
Compositional and Functional Disparities in the Breast Oncobiome Between Patients Living in Urban or Rural Areas

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Enrolment
2.2. Characterization of Breast Tissue Microbiota
2.3. Statistical Analysis
3. Results
3.1. Patients
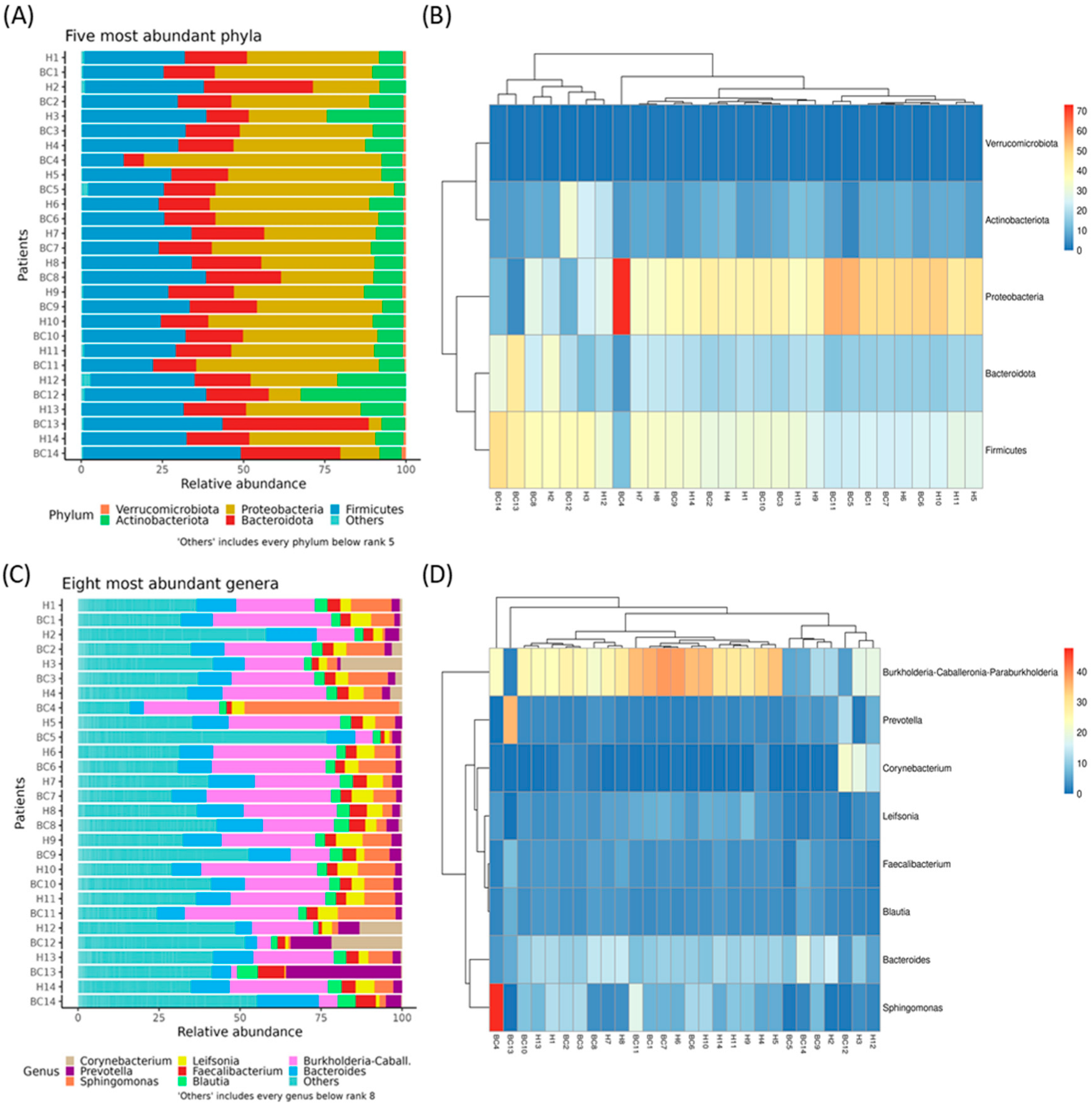
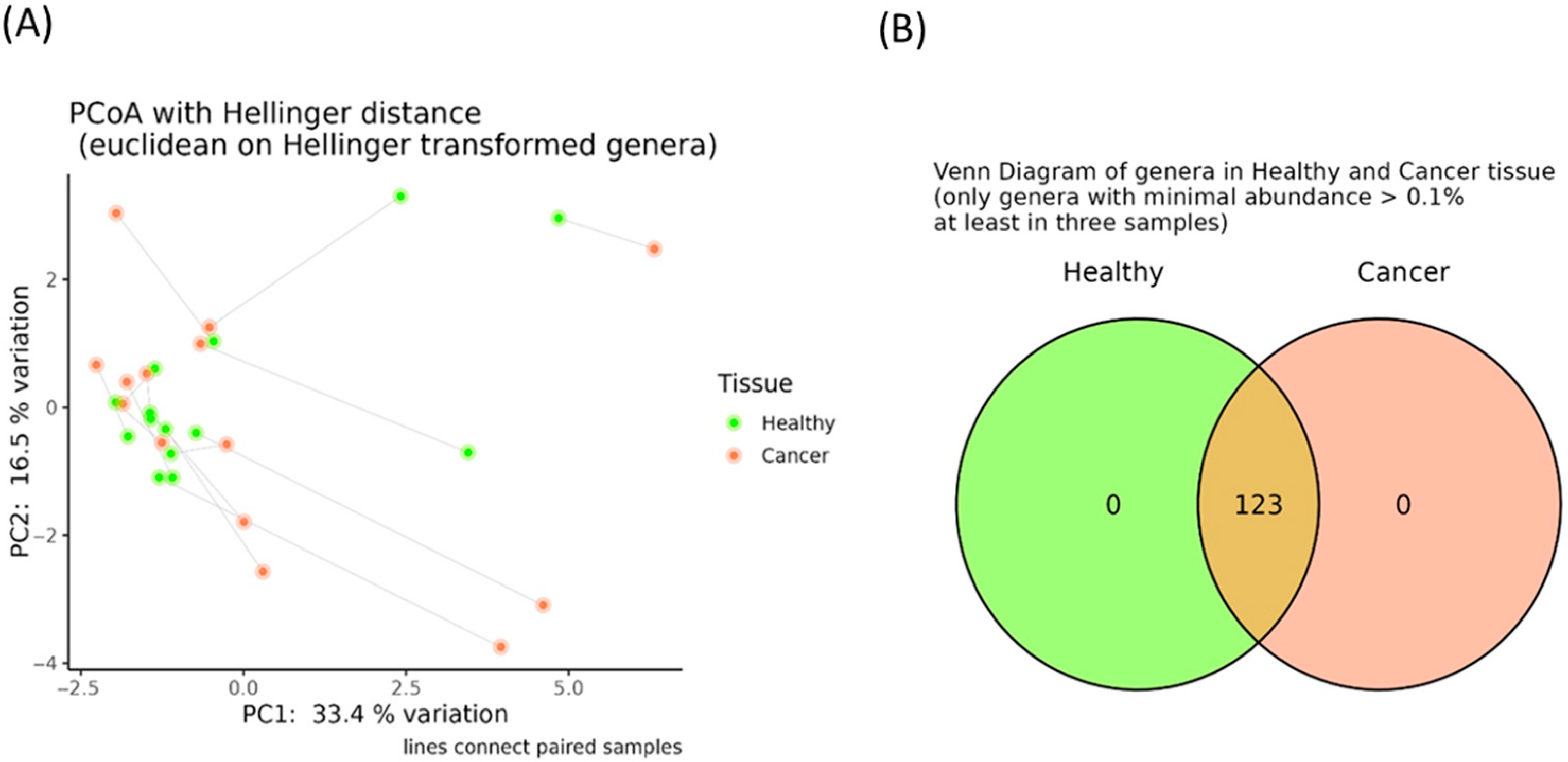
3.2. Evaluation of Breast Tissue-Associated Microbiota in Tumor and Adjacent Healthy Samples
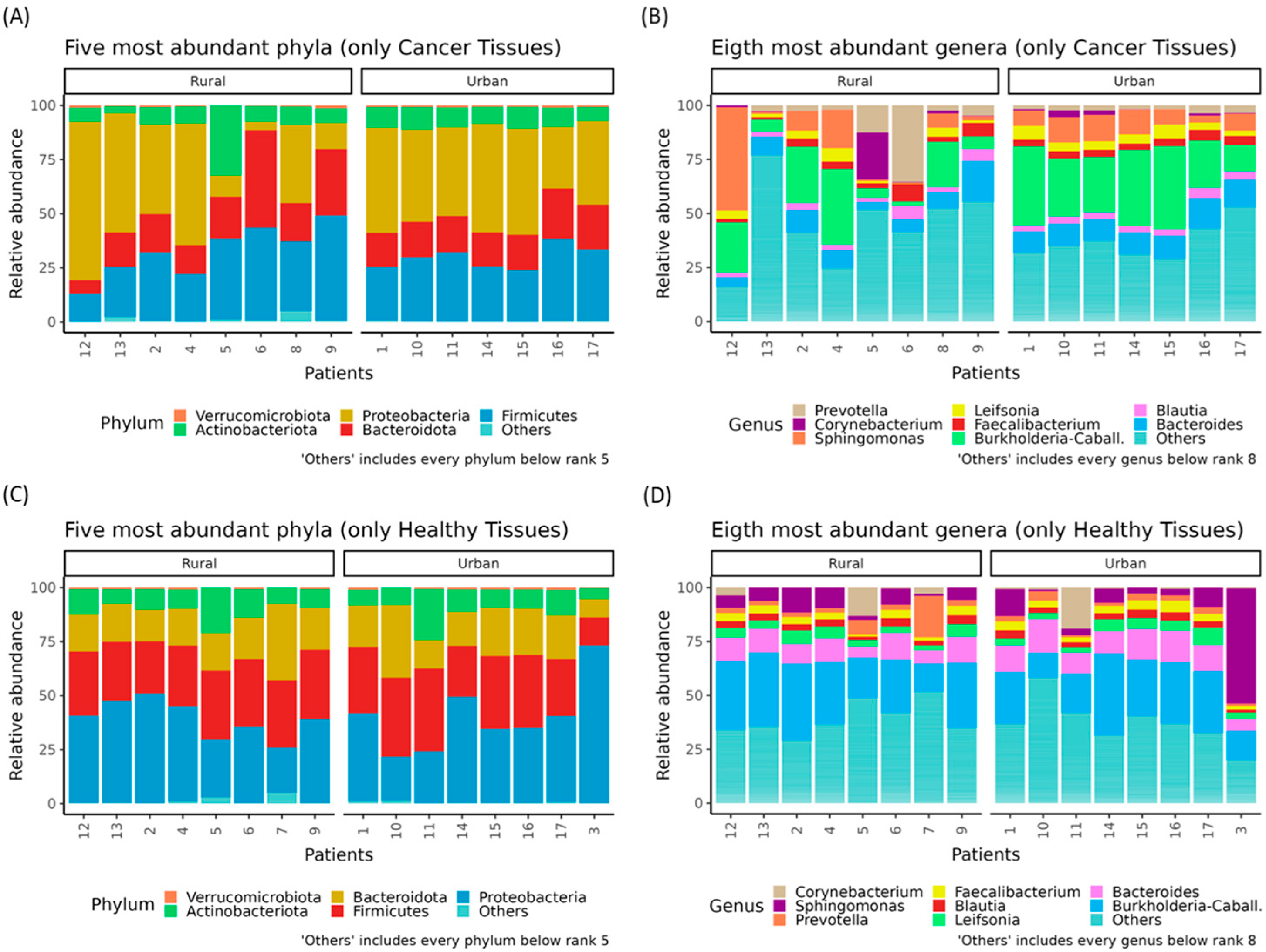
3.3. Rural–Urban Differences in Oncobiome and Paired Healthy Breast Tissues′ Microbiota
3.4. PICRUST Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [PubMed]
- Thun, M.; Linet, M.S.; Cerhan, J.R.; Haiman, C.A.; Schottenfeld, D. (Eds.) Cancer Epidemiology and Prevention; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Najjar, H.; Easson, A. Age at diagnosis of breast cancer in Arab nations. Int. J. Surg. 2010, 8, 448–452. [Google Scholar]
- Uhrhammer, N.; Abdelouahab, A.; Lafarge, L.; Feillel, V.; Ben Dib, A.; Bignon, Y.J. BRCA1 mutations in Algerian breast cancer patients: High frequency in young, sporadic cases. Int. J. Med. Sci. 2008, 5, 197–202. [Google Scholar]
- Corbex, M.; Bouzbid, S.; Boffetta, P. Features of breast cancer in developing countries, examples from North-Africa. Eur. J. Cancer 2014, 50, 1808–1818. [Google Scholar]
- Smaili, F.; Boudjella, A.; Dib, A.; Braikia, S.; Zidane, H.; Reggad, R.; Bendib, A.; Abdelouahab, A.; Bereksi-Reguig, F.; Yekrou, D.; et al. Epidemiology of breast cancer in women based on diagnosis data from oncologists and senologists in Algeria. Cancer Treat. Res. Commun. 2020, 25, 100220. [Google Scholar] [PubMed]
- Bernardo, G.; Le Noci, V.; Di Modica, M.; Montanari, E.; Triulzi, T.; Pupa, S.M.; Tagliabue, E.; Sommariva, M.; Sfondrini, L. The Emerging Role of the Microbiota in Breast Cancer Progression. Cells 2023, 12, 1945. [Google Scholar] [CrossRef]
- German, R.; Marino, N.; Hemmerich, C.; Podicheti, R.; Rusch, D.B.; Stiemsma, L.T.; Gao, H.; Xuei, X.; Rockey, P.; Storniolo, A.M. Exploring breast tissue microbial composition and the association with breast cancer risk factors. Breast Cancer Res. BCR 2023, 25, 82. [Google Scholar] [PubMed]
- Esposito, M.V.; Fosso, B.; Nunziato, M.; Casaburi, G.; D’Argenio, V.; Calabrese, A.; D’Aiuto, M.; Botti, G.; Pesole, G.; Salvatore, F. Microbiome composition indicate dysbiosis and lower richness in tumor breast tissues compared to healthy adjacent paired tissue, within the same women. BMC Cancer 2022, 22, 30. [Google Scholar]
- Thompson, K.J.; Ingle, J.N.; Tang, X.; Chia, N.; Jeraldo, P.R.; Walther-Antonio, M.R.; Kandimalla, K.K.; Johnson, S.; Yao, J.Z.; Harrington, S.C.; et al. A comprehensive analysis of breast cancer microbiota and host gene expression. PLoS ONE 2017, 12, e0188873. [Google Scholar]
- Niccolai, E.; Baldi, S.; Nannini, G.; Gensini, F.; Papi, L.; Vezzosi, V.; Bianchi, S.; Orzalesi, L.; Ramazzotti, M.; Amedei, A. Breast cancer: The first comparative evaluation of oncobiome composition between males and females. Biol. Sex Differ. 2023, 14, 37. [Google Scholar]
- Xuan, C.; Shamonki, J.M.; Chung, A.; Dinome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar]
- Allali, I.; Delgado, S.; Marron, P.I.; Astudillo, A.; Yeh, J.J.; Ghazal, H.; Amzazi, S.; Keku, T.; Azcarate-Peril, M.A. Gut microbiome compositional and functional differences between tumor and non-tumor adjacent tissues from cohorts from the US and Spain. Gut Microbes 2015, 6, 161–172. [Google Scholar] [PubMed]
- Cao, Q.; Sun, X.; Rajesh, K.; Chalasani, N.; Gelow, K.; Katz, B.; Shah, V.H.; Sanyal, A.J.; Smirnova, E. Effects of Rare Microbiome Taxa Filtering on Statistical Analysis. Front. Microbiol. 2020, 11, 607325. [Google Scholar]
- Walker, S.P.; Tangney, M.; Claesson, M.J. Sequence-Based Characterization of Intratumoral Bacteria-A Guide to Best Practice. Front. Oncol. 2020, 10, 179. [Google Scholar]
- Karstens, L.; Asquith, M.; Davin, S.; Fair, D.; Gregory, W.T.; Wolfe, A.J.; Braun, J.; McWeeney, S.; Gilbert, J.A. Controlling for Contaminants in Low-Biomass 16S rRNA Gene Sequencing Experiments. mSystems 2019, 4, e00290-19. [Google Scholar] [PubMed]
- Nearing, J.T.; Douglas, G.M.; Hayes, M.G.; MacDonald, J.; Desai, D.K.; Allward, N.; Jones, C.M.A.; Wright, R.J.; Dhanani, A.S.; Comeau, A.M.; et al. Microbiome differential abundance methods produce different results across 38 datasets. Nat. Commun. 2022, 13, 342. [Google Scholar]
- Fernández, M.F.; Reina-Pérez, I.; Astorga, J.M.; Rodríguez-Carrillo, A.; Plaza-Díaz, J.; Fontana, L. Breast Cancer and Its Relationship with the Microbiota. Int. J. Environ. Res. Public Health 2018, 15, 342. [Google Scholar]
- Bodai, B.I.; Nakata, T.E. Breast Cancer: Lifestyle, the Human Gut Microbiota/Microbiome, and Survivorship. Perm. J. 2020, 24, 19.129. [Google Scholar]
- Wu, A.H.; Vigen, C.; Tseng, C.; Garcia, A.A.; Spicer, D. Effect of Chemotherapy on the Gut Microbiome of Breast Cancer Patients During the First Year of Treatment. Breast Cancer 2022, 14, 433–451. [Google Scholar]
- Parida, S.; Sharma, D. The power of small changes: Comprehensive analyses of microbial dysbiosis in breast cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 392–405. [Google Scholar]
- Tamburini, F.B.; Maghini, D.; Oduaran, O.H.; Brewster, R.; Hulley, M.R.; Sahibdeen, V.; Norris, S.A.; Tollman, S.; Kahn, K.; Wagner, R.G.; et al. Short- and long-read metagenomics of urban and rural South African gut microbiomes reveal a transitional composition and undescribed taxa. Nat. Commun. 2022, 13, 926. [Google Scholar] [PubMed]
- De Filippo, C.; Di Paola, M.; Ramazzotti, M.; Albanese, D.; Pieraccini, G.; Banci, E.; Miglietta, F.; Cavalieri, D.; Lionetti, P. Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front. Microbiol. 2017, 8, 1979. [Google Scholar]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 2016, 21, 400–405. [Google Scholar] [PubMed]
- Mukherjee, P.K.; Wang, H.; Retuerto, M.; Zhang, H.; Burkey, B.; Ghannoum, M.A.; Eng, C. Bacteriome and mycobiome associations in oral tongue cancer. Oncotarget 2017, 8, 97273–97289. [Google Scholar]
- Joshi, V.; Matthews, C.; Aspiras, M.; de Jager, M.; Ward, M.; Kumar, P. Smoking decreases structural and functional resilience in the subgingival ecosystem. J. Clin. Periodontol. 2014, 41, 1037–1047. [Google Scholar]
- Yan, X.; Yang, M.; Liu, J.; Gao, R.; Hu, J.; Li, J.; Zhang, L.; Shi, Y.; Guo, H.; Cheng, J.; et al. Discovery and validation of potential bacterial biomarkers for lung cancer. Am. J. Cancer Res. 2015, 5, 3111–3122. [Google Scholar]
- Rodriguez, R.M.; Hernandez, B.Y.; Menor, M.; Deng, Y.; Khadka, V.S. The landscape of bacterial presence in tumor and adjacent normal tissue across 9 major cancer types using TCGA exome sequencing. Comput. Struct. Biotechnol. J. 2020, 18, 631–641. [Google Scholar]
- Hampelska, K.; Jaworska, M.M.; Babalska, Z.; Karpiński, T.M. The Role of Oral Microbiota in Intra-Oral Halitosis. J. Clin. Med. 2020, 9, 2484. [Google Scholar]
- Khan, A.A.; Sirsat, A.T.; Singh, H.; Cash, P. Microbiota and cancer: Current understanding and mechanistic implications. Clin. Transl. Oncol. 2022, 24, 193–202. [Google Scholar]
- Mikó, E.; Kovács, T.; Sebő, É.; Tóth, J.; Csonka, T.; Ujlaki, G.; Sipos, A.; Szabó, J.; Méhes, G.; Bai, P. Microbiome-Microbial Metabolome-Cancer Cell Interactions in Breast Cancer-Familiar, but Unexplored. Cells 2019, 8, 293. [Google Scholar]

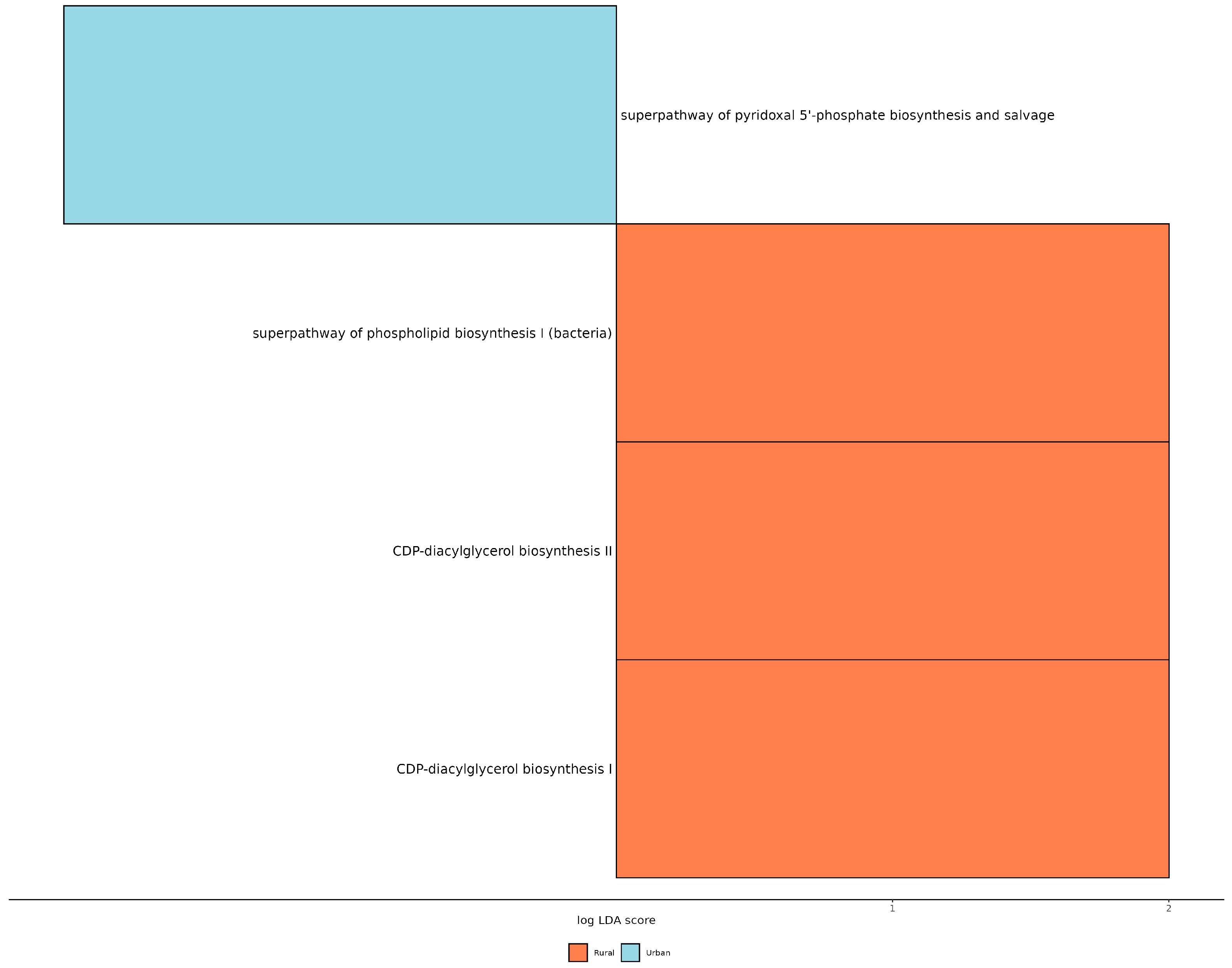

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age | Weight (kg) | Area | Family Background | Cancer Type | Cancer Grade | Radio/Chemotherapy |
|---|---|---|---|---|---|---|---|
| 1 | ≥50 | 50 | U | NO | IDC | Nottingham type II | Chemotherapy |
| 50> | 64 | R | YES | IDC | Nottingham type III | Chemotherapy | |
| 3 | ≥50 | 56 | R | NO | IDC | Nottingham II | Chemotherapy |
| 4 | 50> | 72 | R | NO | IDC | Nottingham type II | Chemotherapy |
| 5 | 50> | 76 | R | NO | IDC | Nottingham II | Chemotherapy |
| 6 | 50> | 72 | R | NO | IDC | Nottingham type II | Chemotherapy |
| 7 | ≥50 | 76 | U | NO | IDC | Nottingham type III | Chemotherapy |
| 8 | ≥50 | 55 | U | NO | IDC | Nottingham type I | Chemotherapy |
| 9 | ≥50 | 70 | R | NO | ILC | Nottingham type II | Radiotherapy |
| 10 | 50> | 60 | R | YES | IDC | Nottingham type II | Chemotherapy |
| 11 | 50> | 80 | U | NO | IDC | Nottingham type II | Chemotherapy |
| 12 | ≥50 | 70 | U | YES | IDC | Nottingham type II | Chemotherapy |
| 13 | 50> | 72 | U | YES | IDC | Nottingham type III | Chemotherapy |
| 14 | 50> | 74 | U | YES | IDC | Nottingham type II | Chemotherapy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ait Zenati, F.; Baldi, S.; Di Gloria, L.; Djoudi, F.; Bertorello, S.; Ramazzotti, M.; Niccolai, E.; Amedei, A. Compositional and Functional Disparities in the Breast Oncobiome Between Patients Living in Urban or Rural Areas. Genes 2025, 16, 806. https://doi.org/10.3390/genes16070806
Ait Zenati F, Baldi S, Di Gloria L, Djoudi F, Bertorello S, Ramazzotti M, Niccolai E, Amedei A. Compositional and Functional Disparities in the Breast Oncobiome Between Patients Living in Urban or Rural Areas. Genes. 2025; 16(7):806. https://doi.org/10.3390/genes16070806
Chicago/Turabian StyleAit Zenati, Fazia, Simone Baldi, Leandro Di Gloria, Ferhat Djoudi, Sara Bertorello, Matteo Ramazzotti, Elena Niccolai, and Amedeo Amedei. 2025. "Compositional and Functional Disparities in the Breast Oncobiome Between Patients Living in Urban or Rural Areas" Genes 16, no. 7: 806. https://doi.org/10.3390/genes16070806
APA StyleAit Zenati, F., Baldi, S., Di Gloria, L., Djoudi, F., Bertorello, S., Ramazzotti, M., Niccolai, E., & Amedei, A. (2025). Compositional and Functional Disparities in the Breast Oncobiome Between Patients Living in Urban or Rural Areas. Genes, 16(7), 806. https://doi.org/10.3390/genes16070806