (2014). Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications—A Twin Study. Genes, 5(3), 508-517. https://doi.org/10.3390/genes5030508