
MYB Confers Sorafenib Resistance in Human Leukemia Cells via Inhibiting Ferroptosis Through FTH1 Upregulation

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Plasmid Construction
2.3. RNA Extraction and Quantitative Real-Time PCR (RT-qPCR)
2.4. Lentivirus Preparation and Infection
2.5. Western Blot
2.6. Determination of Reactive Oxygen Species (ROS)
2.7. Determination of Fe2+
2.8. CCK8 Assay
2.9. Transwell Assay
2.10. Statistical Analysis
3. Results
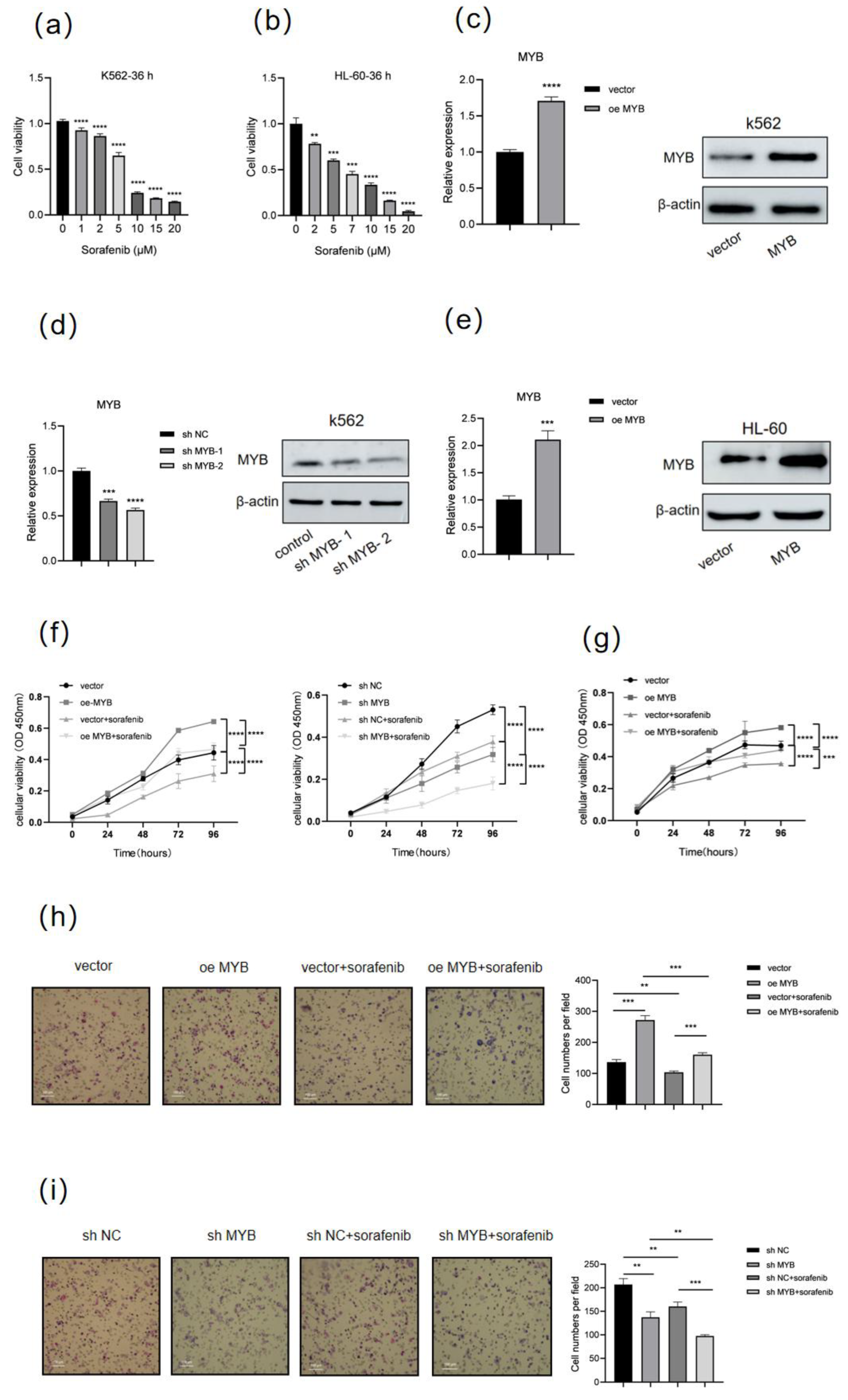
3.1. MYB Induces Drug Resistance in Human Leukemia Cells
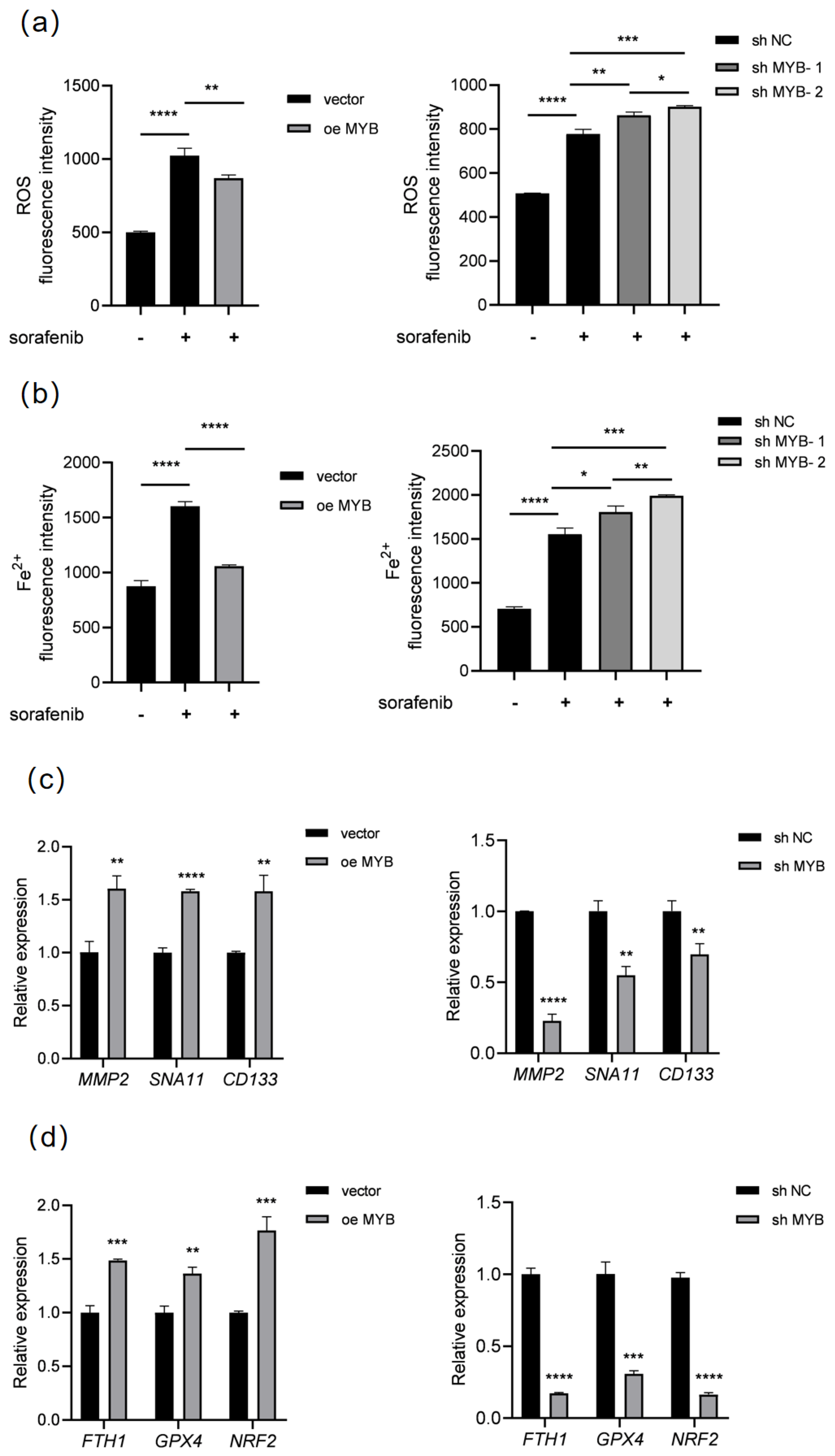
3.2. MYB Inhibits Ferroptosis in K562 Cells
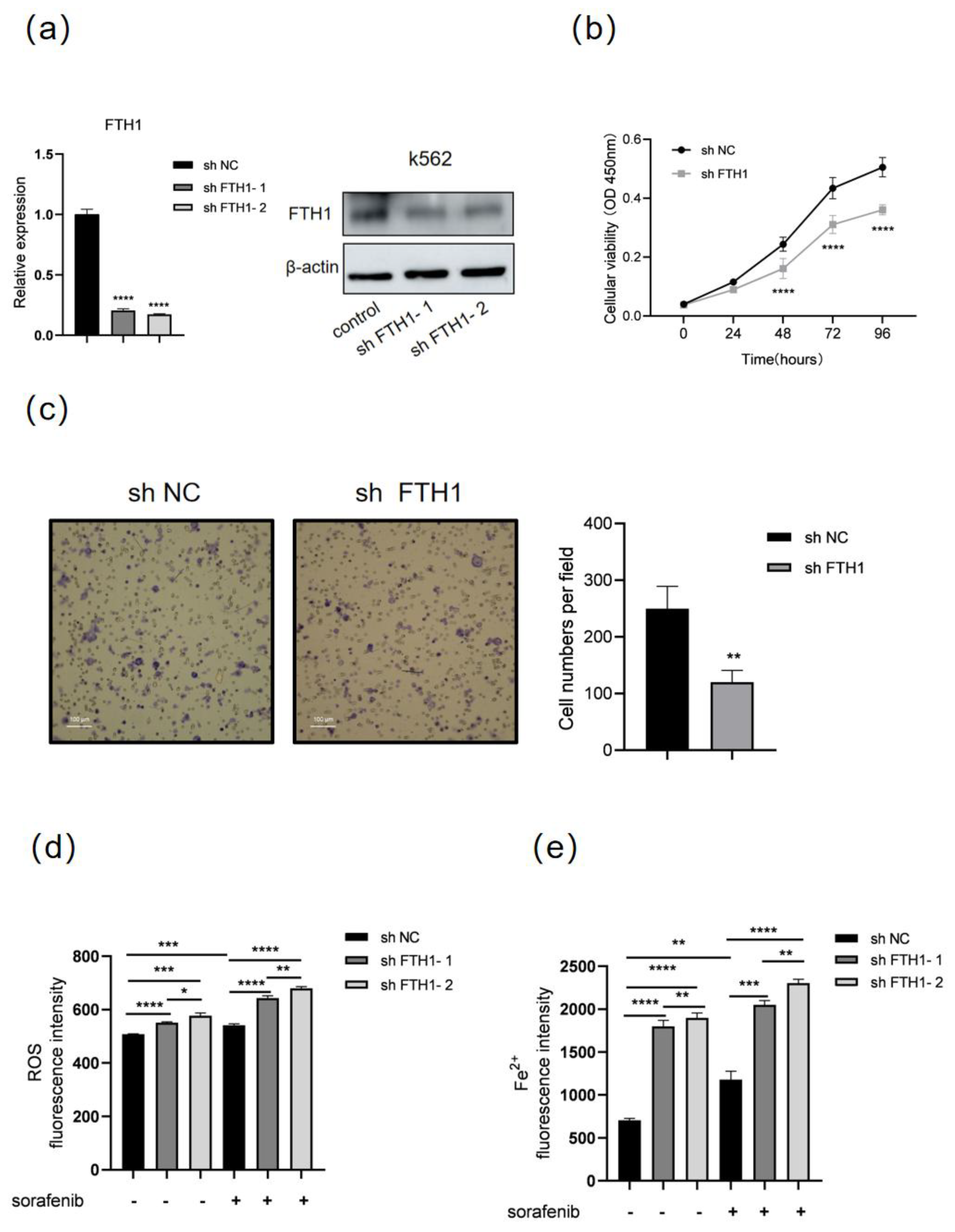
3.3. FTH1 Knockdown Induces Ferroptosis and Enhances Sorafenib Sensitivity in K562 Cells
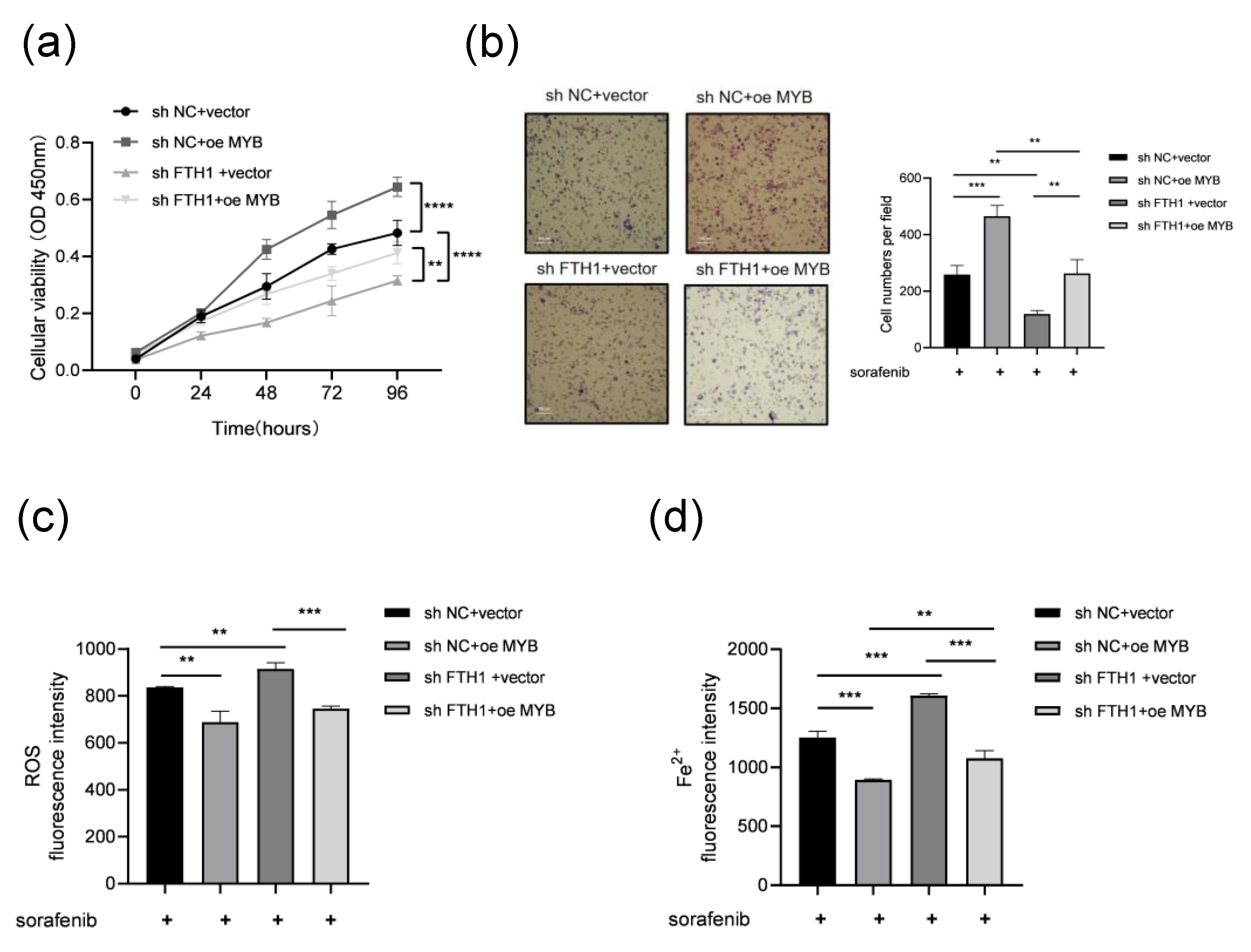
3.4. MYB Inhibits Ferroptosis and Induces Sorafenib Resistance Through FTH1 in Leukemia Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sheiness, D.; Gardinier, M. Expression of a proto-oncogene (proto-myb) in hemopoietic tissues of mice. Mol. Cell. Biol. 1984, 4, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Westin, E.H.; Gallo, R.C.; Arya, S.K.; Eva, A.; Souza, L.M.; Baluda, M.A.; Aaronson, S.A.; Wong-Staal, F. Differential expression of the amv gene in human hematopoietic cells. Proc. Natl. Acad. Sci. USA 1982, 79, 2194–2198. [Google Scholar] [CrossRef]
- Oh, I.H.; Reddy, E.P. The myb gene family in cell growth, differentiation and apoptosis. Oncogene 1999, 18, 3017–3033. [Google Scholar] [CrossRef]
- Pattabiraman, D.R.; Gonda, T.J. Role and potential for therapeutic targeting of MYB in leukemia. Leukemia 2013, 27, 269–277. [Google Scholar] [CrossRef]
- Zhou, Y.; Ness, S.A. Myb proteins: Angels and demons in normal and transformed cells. Front. Biosci. 2011, 16, 1109–1131. [Google Scholar] [CrossRef]
- Stenman, G.; Andersson, M.K.; Andrén, Y. New tricks from an old oncogene: Gene fusion and copy number alterations of MYB in human cancer. Cell Cycle 2010, 9, 2986–2995. [Google Scholar] [CrossRef]
- Biersack, B.; Höpfner, M. Emerging role of MYB transcription factors in cancer drug resistance. Cancer Drug Resist. 2024, 7, 15. [Google Scholar] [CrossRef]
- Iyer, R.; Fetterly, G.; Lugade, A.; Thanavala, Y. Sorafenib: A clinical and pharmacologic review. Expert Opin. Pharmacother. 2010, 11, 1943–1955. [Google Scholar] [CrossRef]
- Abdelgalil, A.A.; Alkahtani, H.M.; Al-Jenoobi, F.I. Sorafenib. Profiles Drug Subst. Excip. Relat. Methodol. 2019, 44, 239–266. [Google Scholar] [CrossRef]
- Metzelder, S.K.; Schroeder, T.; Lübbert, M.; Ditschkowski, M.; Götze, K.; Scholl, S.; Meyer, R.G.; Dreger, P.; Basara, N.; Fey, M.F.; et al. Long-term survival of sorafenib-treated FLT3-ITD-positive acute myeloid leukaemia patients relapsing after allogeneic stem cell transplantation. Eur. J. Cancer 2017, 86, 233–239. [Google Scholar] [CrossRef]
- Lange, A.; Jaskula, E.; Lange, J.; Dworacki, G.; Nowak, D.; Simiczyjew, A.; Mordak-Domagala, M.; Sedzimirska, M. The sorafenib anti-relapse effect after alloHSCT is associated with heightened alloreactivity and accumulation of CD8+PD-1+ (CD279+) lymphocytes in marrow. PLoS ONE 2018, 13, e0190525. [Google Scholar] [CrossRef]
- Muppidi, M.R.; Portwood, S.; Griffiths, E.A.; Thompson, J.E.; Ford, L.A.; Freyer, C.W.; Wetzler, M.; Wang, E.S. Decitabine and Sorafenib Therapy in FLT-3 ITD-Mutant Acute Myeloid Leukemia. Clin. Lymphoma Myeloma Leuk. 2015, 15, S73–S79. [Google Scholar] [CrossRef]
- Xia, S.; Pan, Y.; Liang, Y.; Xu, J.; Cai, X. The microenvironmental and metabolic aspects of sorafenib resistance in hepatocellular carcinoma. EBioMedicine 2020, 51, 102610. [Google Scholar] [CrossRef]
- Li, J.; Jia, Y.C.; Ding, Y.X.; Bai, J.; Cao, F.; Li, F. The crosstalk between ferroptosis and mitochondrial dynamic regulatory networks. Int. J. Biol. Sci. 2023, 19, 2756–2771. [Google Scholar] [CrossRef]
- Raven, E.P.; Lu, P.H.; Tishler, T.A.; Heydari, P.; Bartzokis, G. Increased iron levels and decreased tissue integrity in hippocampus of Alzheimer’s disease detected in vivo with magnetic resonance imaging. J. Alzheimers Dis. 2013, 37, 127–136. [Google Scholar] [CrossRef]
- Eling, N.; Reuter, L.; Hazin, J.; Hamacher-Brady, A.; Brady, N.R. Identification of artesunate as a specific activator of ferroptosis in pancreatic cancer cells. Oncoscience 2015, 2, 517–532. [Google Scholar] [CrossRef]
- Birsen, R.; Larrue, C.; Decroocq, J.; Johnson, N.; Guiraud, N.; Gotanegre, M.; Cantero-Aguilar, L.; Grignano, E.; Huynh, T.; Fontenay, M.; et al. APR-246 induces early cell death by ferroptosis in acute myeloid leukemia. Haematologica 2022, 107, 403–416. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, M.; Liu, M.; Yu, Y.; Zhang, Y.; Yang, J.; Xing, L.; Shao, Z.; Wang, H. Loss of FTH1 Induces Ferritinophagy-Mediated Ferroptosis in Anaemia of Myelodysplastic Syndromes. J. Cell Mol. Med. 2025, 29, e70350. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Wei, J.; Wu, X.; Luo, J.; Wei, H.; Ning, L.; He, Y. High expression level of the FTH1 gene is associated with poor prognosis in children with non-M3 acute myeloid leukemia. Front. Oncol. 2022, 12, 1068094. [Google Scholar] [CrossRef]
- Ali, A.; Shafarin, J.; Abu Jabal, R.; Aljabi, N.; Hamad, M.; Sualeh Muhammad, J.; Unnikannan, H.; Hamad, M. Ferritin heavy chain (FTH1) exerts significant antigrowth effects in breast cancer cells by inhibiting the expression of c-MYC. FEBS Open Bio 2021, 11, 3101–3114. [Google Scholar] [CrossRef]
- Mucenski, M.L.; McLain, K.; Kier, A.B.; Swerdlow, S.H.; Schreiner, C.M.; Miller, T.A.; Pietryga, D.W.; Scott, W.J., Jr.; Potter, S.S. A functional c-myb gene is required for normal murine fetal hepatic hematopoiesis. Cell 1991, 65, 677–689. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J. Long noncoding RNA ZFAS1 enhances adriamycin resistance in pediatric acute myeloid leukemia through the miR-195/Myb axis. RSC Adv. 2019, 9, 28126–28134. [Google Scholar] [CrossRef]
- Sarvaiya, P.J.; Schwartz, J.R.; Hernandez, C.P.; Rodriguez, P.C.; Vedeckis, W.V. Role of c-Myb in the survival of pre B-cell acute lymphoblastic leukemia and leukemogenesis. Am. J. Hematol. 2012, 87, 969–976. [Google Scholar] [CrossRef]
- Tian, M.; Tian, D.; Qiao, X.; Li, J.; Zhang, L. Modulation of Myb-induced NF-kB -STAT3 signaling and resulting cisplatin resistance in ovarian cancer by dietary factors. J. Cell. Physiol. 2019, 234, 21126–21134. [Google Scholar] [CrossRef]
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Broadening horizons: The role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 2021, 18, 280–296. [Google Scholar] [CrossRef] [PubMed]
- Mancias, J.D.; Wang, X.; Gygi, S.P.; Harper, J.W.; Kimmelman, A.C. Quantitative proteomics identifies NCOA4 as the cargo receptor mediating ferritinophagy. Nature 2014, 509, 105–109. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, C.; Sun, Q.; Li, Y.; Zhou, C.; Sun, C. Susceptibility of acute myeloid leukemia cells to ferroptosis and evasion strategies. Front. Mol. Biosci. 2023, 10, 1275774. [Google Scholar] [CrossRef] [PubMed]
- Wolosowicz, M.; Prokopiuk, S.; Kaminski, T.W. The Complex Role of Matrix Metalloproteinase-2 (MMP-2) in Health and Disease. Int. J. Mol. Sci. 2024, 25, 13691. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Li, T.; Bian, H.; Li, F.; Ju, Y.; Gao, S.; Su, J.; Ren, W.; Qin, C. SNAI1 promotes the development of HCC through the enhancement of proliferation and inhibition of apoptosis. FEBS Open Bio 2016, 6, 326–337. [Google Scholar] [CrossRef]
- Gisina, A.; Kim, Y.; Yarygin, K.; Lupatov, A. Can CD133 Be Regarded as a Prognostic Biomarker in Oncology: Pros and Cons. Int. J. Mol. Sci. 2023, 24, 17398. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wan, Y.; Jiang, Y.; Zhang, L.; Cheng, W. GPX4: The hub of lipid oxidation, ferroptosis, disease and treatment. Biochim. Biophys. Acta Rev. Cancer 2023, 1878, 188890. [Google Scholar] [CrossRef]
- Dodson, M.; Castro-Portuguez, R.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef]
- He, M.; Wang, Y.; Xie, J.; Pu, J.; Shen, Z.; Wang, A.; Li, T.; Wang, T.; Li, G.; Liu, Y.; et al. M(7)G modification of FTH1 and pri-miR-26a regulates ferroptosis and chemotherapy resistance in osteosarcoma. Oncogene 2024, 43, 341–353. [Google Scholar] [CrossRef]
- Jin, Y.; Qiu, J.; Lu, X.; Ma, Y.; Li, G. LncRNA CACNA1G-AS1 up-regulates FTH1 to inhibit ferroptosis and promote malignant phenotypes in ovarian cancer cells. Oncol. Res. 2023, 31, 169–179. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, S.S.; Gu, Y.R.; Xiao, L.X.; Ma, X.Y.; Chen, X.R.; Wang, J.L.; Liao, C.H.; Lin, B.L.; Huang, Y.H.; et al. CircPIAS1 promotes hepatocellular carcinoma progression by inhibiting ferroptosis via the miR-455-3p/NUPR1/FTH1 axis. Mol. Cancer 2024, 23, 113. [Google Scholar] [CrossRef] [PubMed]
- Booth, C.A.G.; Bouyssou, J.M.; Togami, K.; Armand, O.; Rivas, H.G.; Yan, K.; Rice, S.; Cheng, S.; Lachtara, E.M.; Bourquin, J.P.; et al. BPDCN MYB fusions regulate cell cycle genes, impair differentiation, and induce myeloid-dendritic cell leukemia. JCI Insight 2024, 9, e183889. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) | TRCN Number |
|---|---|---|---|
| sh MYB-1 | CCGGAACAGAATGGAACAGATGACCTCGAGGTCATCTGTTCCATTCTGTT TTTTTG | AATTCAAAAAAACAGAATGGAACAGATGACCTCGAGGTCATCTGTTCCATTCTGTT | TRCN0000009853 |
| sh MYB-2 | CCGGGCTCCTAATGTCAACCGAGAACTCGAGTTCTCGGTTGACATTAGGAGCTTTTTG | AATTCAAAAAGCTCCTAATGTCAACCGAGAACTCGAGTTCTCGGTTGACATTAGGAGC | TRCN0000288659 |
| sh FTH1-1 | CCGGGCCGAATCTTCCTTCAGGATACTCGAGTATCCTGAAGGAAGATTCGGCTTTTTG | AATTCAAAAAGCCGAATCTTCCTTCAGGATACTCGAGTATCCTGAAGGAAGATTCGGC | TRCN0000029433 |
| sh FTH1-2 | CCGGCCTGTCCATGTCTTACTACTTCTCGAGAAGTAGTAAGACATGGACAGGTTTTTG | AATTCAAAAACCTGTCCATGTCTTACTACTTCTCGAGAAGTAGTAAGACATGGACAGG | TRCN0000029432 |
| sh NC | CCGGGCAAGCTGACCCTGAAGTTCATCTCGAGATGAACTTCAGGGTCAGCTTGCTTTTTG | AATTCAAAAAGCAAGCTGACCCTGAAGTTCATCTCGAGATGAACTTCAGGGTCAGCTTGC |
| Name | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) |
|---|---|---|
| MYB | ATACATGAACGAGGAGCAGCG | TCATGGTCTGACTGTGGGAT |
| FTH1 | TGAAGCTGCAGAACCAACGAGG | GCACACTCCATTGCATTCAGCC |
| GPX4 | ACAAGAACGGCTGCGTGGTGAA | GCCACACACTTGTGGAGCTAGA |
| NRF2 | GCTCAAACTTAGGGGCTCCG | GAAGTTGCGGGAAGGTCTGG |
| MMP2 | ACCTGGATGCCGTCGGGAC | TGTGGCAGCACCAGGGCAGC |
| SNA11 | TCGGAAGCCTAACTACAGCGA | AGATGAGCATTGGCAGCGAG |
| CD133 | GCCACCGCTCTAGATACTGC | TGTTGTGATGGGCTTGTCAT |
| GAPDH | GGAAGGTGAAGGTCGGAGTCA | GTCATTGATGGCAACAATATCCACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, X.; Wang, Y.; Shen, S.; Fang, H.; Song, H.; Zhang, J.; Han, B. MYB Confers Sorafenib Resistance in Human Leukemia Cells via Inhibiting Ferroptosis Through FTH1 Upregulation. Genes 2025, 16, 737. https://doi.org/10.3390/genes16070737
Tao X, Wang Y, Shen S, Fang H, Song H, Zhang J, Han B. MYB Confers Sorafenib Resistance in Human Leukemia Cells via Inhibiting Ferroptosis Through FTH1 Upregulation. Genes. 2025; 16(7):737. https://doi.org/10.3390/genes16070737
Chicago/Turabian StyleTao, Xiaoxiao, Yucheng Wang, Siyu Shen, Huiying Fang, Hongkuan Song, Junfang Zhang, and Bingshe Han. 2025. "MYB Confers Sorafenib Resistance in Human Leukemia Cells via Inhibiting Ferroptosis Through FTH1 Upregulation" Genes 16, no. 7: 737. https://doi.org/10.3390/genes16070737
APA StyleTao, X., Wang, Y., Shen, S., Fang, H., Song, H., Zhang, J., & Han, B. (2025). MYB Confers Sorafenib Resistance in Human Leukemia Cells via Inhibiting Ferroptosis Through FTH1 Upregulation. Genes, 16(7), 737. https://doi.org/10.3390/genes16070737