
Integrated Transcriptome and Metabolome Analysis of the Porcine Small Intestine During Weaning


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling for Gene Expression Profiling and Metabolite Analysis
2.2. Gene Expression Profiling
2.3. Metabolite Analysis
2.4. Statistical Analysis
3. Results
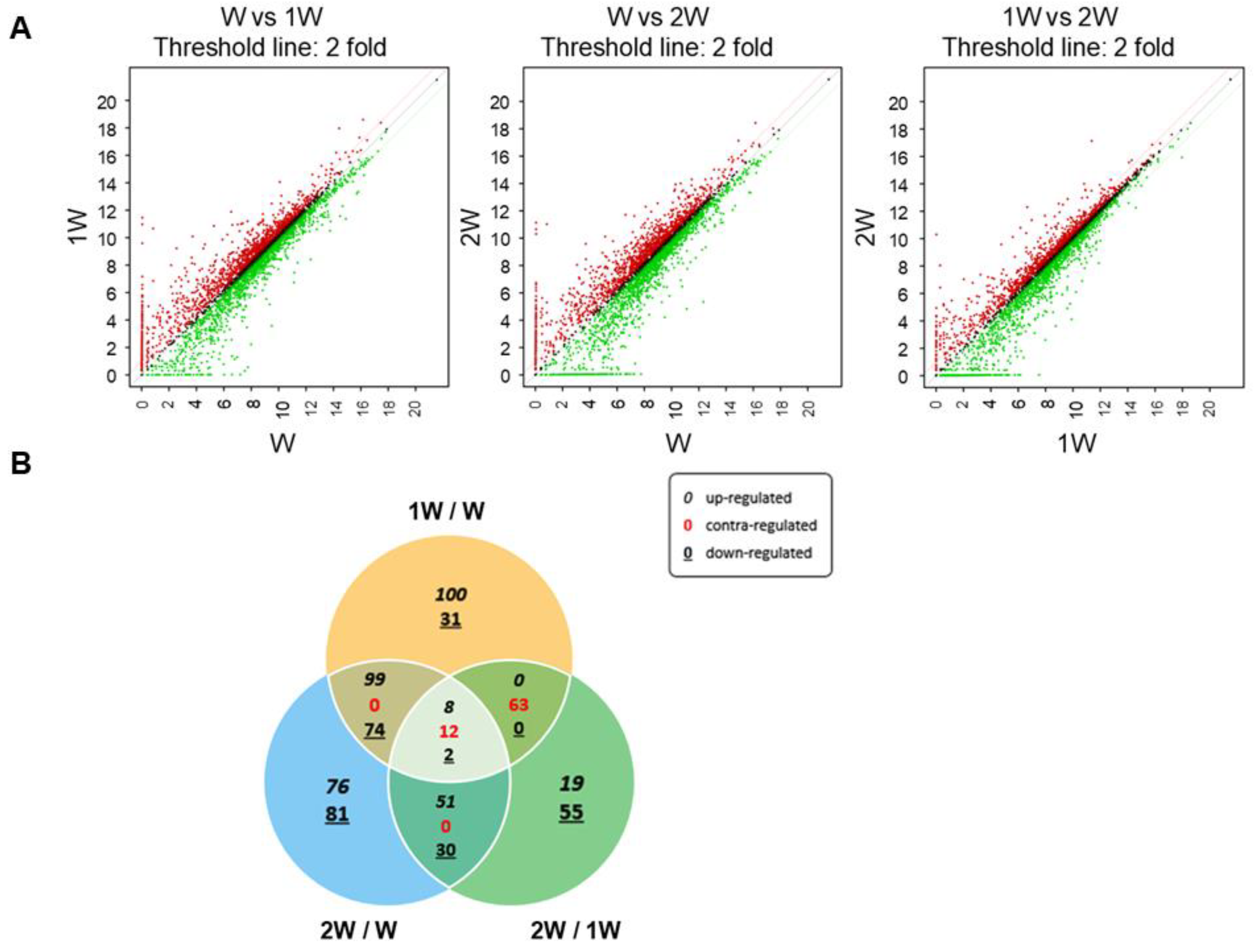
3.1. Differentially Expressed Genes in Pigs at Three Weaning Stages
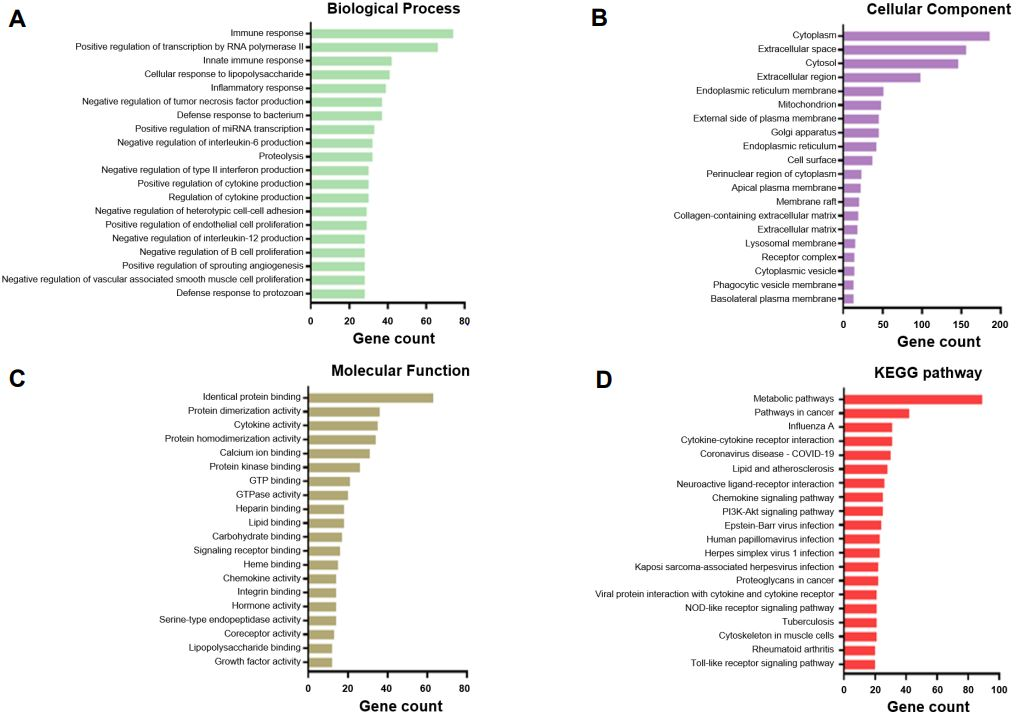
3.2. Functional Annotations of the Differentially Expressed Genes in the Three Weaning Stages
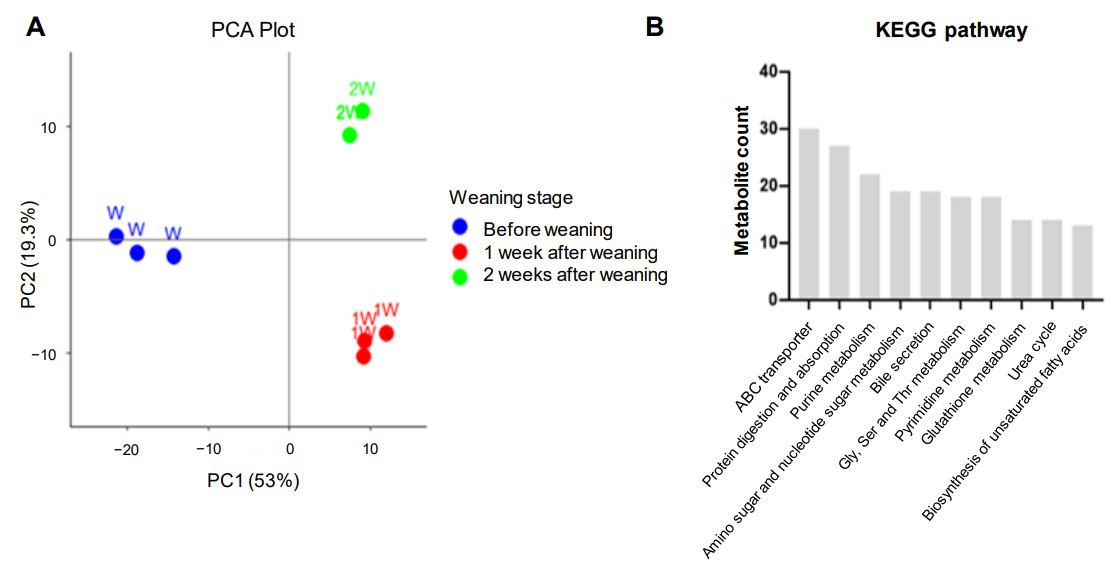
3.3. Alterations to the Porcine Intestine Metabolome During Weaning
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vicente, F.; Pereira, P.C. Pork Meat Composition and Health: A Review of the Evidence. Foods 2024, 13, 1905. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Gormley, A.; Jang, K.B.; Duarte, M.E. -Invited Review-Current status of global pig production: An overview and research trends. Anim. Biosci. 2024, 37, 719–729. [Google Scholar] [CrossRef]
- Knox, R.V. Impact of swine reproductive technologies on pig and global food production. Adv. Exp. Med. Biol. 2014, 752, 131–160. [Google Scholar] [CrossRef]
- Geng, J.; Huang, C.; Jiang, S. Roles and regulation of the matrix metalloproteinase system in parturition. Mol. Reprod. Dev. 2016, 83, 276–286. [Google Scholar] [CrossRef]
- Pearce, S.C.; Lonergan, S.M.; Huff-Lonergan, E.; Baumgard, L.H.; Gabler, N.K. Acute Heat Stress and Reduced Nutrient Intake Alter Intestinal Proteomic Profile and Gene Expression in Pigs. PLoS ONE 2015, 10, e0143099. [Google Scholar] [CrossRef] [PubMed]
- Maskal, J.M.; Duttlinger, A.W.; Kpodo, K.R.; McConn, B.R.; Byrd, C.J.; Richert, B.T.; Marchant-Forde, J.N.; Lay, D.C.; Perry, S.D.; Lucy, M.C.; et al. Evaluation and mitigation of the effects of in utero heat stress on piglet growth performance, postabsorptive metabolism, and stress response following weaning and transport. J. Anim. Sci. 2020, 98, skaa265. [Google Scholar] [CrossRef]
- Engelsmann, M.N.; Nielsen, T.S.; Hedemann, M.S.; Krogh, U.; Nørgaard, J.V. Effect of postweaning feed intake on performance, intestinal morphology, and the probability of diarrhoea in piglets. Anim. Int. J. Anim. Biosci. 2023, 17, 100891. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Yoon, J.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lee, S.H.; Park, Y.K.; Kwon, I.K.; Chae, B.J. Effects of dietary supplementation of antimicrobial peptide-A3 on growth performance, nutrient digestibility, intestinal and fecal microflora and intestinal morphology in weanling pigs. Anim. Feed Sci. Technol. 2012, 177, 98–107. [Google Scholar] [CrossRef]
- Lee, S.I.; Kim, I.H. Nucleotide-mediated SPDEF modulates TFF3-mediated wound healing and intestinal barrier function during the weaning process. Sci. Rep. 2018, 8, 4827. [Google Scholar] [CrossRef]
- Kang, T.H.; Shin, S.S.; Kim, T.H.; Lee, S.I. Ivangustin Alleviates Deoxynivalenol-Induced Apoptosis by Regulating FOXO3a Translocation in Porcine Intestinal Epithelial Cells. Toxins 2025, 17, 174. [Google Scholar] [CrossRef] [PubMed]
- Pácha, J. Development of intestinal transport function in mammals. Physiol. Rev. 2000, 80, 1633–1667. [Google Scholar] [CrossRef]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiong, X.; Wang, X.; Li, T.; Yin, Y. Effects of weaning on intestinal crypt epithelial cells in piglets. Sci. Rep. 2016, 6, 36939. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xiong, K.; Fang, R.; Li, M. Weaning stress and intestinal health of piglets: A review. Front. Immunol. 2022, 13, 1042778. [Google Scholar] [CrossRef]
- Ming, D.; Wang, W.; Huang, C.; Wang, Z.; Shi, C.; Ding, J.; Liu, H.; Wang, F. Effects of Weaning Age at 21 and 28 Days on Growth Performance, Intestinal Morphology and Redox Status in Piglets. Animals 2021, 11, 2169. [Google Scholar] [CrossRef]
- Huang, T.; Yang, M.; Dong, K.; Xu, M.; Liu, J.; Chen, Z.; Zhu, S.; Chen, W.; Yin, J.; Jin, K.; et al. A transcriptional landscape of 28 porcine tissues obtained by super deepSAGE sequencing. BMC Genom. 2020, 21, 229. [Google Scholar] [CrossRef]
- Wang, J.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. RNA sequencing (RNA-Seq) and its application in ovarian cancer. Gynecol. Oncol. 2019, 152, 194–201. [Google Scholar] [CrossRef]
- Pan, Z.; Yao, Y.; Yin, H.; Cai, Z.; Wang, Y.; Bai, L.; Kern, C.; Halstead, M.; Chanthavixay, G.; Trakooljul, N.; et al. Pig genome functional annotation enhances the biological interpretation of complex traits and human disease. Nat. Commun. 2021, 12, 5848. [Google Scholar] [CrossRef]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef]
- Herrmann, J.; Hermes, R.; Breves, G. Transepithelial transport and intraepithelial metabolism of short-chain fatty acids (SCFA) in the porcine proximal colon are influenced by SCFA concentration and luminal pH. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hui, P.; Uemoto, Y.; Ding, Y.; Yin, Z.; Bao, W. Metabolomic and Proteomic Profiling of Porcine Intestinal Epithelial Cells Infected with Porcine Epidemic Diarrhea Virus. Int. J. Mol. Sci. 2023, 24, 5071. [Google Scholar] [CrossRef] [PubMed]
- Toller-Kawahisa, J.E.; O’Neill, L.A.J. How neutrophil metabolism affects bacterial killing. Open Biol. 2022, 12, 220248. [Google Scholar] [CrossRef]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef]
- Tang, W.; Liu, J.; Ma, Y.; Wei, Y.; Liu, J.; Wang, H. Impairment of Intestinal Barrier Function Induced by Early Weaning via Autophagy and Apoptosis Associated With Gut Microbiome and Metabolites. Front. Immunol. 2021, 12, 804870. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kim, I.H. Eckol Alleviates Intestinal Dysfunction during Suckling-to-Weaning Transition via Modulation of PDX1 and HBEGF. Int. J. Mol. Sci. 2020, 21, 4755. [Google Scholar] [CrossRef] [PubMed]
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early weaning stress in pigs impairs innate mucosal immune responses to enterotoxigenic E. coli challenge and exacerbates intestinal injury and clinical disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef]
- Xu, S.S.; Wang, N.; Huang, L.; Zhang, X.L.; Feng, S.T.; Liu, S.S.; Wang, Y.; Liu, Z.G.; Wang, B.Y.; Wu, T.W.; et al. Changes in the Mucosa-Associated Microbiome and Transcriptome across Gut Segments Are Associated with Obesity in a Metabolic Syndrome Porcine Model. Microbiol. Spectr. 2022, 10, e0071722. [Google Scholar] [CrossRef]
- Yoon, J.W.; Kim, M.O.; Shin, S.; Kwon, W.S.; Kim, S.H.; Kwon, Y.J.; Lee, S.I. Spirobenzofuran Mitigates Ochratoxin A-Mediated Intestinal Adverse Effects in Pigs through Regulation of Beta Defensin 1. Toxics 2024, 12, 487. [Google Scholar] [CrossRef]
- Ding, S.; Cheng, Y.; Azad, M.A.K.; Zhu, Q.; Huang, P.; Kong, X. Developmental Changes of Immunity and Different Responses to Weaning Stress of Chinese Indigenous Piglets and Duroc Piglets during Suckling and Weaning Periods. Int. J. Mol. Sci. 2022, 23, 15781. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.; Blikslager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef]
- Tang, M.; Laarveld, B.; Van Kessel, A.G.; Hamilton, D.L.; Estrada, A.; Patience, J.F. Effect of segregated early weaning on postweaning small intestinal development in pigs. J. Anim. Sci. 1999, 77, 3191–3200. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning Stress Perturbs Gut Microbiome and Its Metabolic Profile in Piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.; Yang, C.; Nyachoti, C.M. Effects of sanitary conditions with lipopolysaccharide injection and dietary valine supplementation on growth performance, immune response, bacterial profile, and microbial metabolites in weaned pigs. Arch. Anim. Nutr. 2024, 78, 225–241. [Google Scholar] [CrossRef]
- Hou, L.; Cao, S.; Qiu, Y.; Xiong, Y.; Xiao, H.; Wen, X.; Yang, X.; Gao, K.; Wang, L.; Jiang, Z. Effects of early sub-therapeutic antibiotic administration on body tissue deposition, gut microbiota and metabolite profiles of weaned piglets. J. Sci. Food Agric. 2022, 102, 5913–5924. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Cheng, X.; Gu, L.; Zhou, J.; Zhu, X.; Zhang, X.; Guo, R.; Wang, W.; Li, B. Lipidomics reveals the significance and mechanism of the cellular ceramide metabolism for rotavirus replication. J. Virol. 2024, 98, e0006424. [Google Scholar] [CrossRef]
- Álvarez, B.; Revilla, C.; Poderoso, T.; Ezquerra, A.; Domínguez, J. Porcine Macrophage Markers and Populations: An Update. Cells 2023, 12, 2103. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiong, X.; Wang, X.; Tan, B.; Li, T.; Yin, Y. Effects of Weaning on Intestinal Upper Villus Epithelial Cells of Piglets. PLoS ONE 2016, 11, e0150216. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Zhu, L.; Chen, X.; Sheng, Y.; Guo, Q.; Bao, J.; Xu, J. X/XO or H2O2 induced IPEC-J2 cell as a new in vitro model for studying apoptosis in post-weaning piglets. Cytotechnology 2016, 68, 713–724. [Google Scholar] [CrossRef]
- Yan, S.; Long, L.; Zong, E.; Huang, P.; Li, J.; Li, Y.; Ding, X.; Xiong, X.; Yin, Y.; Yang, H. Dietary sulfur amino acids affect jejunal cell proliferation and functions by affecting antioxidant capacity, Wnt/β-catenin, and the mechanistic target of rapamycin signaling pathways in weaning piglets. J. Anim. Sci. 2018, 96, 5124–5133. [Google Scholar] [CrossRef] [PubMed]
- Magadum, A.; Engel, F.B. PPARβ/δ: Linking Metabolism to Regeneration. Int. J. Mol. Sci. 2018, 19, 2013. [Google Scholar] [CrossRef]
- Bhagwat, A.; Haldar, T.; Kanojiya, P.; Saroj, S.D. Bacterial metabolism in the host and its association with virulence. Virulence 2025, 16, 2459336. [Google Scholar] [CrossRef]
- Yang, B.; Guo, X.; Shi, C.; Liu, G.; Qin, X.; Chen, S.; Gan, L.; Liang, D.; Shao, K.; Xu, R.; et al. Alterations in purine and pyrimidine metabolism associated with latent tuberculosis infection: Insights from gut microbiome and metabolomics analyses. mSystems 2024, 9, e0081224. [Google Scholar] [CrossRef]
- Reiter, W.D. Biochemical genetics of nucleotide sugar interconversion reactions. Curr. Opin. Plant Biol. 2008, 11, 236–243. [Google Scholar] [CrossRef]
- Bekebrede, A.F.; Keijer, J.; Gerrits, W.J.J.; Boer, V.C.J. The Molecular and Physiological Effects of Protein-Derived Polyamines in the Intestine. Nutrients 2020, 12, 197. [Google Scholar] [CrossRef]
- Liu, X. ABC Family Transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar] [CrossRef]
- Su, X.; Li, X.; Wang, H.; Cai, Z. Simultaneous determination of methionine cycle metabolites, urea cycle intermediates and polyamines in serum, urine and intestinal tissue by using UHPLC-MS/MS. Talanta 2021, 224, 121868. [Google Scholar] [CrossRef] [PubMed]
- Mino, M.; Kakazu, E.; Sano, A.; Tsuruoka, M.; Matsubara, H.; Kakisaka, K.; Kogure, T.; Sekine, K.; Aoki, Y.; Imamura, M.; et al. Comprehensive analysis of peripheral blood free amino acids in MASLD: The impact of glycine-serine-threonine metabolism. Amino Acids 2024, 57, 3. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.W.; Shin, S.; Kim, T.H.; Lee, S.I. Integrated Transcriptome and Metabolome Analysis of the Porcine Small Intestine During Weaning. Genes 2025, 16, 727. https://doi.org/10.3390/genes16070727
Yoon JW, Shin S, Kim TH, Lee SI. Integrated Transcriptome and Metabolome Analysis of the Porcine Small Intestine During Weaning. Genes. 2025; 16(7):727. https://doi.org/10.3390/genes16070727
Chicago/Turabian StyleYoon, Jung Woong, Sangsu Shin, Tae Hyun Kim, and Sang In Lee. 2025. "Integrated Transcriptome and Metabolome Analysis of the Porcine Small Intestine During Weaning" Genes 16, no. 7: 727. https://doi.org/10.3390/genes16070727
APA StyleYoon, J. W., Shin, S., Kim, T. H., & Lee, S. I. (2025). Integrated Transcriptome and Metabolome Analysis of the Porcine Small Intestine During Weaning. Genes, 16(7), 727. https://doi.org/10.3390/genes16070727