
Genome-Wide Analysis of HIPP Gene Family in Maize Reveals Its Role in the Cadmium Stress Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Treatments
2.2. RNA Isolation and Real-Time RT-PCR Analysis
2.3. Identification and Physicochemical Properties of HIPP Genes in Maize
2.4. Phylogenetic Analysis and Classification
2.5. Conserved Motif, Domain, and Gene Structure Analysis of ZmHIPPs
2.6. Chromosome Distribution and Synteny Analysis in ZmHIPP Gene Family
2.7. Analysis of ZmHIPP Cis-Acting Elements
2.8. Expression Profiling of ZmHIPPs
2.9. Determination of Cd Concentration
3. Results
3.1. Characterization and Physicochemical Properties of HIPP Genes in Maize
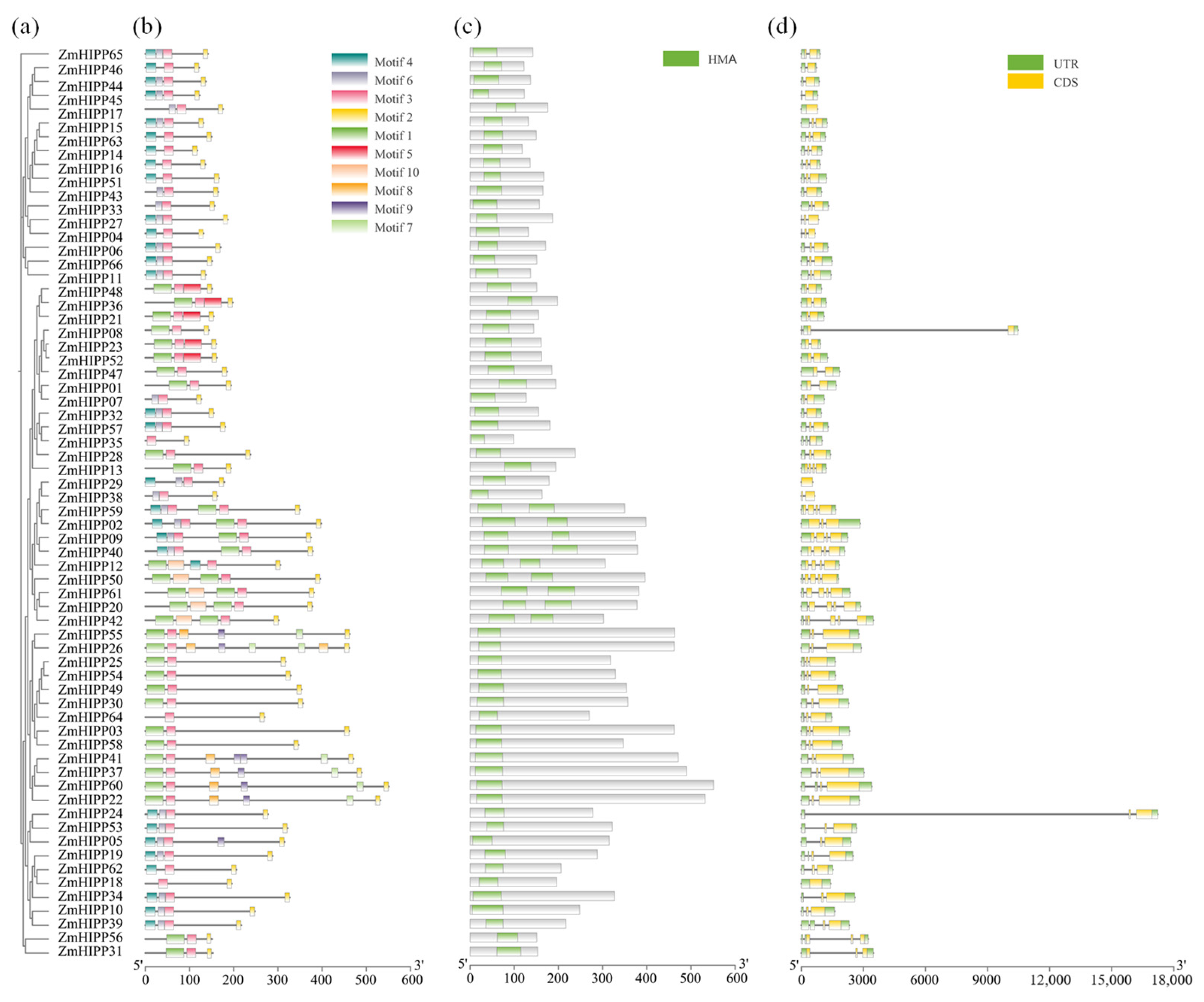
3.2. Phylogenetic Analysis of ZmHIPP Proteins Based on Conserved Motifs and Gene Structure
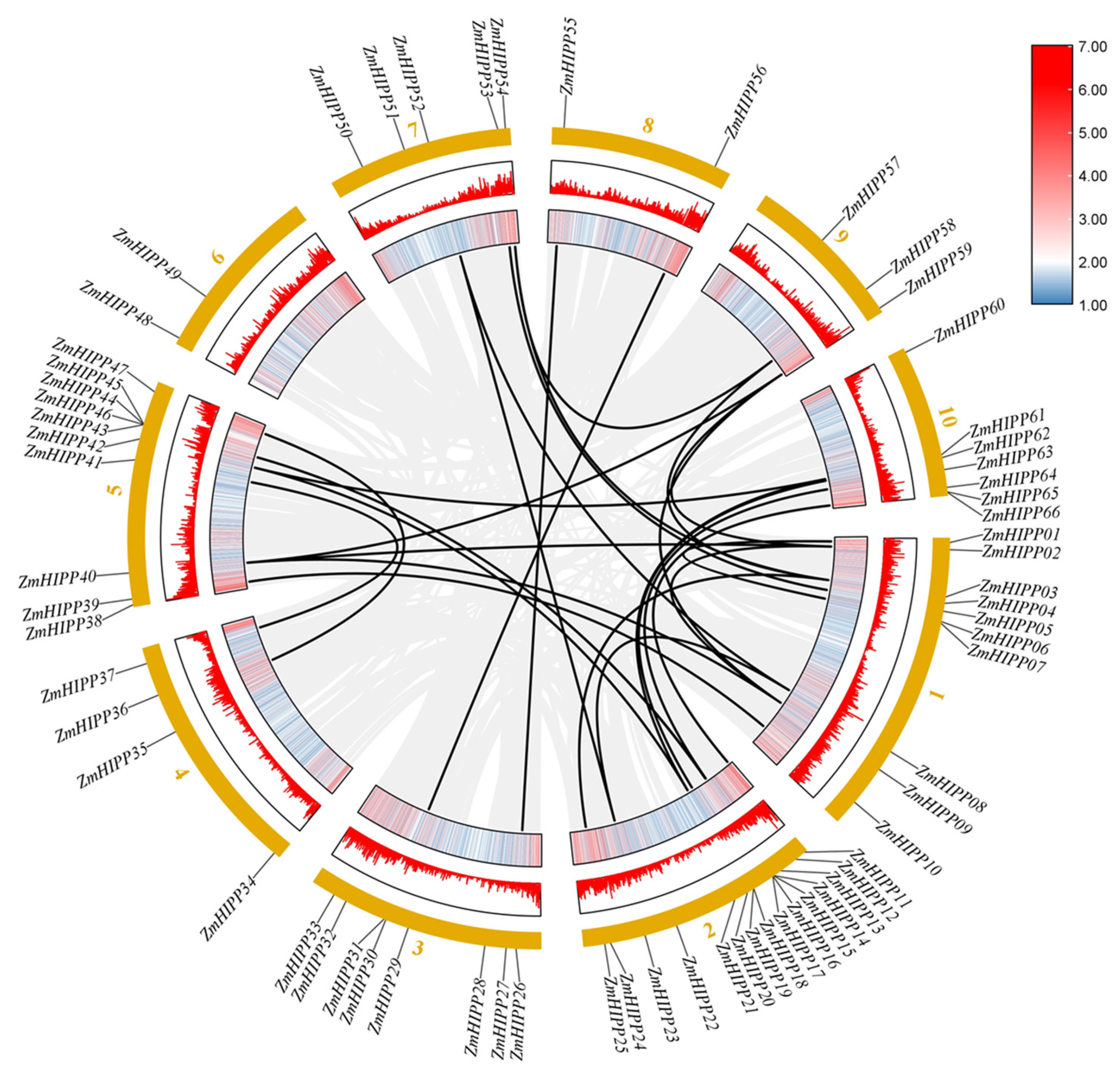
3.3. Synteny Analysis in HIPP Genes
3.4. Analyses of ZmHIPP Gene Promoter Cis-Regulatory Elements
3.5. Expression Profiling of ZmHIPP Genes in Maize
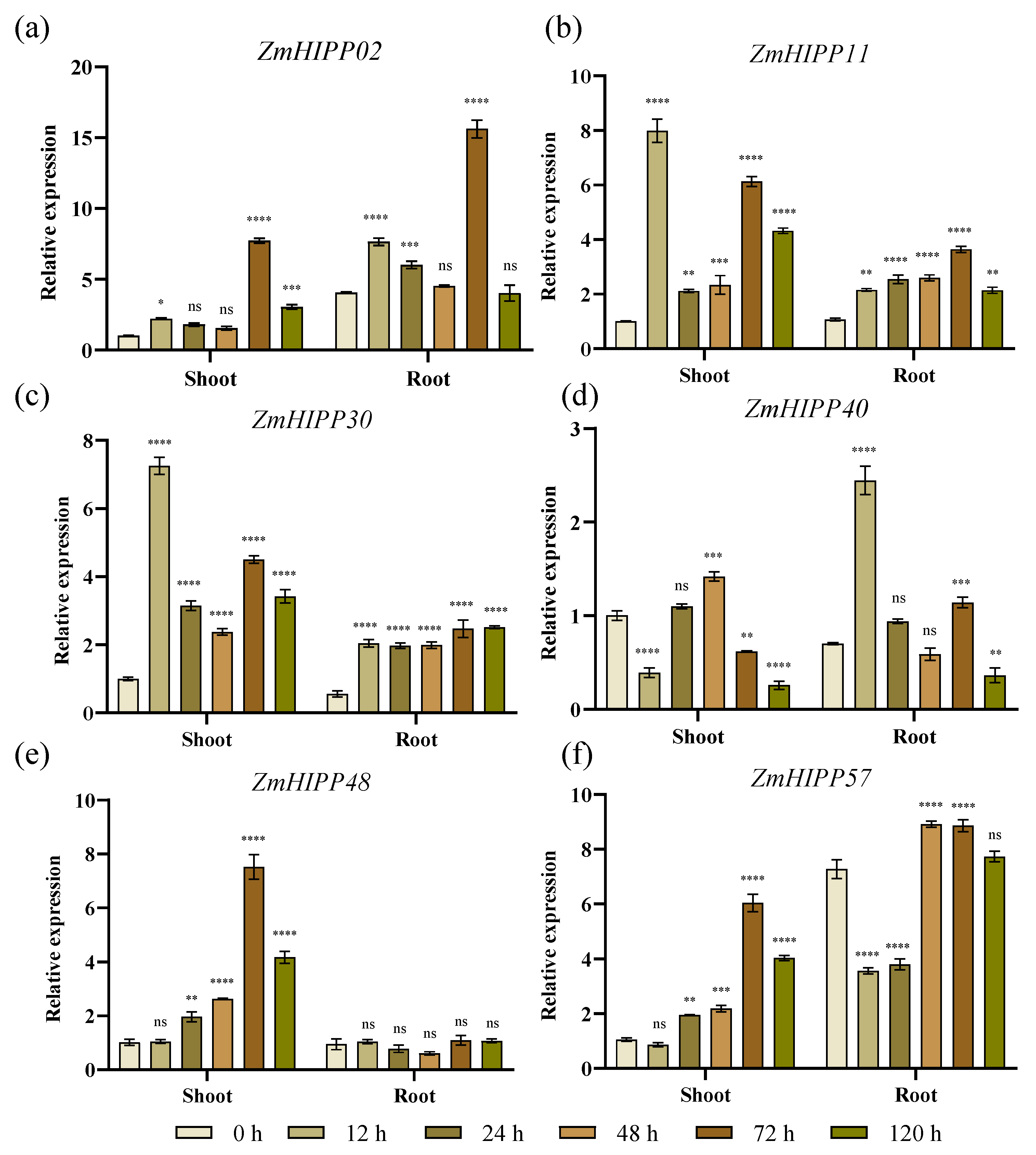
3.6. ZmHIPP Expression Under Cd Stress and Corresponding Cd Accumulation in Maize
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| As | Arsenic |
| Cd | Cadmium |
| Cu | Copper |
| FPKM | Fragments Per Kilobase of transcript per Million mapped reads |
| GFF | General Feature Format |
| GTF | Gene Transfer Format |
| HIPP | Heavy metal-associated isoprenylated plant protein |
| HMA | Heavy metal-associated |
| HMM | Hidden Markov model |
| Pb | Lead |
| STRE | Stress response promoter element |
References
- Liu, C.C.; Wen, L.; Cui, Y.J.; Ahammed, G.J.; Cheng, Y. Metal transport proteins and transcription factor networks in plant responses to cadmium stress. Plant Cell Rep. 2024, 43, 218. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.P.; Cao, L.; Zhang, C.; Pan, W.C.; Wang, W.; Yu, X.; Li, Y.P.; Fan, T.T.; Miao, M.; Tang, X.F.; et al. MYB43 as a novel substrate for CRL4PRL1 E3 ligases negatively regulates cadmium tolerance through transcriptional inhibition of HMAs in Arabidopsis. New Phytol. 2022, 234, 884–901. [Google Scholar] [CrossRef] [PubMed]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Tack, F.M.G.; Sebastian, A.; Prasad, M.N.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Khan, I.U.; Rono, J.K.; Zhang, B.Q.; Liu, X.S.; Wang, M.Q.; Wang, L.L.; Wu, X.C.; Chen, X.; Cao, H.W.; Yang, Z.M. Identification of novel rice (Oryza sativa) HPP and HIPP genes tolerant to heavy metal toxicity. Ecotoxicol. Environ. Saf. 2019, 175, 8–18. [Google Scholar] [CrossRef]
- de Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; de Oliveira, L.F.V.; Zanettini, M.H.B.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef]
- Parasyri, A.; Barth, O.; Zschiesche, W.; Humbeck, K. The Barley Heavy Metal Associated Isoprenylated Plant Protein HvFP1 Is Involved in a Crosstalk between the Leaf Development and Abscisic Acid-Related Drought Stress Responses. Plants 2022, 11, 21. [Google Scholar] [CrossRef]
- Wang, Z.K.; Zhang, H.; Li, Y.B.; Chen, Y.M.; Tang, X.; Zhao, J.; Yu, F.F.; Wang, H.Y.; Xiao, J.; Liu, J.; et al. Isoprenylation modification is required for HIPP1-mediated powdery mildew resistance in wheat. Plant Cell Environ. 2023, 46, 288–305. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.; Xu, J. The R2R3-MYB Transcription Factor MYB49 Regulates Cadmium Accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef]
- Tehseen, M.; Cairns, N.; Sherson, S.; Cobbett, C.S. Metallochaperone-like genes in Arabidopsis thaliana. Metallomics 2010, 2, 556–564. [Google Scholar] [CrossRef]
- Suzuki, N.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Functional characterization of a heavy metal binding protein CdI19 from Arabidopsis. Plant. J. 2002, 32, 165–173. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, X.; Feng, S.; Zhao, Y.; Wang, L.; Rono, J.; Li, H.; Yang, Z. Developing a cadmium resistant rice genotype with OsHIPP29 locus for limiting cadmium accumulation in the paddy crop. Chemosphere 2020, 247, 125958. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.N.; Wang, M.Q.; Li, C.; Cao, H.W.; Rono, J.K.; Yang, Z.M. The metallochaperone OsHIPP56 gene is required for cadmium detoxification in rice crops. Environ. Exp. Bot. 2022, 193, 10. [Google Scholar] [CrossRef]
- Chen, K.; Chen, P.; Qiu, X.; Chen, J.; Gao, G.; Wang, X.; Zhu, A.; Yu, C. Regulating role of abscisic acid on cadmium enrichment in ramie (Boehmeria nivea L.). Sci. Rep. 2021, 11, 22045. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Rono, J.K.; Liu, X.; Feng, S.; Li, H.; Chen, X.; Yang, Z. Functional characterization of a new metallochaperone for reducing cadmium concentration in rice crop. J. Clean. Prod. 2020, 272, 11. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, M.; Zhang, J.; Huang, L.; Chen, X.; Jiang, M.; Tan, M. Profiling of rice Cd-tolerant genes through yeast-based cDNA library survival screening. Plant Physiol. Biochem. 2020, 155, 429–436. [Google Scholar] [CrossRef]
- Cheng, D.; Tan, M.; Yu, H.; Li, L.; Zhu, D.; Chen, Y.; Jiang, M. Comparative analysis of Cd-responsive maize and rice transcriptomes highlights Cd co-modulated orthologs. BMC Genom. 2018, 19, 709. [Google Scholar] [CrossRef]
- Radakovic, Z.S.; Anjam, M.S.; Escobar, E.; Chopra, D.; Cabrera, J.; Silva, A.C.; Escobar, C.; Sobczak, M.; Grundler, F.M.W.; Siddique, S. Arabidopsis HIPP27 is a host susceptibility gene for the beet cyst nematode Heterodera schachtii. Mol. Plant Pathol. 2018, 19, 1917–1928. [Google Scholar] [CrossRef]
- Zhang, H.; Zhai, G.W.; Ni, X.L.; Liu, Z.W.; Song, T.; Han, Y.; Wang, Y.; Shao, Y.; Wang, F.L.; Zou, G.H.; et al. Genome-wide identification of HIPP genes family in sorghum reveals the novel role of SbHIPP40 in accumulation of cadmium. J. Hazard. Mater. 2025, 494, 138478. [Google Scholar] [CrossRef]
- Xia, H.Y.; Jing, X.; He, H.Q.; Peng, J.W.; Liu, Y.Y.; Sun, W.Y.; Wang, X.Z.; Yuan, Z.; Wu, J.X.; Zhang, M.Y.; et al. Genome-wide identification of the HIPPs gene family and functional validation of MsHIPP12 in enhancing cadmium tolerance in Medicago sativa. J. Hazard. Mater. 2025, 491, 137894. [Google Scholar] [CrossRef]
- Wei, Y.F.; Peng, X.Q.; Wang, X.J.; Wang, C. The heavy metal-associated isoprenylated plant protein (HIPP) gene family plays a crucial role in cadmium resistance and accumulation in the tea plant (Camellia sinensis L.). Ecotoxicol. Environ. Saf. 2023, 260, 115077. [Google Scholar] [CrossRef]
- Huang, G.Y.; Hu, Y.N.; Li, F.X.; Zuo, X.R.; Wang, X.Y.; Li, F.Y.; Li, R.M. Genome-wide characterization of heavy metal-associated isoprenylated plant protein gene family from Citrus sinensis in response to huanglongbing. Front. Plant Sci. 2024, 15, 1369883. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Sartor, R.C.; Shen, Z.; Schmitz, R.J.; Wu, K.J.; Urich, M.A.; Nery, J.R.; Smith, L.G.; Schnable, J.C.; Ecker, J.R.; et al. Integration of omic networks in a developmental atlas of maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhu, J.L.; Yin, Y.Z.; Zhang, X.Y.; Dai, Y.X.; Xing, Y.P.; Cheng, X.P.; Zhang, A.; Li, C.; Zhu, Y.S.; et al. Dynamic transcriptome profiling revealed a key gene ZmJMJ20 and pathways associated with cadmium stress in maize. Ecotoxicol. Environ. Saf. 2024, 277, 116352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Liu, J.; Niu, Y.; Chen, Y.; Hao, Y.; Zhao, J.; Sun, L.; Wang, H.; Xiao, J.; et al. Characterization of the Heavy-Metal-Associated Isoprenylated Plant Protein (HIPP) Gene Family from Triticeae Species. Int. J. Mol. Sci. 2020, 21, 6191. [Google Scholar] [CrossRef]
- Barr, Z.K.; Werner, T.; Tilsner, J. Heavy Metal-Associated Isoprenylated Plant Proteins (HIPPs) at Plasmodesmata: Exploring the Link between Localization and Function. Plants 2023, 12, 3015. [Google Scholar] [CrossRef]
- Cao, H.W.; Li, C.; Zhang, B.Q.; Rono, J.K.; Yang, Z.M. A Metallochaperone HIPP33 Is Required for Rice Zinc and Iron Homeostasis and Productivity. Agronomy 2022, 12, 488. [Google Scholar] [CrossRef]
- Ma, L.; An, R.; Jiang, L.; Zhang, C.; Li, Z.; Zou, C.; Yang, C.; Pan, G.; Lubberstedt, T.; Shen, Y. Effects of ZmHIPP on lead tolerance in maize seedlings: Novel ideas for soil bioremediation. J. Hazard. Mater. 2022, 430, 128457. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, Z.; Yan, T.; Lu, R.; Peng, C.; Li, S.; Jing, Y. Arbuscular mycorrhizal fungi alleviate Cd phytotoxicity by altering Cd subcellular distribution and chemical forms in Zea mays. Ecotoxicol. Environ. Saf. 2019, 171, 352–360. [Google Scholar] [CrossRef]
- Chang, J.; Xie, Y.; Zhang, H.; Zhang, S.; Zhao, F. The vacuolar transporter OsNRAMP2 mediates Fe remobilization during germination and affects Cd distribution to rice grain. Plant Soil. 2022, 476, 79–95. [Google Scholar] [CrossRef]
- Cao, H.W.; Zhao, Y.N.; Liu, X.S.; Rono, J.K.; Yang, Z.M. A metal chaperone OsHIPP16 detoxifies cadmium by repressing its accumulation in rice crops. Environ. Pollut. 2022, 311, 120058. [Google Scholar] [CrossRef]
- Zhang, X.; Feng, H.; Feng, C.; Xu, H.; Huang, X.; Wang, Q.; Duan, X.; Wang, X.; Wei, G.; Huang, L.; et al. Isolation and characterisation of cDNA encoding a wheat heavy metal-associated isoprenylated protein involved in stress responses. Plant Biol. 2015, 17, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Zschiesche, W.; Barth, O.; Daniel, K.; Böhme, S.; Rausche, J.; Humbeck, K. The zinc-binding nuclear protein HIPP3 acts as an upstream regulator of the salicylate-dependent plant immunity pathway and of flowering time in Arabidopsis thaliana. New Phytol. 2015, 207, 1084–1096. [Google Scholar] [CrossRef]
- Sheng, Y.B.; Yan, X.X.; Huang, Y.; Han, Y.Y.; Zhang, C.; Ren, Y.B.; Fan, T.T.; Xiao, F.M.; Liu, Y.S.; Cao, S.Q. The WRKY transcription factor, WRKY13, activates PDR8 expression to positively regulate cadmium tolerance in Arabidopsis. Plant Cell Environ. 2019, 42, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Hou, Y.Y.; Sun, Y.Y.; Chen, X.X.; Wang, G.Y.; Wang, H.C.; Zhu, B.; Du, X.Y. The maize WRKY transcription factor ZmWRKY64 confers cadmium tolerance in Arabidopsis and maize (Zea mays L.). Plant Cell Rep. 2024, 43, 44. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.X.; Huang, Y.; Song, H.; Chen, F.; Geng, Q.L.; Hu, M.; Zhang, C.; Wu, X.; Fan, T.T.; Cao, S.Q. A MYB4-MAN3-Mannose-MNB1 signaling cascade regulates cadmium tolerance in arabidopsis. PLoS Genet. 2021, 17, e1009636. [Google Scholar] [CrossRef]
- Wei, X.; Geng, M.H.; Yuan, J.C.; Zhan, J.J.; Liu, L.S.; Chen, Y.L.; Wang, Y.; Qin, W.Q.; Duan, H.Y.; Zhao, H.; et al. GhRCD1 promotes cotton tolerance to cadmium by regulating the GhbHLH12-GhMYB44-GhHMA1 transcriptional cascade. Plant Biotechnol. J. 2024, 22, 1777–1796. [Google Scholar] [CrossRef]
- Zhang, M.; Gao, J.Y.; Dong, S.C.; Chang, M.H.; Zhu, J.X.; Guo, D.L.; Guo, C.H.; Bi, Y.D. Alfalfa MsbHLH115 confers tolerance to cadmium stress through activating the iron deficiency response in Arabidopsis thaliana. Front. Plant Sci. 2024, 15, 1358673. [Google Scholar] [CrossRef]
- Anwar, A.; Wang, Y.D.; Chen, M.Q.; Zhang, S.W.; Wang, J.M.; Feng, Y.Q.; Xue, Y.X.; Zhao, M.F.; Su, W.; Chen, R.Y.; et al. Zero-valent iron (nZVI) nanoparticles mediate SlERF1 expression to enhance cadmium stress tolerance in tomato. J. Hazard. Mater. 2024, 468, 133829. [Google Scholar] [CrossRef]
- Chang, J.; Huang, S.; Konishi, N.; Wang, P.; Chen, J.; Huang, X.; Ma, J.; Zhao, F. Overexpression of the manganese/cadmium transporter OsNRAMP5 reduces cadmium accumulation in rice grain. J. Exp. Bot. 2020, 71, 5705–5715. [Google Scholar] [CrossRef]
- Zhang, W.; Guan, M.; Chen, M.; Lin, X.; Xu, P.; Cao, Z. Mutation of OsNRAMP5 reduces cadmium xylem and phloem transport in rice plants and its physiological mechanism. Environ. Pollut. 2024, 341, 11. [Google Scholar] [CrossRef]
- Chen, Y.; Chao, Z.; Jin, M.; Wang, Y.; Li, Y.; Wu, J.; Xiao, Y.; Peng, Y.; Lv, Q.; Gui, S.; et al. A heavy metal transporter gene ZmHMA3a promises safe agricultural production on cadmium-polluted arable land. J. Genet. Genomics 2023, 50, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Chai, J.; Zhu, P.; Shanklin, J.; Liu, Q. Structural mechanism of intracellular autoregulation of zinc uptake in ZIP transporters. Nat. Commun. 2023, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Wang, M.; Jiang, Y.; Wang, C.; Ow, D. Overexpression of OsABCG48 Lowers Cadmium in Rice (Oryza sativa L.). Agronomy 2021, 11, 12. [Google Scholar] [CrossRef]
- Guo, T.; Weber, H.; Niemann, M.C.E.; Theisl, L.; Leonte, G.; Novak, O.; Werner, T. Arabidopsis HIPP proteins regulate endoplasmic reticulum-associated degradation of CKX proteins and cytokinin responses. Mol. Plant 2021, 14, 1918–1934. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, C.; Zhang, Z.; Zhu, Y.; Tian, J.; Yu, K.; Hou, J.; Luo, D.; Cai, J.; Zhu, Y. Genome-Wide Analysis of HIPP Gene Family in Maize Reveals Its Role in the Cadmium Stress Response. Genes 2025, 16, 770. https://doi.org/10.3390/genes16070770
Gao C, Zhang Z, Zhu Y, Tian J, Yu K, Hou J, Luo D, Cai J, Zhu Y. Genome-Wide Analysis of HIPP Gene Family in Maize Reveals Its Role in the Cadmium Stress Response. Genes. 2025; 16(7):770. https://doi.org/10.3390/genes16070770
Chicago/Turabian StyleGao, Chunyan, Zhirui Zhang, Yuxuan Zhu, Jiaxin Tian, Kaili Yu, Jinbo Hou, Dan Luo, Jian Cai, and Youcheng Zhu. 2025. "Genome-Wide Analysis of HIPP Gene Family in Maize Reveals Its Role in the Cadmium Stress Response" Genes 16, no. 7: 770. https://doi.org/10.3390/genes16070770
APA StyleGao, C., Zhang, Z., Zhu, Y., Tian, J., Yu, K., Hou, J., Luo, D., Cai, J., & Zhu, Y. (2025). Genome-Wide Analysis of HIPP Gene Family in Maize Reveals Its Role in the Cadmium Stress Response. Genes, 16(7), 770. https://doi.org/10.3390/genes16070770