
Incidence of Homozygous SMN2 Deletion in Japan: Cross-Reactivity of SMN2 Primers with SMN1 Sequence Causes False Negatives in Real-Time PCR Screening

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Residual DBS Samples
2.2. Real-Time PCR
2.3. PCR-Restriction Fragment Length Polymorphism (RFLP)
2.4. Nucleotide Sequencing
2.5. Statistical Analyses
3. Results
3.1. Detection of Homozygous SMN2 Deletion Using Real-Time PCR
3.2. Confirmation of the Real-Time PCR Results Using PCR-RFLP
3.3. Nucleotide Sequencing Analysis of the Real-Time PCR Amplified Products
3.4. Sensitivity and Specificity of the Screening, and Incidence of Homozygous SMN2 Deletion
4. Discussion
4.1. Establishment of a Screening System for Homozygous SMN2 Deletion
4.2. Incidence of Homozygous SMN2 Deletion in Japan
4.3. False Results Identified in the Present Study
4.4. Possible Pitfalls of Neonatal SMN1 Deletion Screening
4.5. Anthropological Importance of SMN2 Deletion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country/ Ethnicity | Controls | NeuromuscularDisorders | References | |||
|---|---|---|---|---|---|---|
| SMN2-Deletion/ Sample Number | Frequency | SMN2-Deletion/ Sample Number | Frequency | |||
| European | ||||||
| 1 | UK | (-) | (-) | 16/154 | 10.4% | [22] |
| 2 | USA and Canada | 4/54 | 7.4% | (-) | (-) | [23] |
| 3 | France | 8/90 | 8.9% | (-) | (-) | [24] |
| France | 15/167 | 9.0% | 16/167 | 9.6% | [25] | |
| 4 | France | 52/621 | 8.4% | 54/600 | 9.0% | [26] |
| 5 | Sweeden | 37/502 | 7.4% | 29/502 | 5.8% | [26] |
| 6 | Germany | 9/100 | 9.0% | (-) | (-) | [27] |
| 7 | The Netherlands | 78/984 | 7.9% | 62/847 | 7.3% | [28] |
| African | ||||||
| 8 | Sub-Saharan (Mali) | 150/613 | 24.5% | (-) | (-) | [21] |
| 9 | Sub-Saharan (Nigeria) | 33/120 | 27.5% | (-) | (-) | [21] |
| 10 | Sub-Saharan (Kenya) | 23/120 | 19.2% | (-) | (-) | [21] |
| 11 | Black South African | 15/122 | 12.3% | 60/122 | 49.2% | [11] |
| Asian | ||||||
| 12 | Vietnam | 2/52 | 3.9% | (-) | (-) | [29] |
| 13 | Chinese | 89/1712 | 5.29% | (-) | (-) | [30] |
| 14 | Taiwan | 30/520 | 5.8% | (-) | (-) | [31] |
| 15 | Taiwan | 5147/107,611 | 4.8% | (-) | (-) | [32] |
| 16 | Korea | 2/100 | 2.0% | 5/25 | 20.0% | [33] |
| 17 | Korea | 49/1581 | 3.1% | (-) | (-) | [34] |
| 18 | Japan | 16/300 | 5.3% | (-) | (-) | [12] |
| 19 | Japan | 19/399 | 4.8% | 41/537 | 7.6% | [35] |
| 20 | Japan | 49/1000 | 4.9% | (-) | (-) | This study |
- (1)
- Vorster et al. [11] noted that primates have only one copy of SMN1, suggesting that the SMN region in early humans might have consisted of only the SMN1 gene. At a later stage, the hypervariable nature of the SMN region might have resulted in multiple copies of SMN1. This duplicated SMN1 might have then diverged into SMN2 as a result of mutations. Thus, a high copy number of SMN1 and a lack of SMN2 may reflect a state before the divergence of SMN2 from SMN1.
- (2)
- Sangaré et al. [23] posited that the population that migrated out of Africa to Asia/Europe may have had a lower SMN1 copy number by chance, or might have randomly drifted in this direction after the migration. Black South African individuals may therefore represent the descendants who retained genetic diversity, whereas European/Asian individuals represent the descendants with reduced genetic diversity caused by the bottleneck phenomenon. Thus, the bottleneck phenomenon might be the driving force underlying the differences in SMA-related genotypes between Black South African and European/Asian individuals.
4.6. Perspectives from SMN2 Studies
4.7. Limitations
4.8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALS | Amyotrophic lateral sclerosis. |
| Ct | Cycle threshold. |
| DBS | Dried blood spot. |
| LMND | Lower motor neuron disease. |
| NBS | Newborn screening. |
| PCR | Polymerase chain reaction. |
| RFLP | Restriction fragment length polymorphism. |
| SD | Standard deviation. |
| SMA | Spinal muscular atrophy. |
| SMA-NBS | Newborn screening programs to detect SMN1 deletion. |
| SMN | Survival motor neuron. |
References
- Arnold, W.D.; Kassar, D.; Kissel, J.T. Spinal muscular atrophy: Diagnosis and management in a new therapeutic era. Muscle Nerve 2015, 51, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, S.; Bürglen, L.; Reboullet, S.; Clermont, O.; Burlet, P.; Viollet, L.; Benichou, B.; Cruaud, C.; Millasseau, P.; Zeviani, M.; et al. Identification and characterization of a spinal muscular atrophy-determining gene. Cell 1995, 80, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Verhaart, I.E.C.; Robertson, A.; Wilson, I.J.; Aartsma-Rus, A.; Cameron, S.; Jones, C.C.; Cook, S.F.; Lochmüller, H. Prevalence, incidence and carrier frequency of 5q-linked spinal muscular atrophy—A literature review. Orphanet J. Rare Dis. 2017, 12, 124. [Google Scholar] [CrossRef]
- Ito, M.; Yamauchi, A.; Urano, M.; Kato, T.; Matsuo, M.; Nakashima, K.; Saito, K. Epidemiological investigation of spinal muscular atrophy in Japan. Brain Dev. 2022, 44, 2–16. [Google Scholar] [CrossRef]
- Noguchi, Y.; Bo, R.; Nishio, H.; Matsumoto, H.; Matsui, K.; Yano, Y.; Sugawara, M.; Ueda, G.; Wijaya, Y.O.S.; Niba, E.T.E.; et al. PCR-based screening of spinal muscular atrophy for newborn infants in Hyogo Prefecture, Japan. Genes 2022, 13, 2110. [Google Scholar] [CrossRef]
- Nishio, H.; Niba, E.T.E.; Saito, T.; Okamoto, K.; Takeshima, Y.; Awano, H. Spinal muscular atrophy: The past, present, and future of diagnosis and treatment. Int. J. Mol. Sci. 2023, 26, 11939. [Google Scholar] [CrossRef] [PubMed]
- Feldkötter, M.; Schwarzer, V.; Wirth, R.; Wienker, T.F.; Wirth, B. Quantitative analyses of SMN1 and SMN2 based on real-time lightCycler PCR: Fast and highly reliable carrier testing and prediction of severity of spinal muscular atrophy. Am. J. Hum. Genet. 2002, 70, 358–368. [Google Scholar] [CrossRef]
- Moulard, B.; Salachas, F.; Chassande, B.; Briolotti, V.; Meininger, V.; Malafosse, A.; Camu, W. Association between centromeric deletions of the SMN gene and sporadic adult-onset lower motor neuron disease. Ann. Neurol. 1998, 43, 640–644. [Google Scholar] [CrossRef]
- Srivastava, S.; Mukherjee, M.; Panigrahi, I.; Shanker Pandey, G.; Pradhan, S.; Mittal, B. SMN2-deletion in childhood-onset spinal muscular atrophy. Am. J. Med. Genet. 2001, 101, 198–202. [Google Scholar] [CrossRef]
- Liping, L.; Hongwei, M.; Lin, W. Homozygous survival motor neuron 2 gene deletion and sporadic lower motor neuron disease in children: Case report and literature review. J. Child. Neurol. 2013, 28, 509–516. [Google Scholar] [CrossRef]
- Vorster, E.; Essop, F.B.; Rodda, J.L.; Krause, A. Spinal muscular atrophy in the Black South African population: A matter of rearrangement? Front. Genet. 2020, 11, 54. [Google Scholar] [CrossRef]
- Bouike, Y.; Sakima, M.; Taninishi, Y.; Matsutani, T.; Noguchi, Y.; Bo, R.; Awano, H.; Nishio, H. Real-time PCR-based screening for homozygous SMN2 deletion using residual dried blood spots. Genes 2023, 14, 2159. [Google Scholar] [CrossRef]
- Shinohara, M.; Niba, E.T.E.; Wijaya, Y.O.S.; Takayama, I.; Mitsuishi, C.; Kumasaka, S.; Kondo, Y.; Takatera, A.; Hokuto, I.; Morioka, I.; et al. A novel system for spinal muscular atrophy screening in newborns: Japanese pilot study. Int. J. Neonatal Screen. 2019, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Van der Steege, G.; Grootscholten, P.M.; Van der Vlies, P.; Draaijers, T.G.; Osinga, J.; Cobben, J.M.; Scheffer, H.; Buys, C.H. PCR-based DNA test to confirm clinical diagnosis of autosomal recessive spinal muscular atrophy. Lancet 1995, 345, 985–986. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhang, C.; Teng, Y.; Zeng, S.; Chen, S.; Liang, D.; Li, Z.; Wu, L. Detection of spinal muscular atrophy using a duplexed real-time PCR approach with locked nucleic acid-modified primers. Ann. Lab. Med. 2021, 41, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; Lee, F.K.; Yazdanpanah, G.K.; Staropoli, J.F.; Liu, M.; Carulli, J.P.; Sun, C.; Dobrowolski, S.F.; Hannon, W.H.; Vogt, R.F. Newborn blood spot screening test using multiplexed real-time PCR to simultaneously screen for spinal muscular atrophy and severe combined immunodeficiency. Clin. Chem. 2015, 61, 412–419. [Google Scholar] [CrossRef]
- Dangouloff, T.; Vrščaj, E.; Servais, L.; Osredkar, D.; SMA NBS World Study Group. Newborn screening programs for spinal muscular atrophy worldwide: Where we stand and where to go. Neuromuscul. Disord. 2021, 31, 574–582. [Google Scholar] [CrossRef]
- Cooper, K.; Nalbant, G.; Sutton, A.; Harnan, S.; Thokala, P.; Chilcott, J.; McNeill, A.; Bessey, A. Systematic review of newborn screening programmes for spinal muscular atrophy. Int. J. Neonatal Screen. 2024, 10, 49. [Google Scholar] [CrossRef]
- Rochette, C.F.; Gilbert, N.; Simard, L.R. SMN gene duplication and the emergence of the SMN2 gene occurred in distinct hominids: SMN2 is unique to Homo sapiens. Hum. Genet. 2001, 108, 255–266. [Google Scholar] [CrossRef]
- Blasco-Pérez, L.; Costa-Roger, M.; Leno-Colorado, J.; Bernal, S.; Alias, L.; Codina-Solà, M.; Martínez-Cruz, D.; Castiglioni, C.; Bertini, E.; Travaglini, L.; et al. Deep molecular characterization of milder spinal muscular atrophy patients carrying the c.859G>C variant in SMN2. Int. J. Mol. Sci. 2022, 23, 8289. [Google Scholar] [CrossRef]
- Sangaré, M.; Hendrickson, B.; Sango, H.A.; Chen, K.; Nofziger, J.; Amara, A.; Dutra, A.; Schindler, A.B.; Guindo, A.; Traoré, M.; et al. Genetics of low spinal muscular atrophy carrier frequency in sub-Saharan Africa. Ann. Neurol. 2014, 75, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Morrison, K.E.; Al-Chalabi, A.; Bakker, M.; Leigh, P.N. Analysis of chromosome 5q13 genes in amyotrophic lateral sclerosis: Homozygous NAIP deletion in a sporadic case. Ann. Neurol. 1996, 39, 796–800. [Google Scholar] [CrossRef] [PubMed]
- McAndrew, P.E.; Parsons, D.W.; Simard, L.R.; Rochette, C.; Ray, P.N.; Mendell, J.R.; Prior, T.W.; Burghes, A.H. Identification of proximal spinal muscular atrophy carriers and patients by analysis of SMNT and SMNC gene copy number. Am. J. Hum. Genet. 1997, 60, 1411–1422. [Google Scholar] [CrossRef]
- Gérard, B.; Ginet, N.; Matthijs, G.; Evrard, P.; Baumann, C.; Da Silva, F.; Gérard-Blanluet, M.; Mayer, M.; Grandchamp, B.; Elion, J. Genotype determination at the survival motor neuron locus in a normal population and SMA carriers using competitive PCR and primer extension. Hum. Mutat. 2000, 16, 253–263. [Google Scholar] [CrossRef]
- Corcia, P.; Camu, W.; Halimi, J.M.; Vourc’h, P.; Antar, C.; Vedrine, S.; Giraudeau, B.; de Toffol, B.; Andres, C.R.; French ALS Study Group. SMN1 gene, but not SMN2, is a risk factor for sporadic ALS. Neurology 2006, 67, 1147–1150. [Google Scholar] [CrossRef]
- Corcia, P.; Ingre, C.; Blasco, H.; Press, R.; Praline, J.; Antar, C.; Veyrat-Durebex, C.; Guettard, Y.O.; Camu, W.; Andersen, P.M.; et al. Homozygous SMN2 deletion is a protective factor in the Swedish ALS population. Eur. J. Hum. Genet. 2012, 20, 588–591. [Google Scholar] [CrossRef]
- Anhuf, D.; Eggermann, T.; Rudnik-Schöneborn, S.; Zerres, K. Determination of SMN1 and SMN2 copy number using TaqMan technology. Hum. Mutat. 2003, 22, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Blauw, H.M.; Barnes, C.P.; Van Vught, P.W.; Van Rheenen, W.; Verheul, M.; Cuppen, E.; Veldink, J.H.; Van den Berg, L.H. SMN1 gene duplications are associated with sporadic ALS. Neurology 2012, 78, 776–780. [Google Scholar] [CrossRef]
- Tran, V.K.; Sasongko, T.H.; Hong, D.D.; Hoan, N.T.; Dung, V.C.; Lee, M.J.; Gunadi Takeshima, Y.; Matsuo, M.; Nishio, H. SMN2 and NAIP gene dosages in Vietnamese patients with spinal muscular atrophy. Pediatr. Int. 2008, 50, 346–351. [Google Scholar] [CrossRef]
- Sheng-Yuan, Z.; Xiong, F.; Chen, Y.J.; Yan, T.Z.; Zeng, J.; Li, L.; Zhang, Y.N.; Chen, W.Q.; Bao, X.H.; Zhang, C.; et al. Molecular characterization of SMN copy number derived from carrier screening and from core families with SMA in a Chinese population. Eur. J. Hum. Genet. 2010, 18, 978–984. [Google Scholar] [CrossRef]
- Kao, H.Y.; Su, Y.N.; Liao, H.K.; Liu, M.S.; Chen, Y.J. Determination of SMN1/SMN2 gene dosage by a quantitative genotyping platform combining capillary electrophoresis and MALDI-TOF mass spectrometry. Clin. Chem. 2006, 52, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.N.; Hung, C.C.; Lin, S.Y.; Chen, F.Y.; Chern, J.P.; Tsai, C.; Chang, T.S.; Yang, C.C.; Li, H.; Ho, H.N.; et al. Carrier screening for spinal muscular atrophy (SMA) in 107,611 pregnant women during the period 2005–2009: A prospective population-based cohort study. PLoS ONE 2011, 6, 17067. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Lee, K.A.; Hong, J.M.; Suh, G.I.; Choi, Y.C. Homozygous SMN2 deletion is a major risk factor among twenty-five Korean sporadic amyotrophic lateral sclerosis patients. Yonsei Med. J. 2012, 53, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Yun, S.A.; Roh, E.Y.; Yoon, J.H.; Shin, S.; Ki, C.S. Carrier frequency of spinal muscular atrophy in a large-scale Korean population. Ann. Lab. Med. 2020, 40, 326–330. [Google Scholar] [CrossRef]
- Ishihara, T.; Koyama, A.; Atsuta, N.; Tada, M.; Toyoda, S.; Kashiwagi, K.; Hirokawa, S.; Hatano, Y.; Yokoseki, A.; Nakamura, R.; et al. SMN2 gene copy number affects the incidence and prognosis of motor neuron diseases in Japan. BMC Med. Genom. 2024, 17, 263. [Google Scholar] [CrossRef] [PubMed]
- Hsieh-Li, H.M.; Chang, J.G.; Jong, Y.J.; Wu, M.H.; Wang, N.M.; Tsai, C.H.; Li, H. A mouse model for spinal muscular atrophy. Nat. Genet. 2000, 24, 66–70. [Google Scholar] [CrossRef]
- Monani, U.R.; Sendtner, M.; Coovert, D.D.; Parsons, D.W.; Andreassi, C.; Le, T.T.; Jablonka, S.; Schrank, B.; Rossoll, W.; Prior, T.W.; et al. The human centromeric survival motor neuron gene (SMN2) rescues embryonic lethality in Smn−/− mice and results in a mouse with spinal muscular atrophy. Hum. Mol. Genet. 2000, 9, 333–339. [Google Scholar] [CrossRef]
- Echaniz-Laguna, A.; Guiraud-Chaumeil, C.; Tranchant, C.; Reeber, A.; Melki, J.; Warter, J.M. Homozygous exon 7 deletion of the SMN centromeric gene (SMN2): A potential susceptibility factor for adult-onset lower motor neuron disease. J. Neurol. 2002, 249, 290–293. [Google Scholar] [CrossRef]
- Gamez, J.; Barceló, M.J.; Muñoz, X.; Carmona, F.; Cuscó, I.; Baiget, M.; Cervera, C.; Tizzano, E.F. Survival and respiratory decline are not related to homozygous SMN2 deletions in ALS patients. Neurology 2002, 59, 1456–1460. [Google Scholar] [CrossRef]



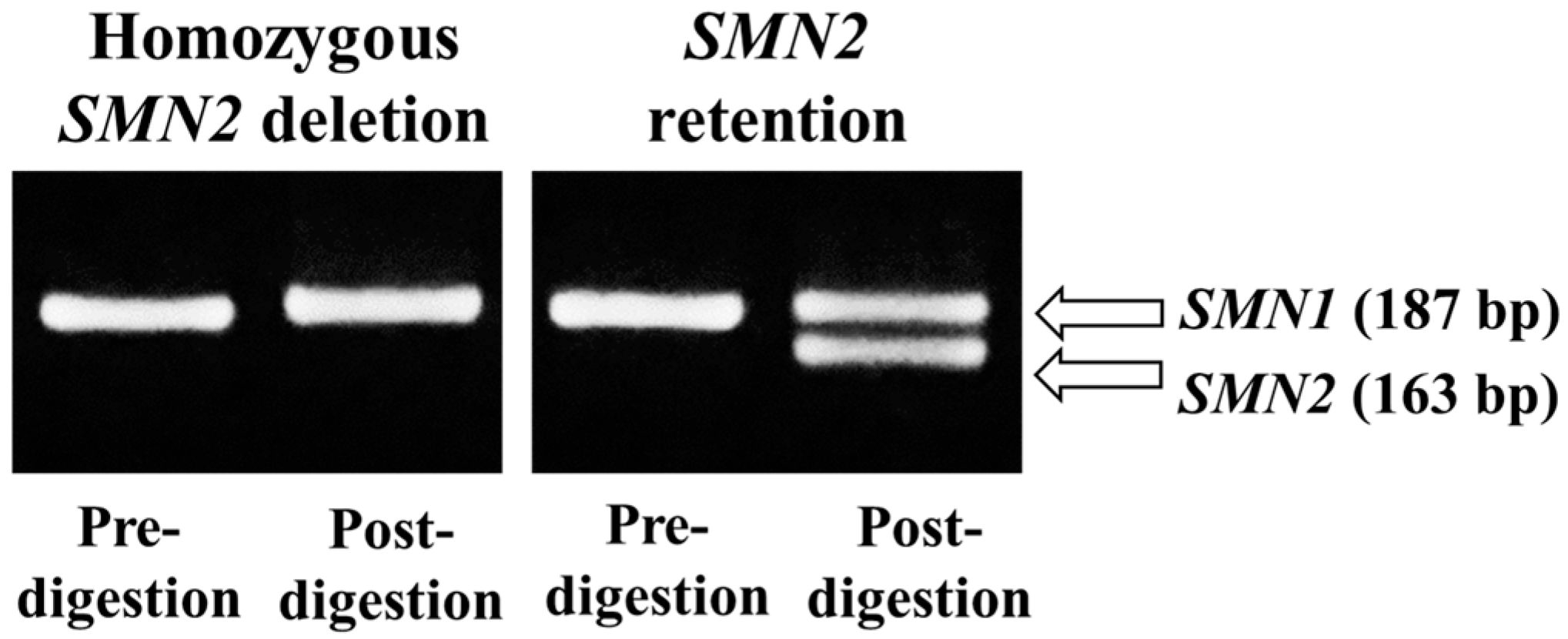
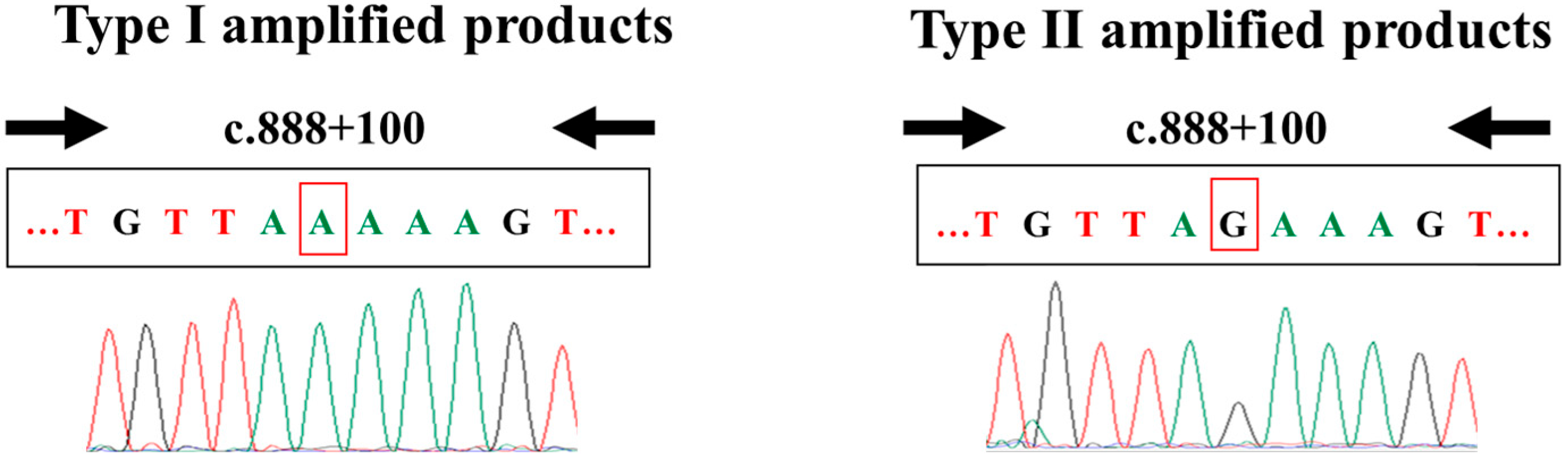
| Screening Assay by Real-Time PCR | Confirmatory Assay by PCR-RFLP | ||
|---|---|---|---|
| Homozygous SMN2 Deletion | SMN2 Retention | Total | |
| Ct ≥ mean + 2SD (Group A) | 45 (TP) | 6 (FP) | 51 |
| Ct ≥ mean + 1SD and Ct < mean + 2SD (Group B) | 4 (FN) | 42 (TN) | 46 |
| Total | 49 | 48 | 97 |
| Screening Assay by Real-Time PCR | Nucleotide Sequencing | ||
|---|---|---|---|
| Type I (with A) | Type II (with G) | Total | |
| Ct ≥ mean + 2SD (Group A) | 45 (TP) | 6 (FP) | 51 |
| Ct ≥ mean + 1SD and <mean + 2SD (Group B) | 4 (FN) | 42 (TN) | 46 |
| Total | 49 | 48 | 97 |
| Screening Assay | Homozygous SMN2 Deletion | SMN2 Retention | Total | |
|---|---|---|---|---|
| Screen-positive (Group A) | 45 (TP) | 6 (FP) | 51 | PPV 0.882 |
| Screen-negative (Groups B and C) | 4 (FN) | 945 (TN) * | 949 | NPV 0.996 |
| Total | 49 | 951 | 1000 | |
| Sensitivity 0.918 | Specificity 0.994 |
| Homozygous SMN2 Deletion | SMN2 Retention | Total | |
|---|---|---|---|
| Case number | 49 | 951 | 1000 |
| Percentage | 4.9 | 95.1 | 100.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakima, M.; Bouike, Y.; Wada, S.-I.; Nakamae, M.; Noguchi, Y.; Bo, R.; Awano, H.; Oba, J.; Nishio, H. Incidence of Homozygous SMN2 Deletion in Japan: Cross-Reactivity of SMN2 Primers with SMN1 Sequence Causes False Negatives in Real-Time PCR Screening. Genes 2025, 16, 712. https://doi.org/10.3390/genes16060712
Sakima M, Bouike Y, Wada S-I, Nakamae M, Noguchi Y, Bo R, Awano H, Oba J, Nishio H. Incidence of Homozygous SMN2 Deletion in Japan: Cross-Reactivity of SMN2 Primers with SMN1 Sequence Causes False Negatives in Real-Time PCR Screening. Genes. 2025; 16(6):712. https://doi.org/10.3390/genes16060712
Chicago/Turabian StyleSakima, Makoto, Yoshihiro Bouike, Shin-Ichi Wada, Masami Nakamae, Yoriko Noguchi, Ryosuke Bo, Hiroyuki Awano, Jumpei Oba, and Hisahide Nishio. 2025. "Incidence of Homozygous SMN2 Deletion in Japan: Cross-Reactivity of SMN2 Primers with SMN1 Sequence Causes False Negatives in Real-Time PCR Screening" Genes 16, no. 6: 712. https://doi.org/10.3390/genes16060712
APA StyleSakima, M., Bouike, Y., Wada, S.-I., Nakamae, M., Noguchi, Y., Bo, R., Awano, H., Oba, J., & Nishio, H. (2025). Incidence of Homozygous SMN2 Deletion in Japan: Cross-Reactivity of SMN2 Primers with SMN1 Sequence Causes False Negatives in Real-Time PCR Screening. Genes, 16(6), 712. https://doi.org/10.3390/genes16060712