
Comparative Genomic and Phylogenetic Analysis of Chloroplasts in Citrus paradisi Mac.cv. Cocktail

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. Assembly and Annotation of Chloroplast Gene Sequences
2.3. Analysis of Simple Sequence Repeats (SSR)
2.4. Analysis of Codon Usage Bias
2.5. Comparative Analysis
2.6. Analysis of Selective Pressures in the Evolution
2.7. Phylogenetic Analysis
3. Results
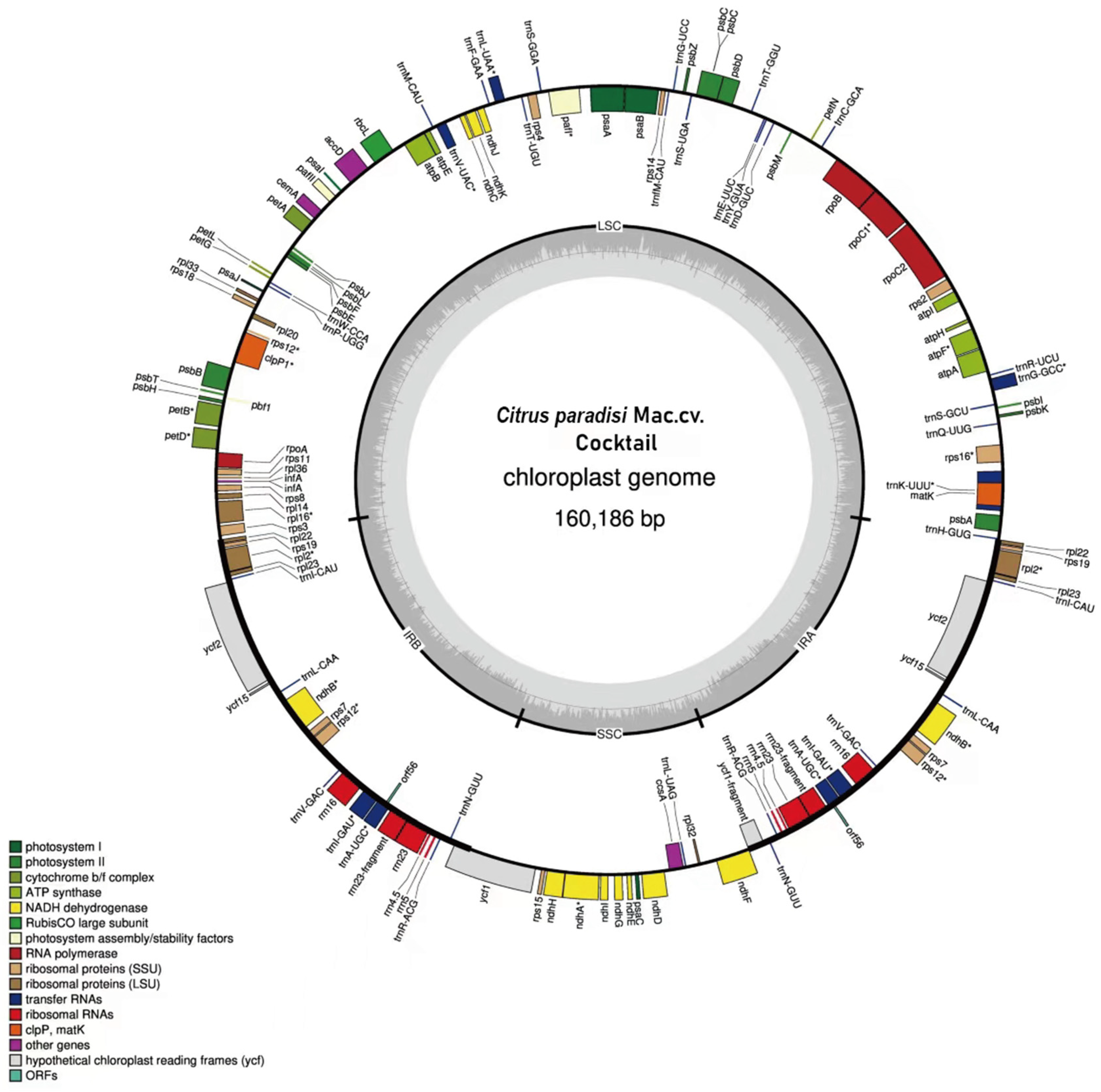
3.1. Characterization of Chloroplast Genomes
3.2. Analysis of Simple Sequence Repeats (SSRs)
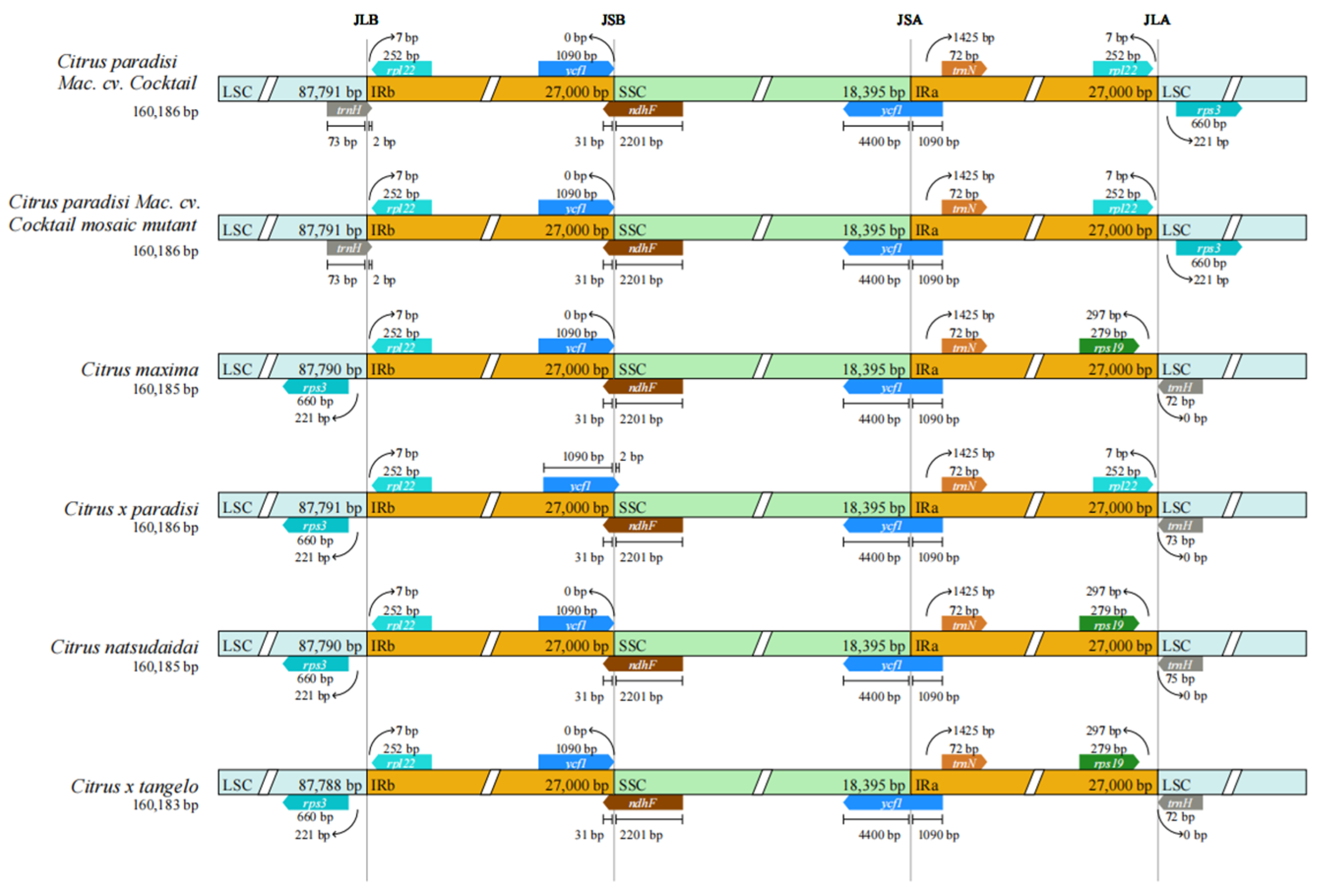
3.3. IRs Boundary Analysis
3.4. Codon Usage Analysis
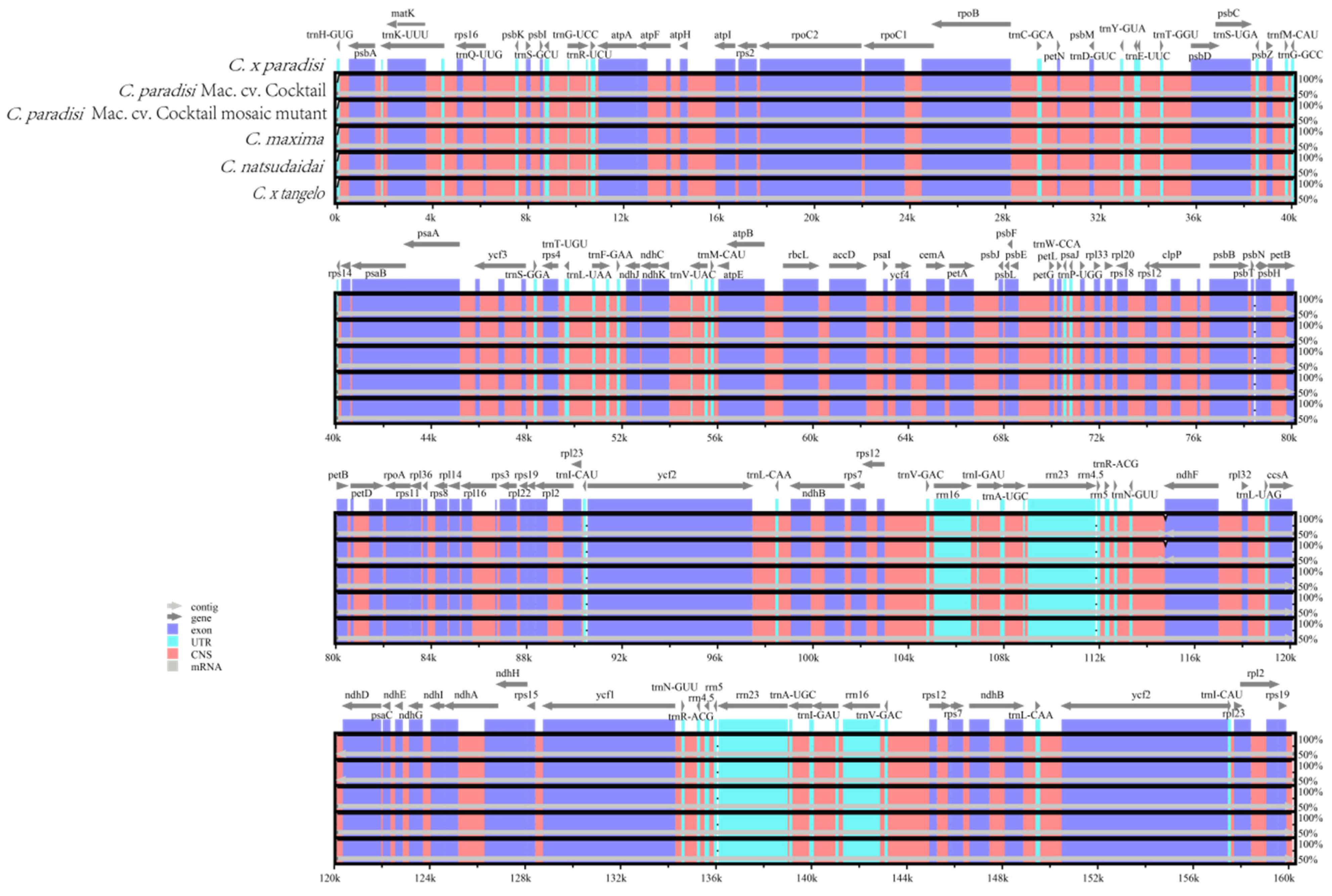
3.5. Genome Comparison
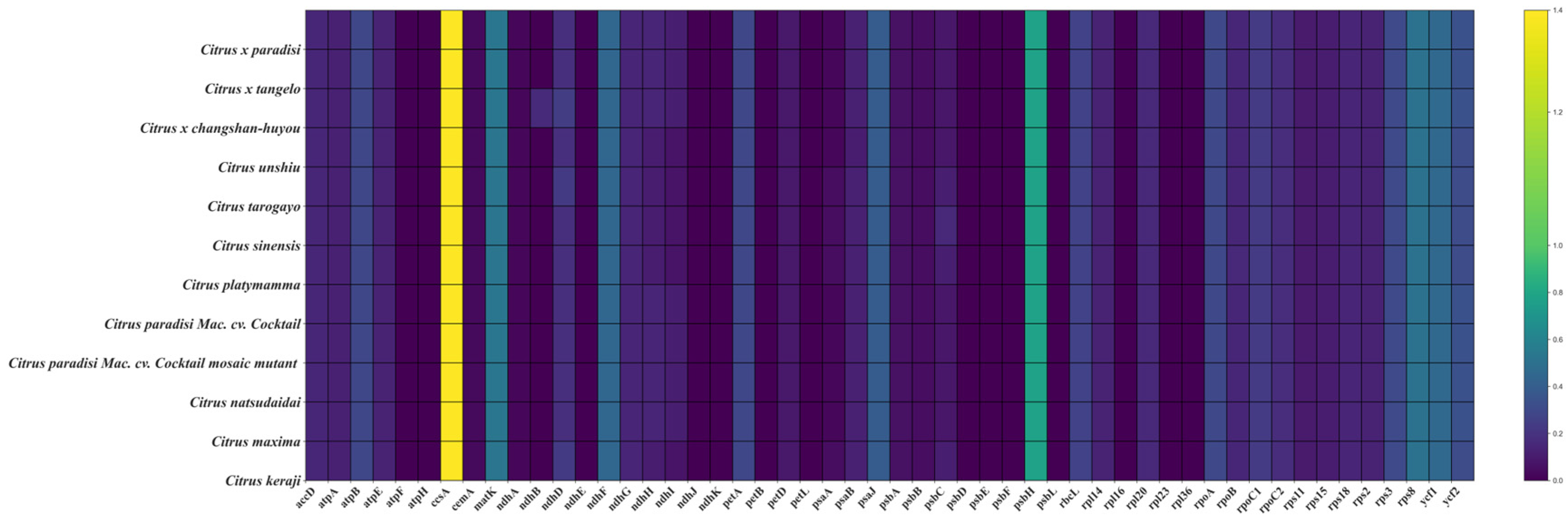
3.6. Selective Pressures in the Evolution
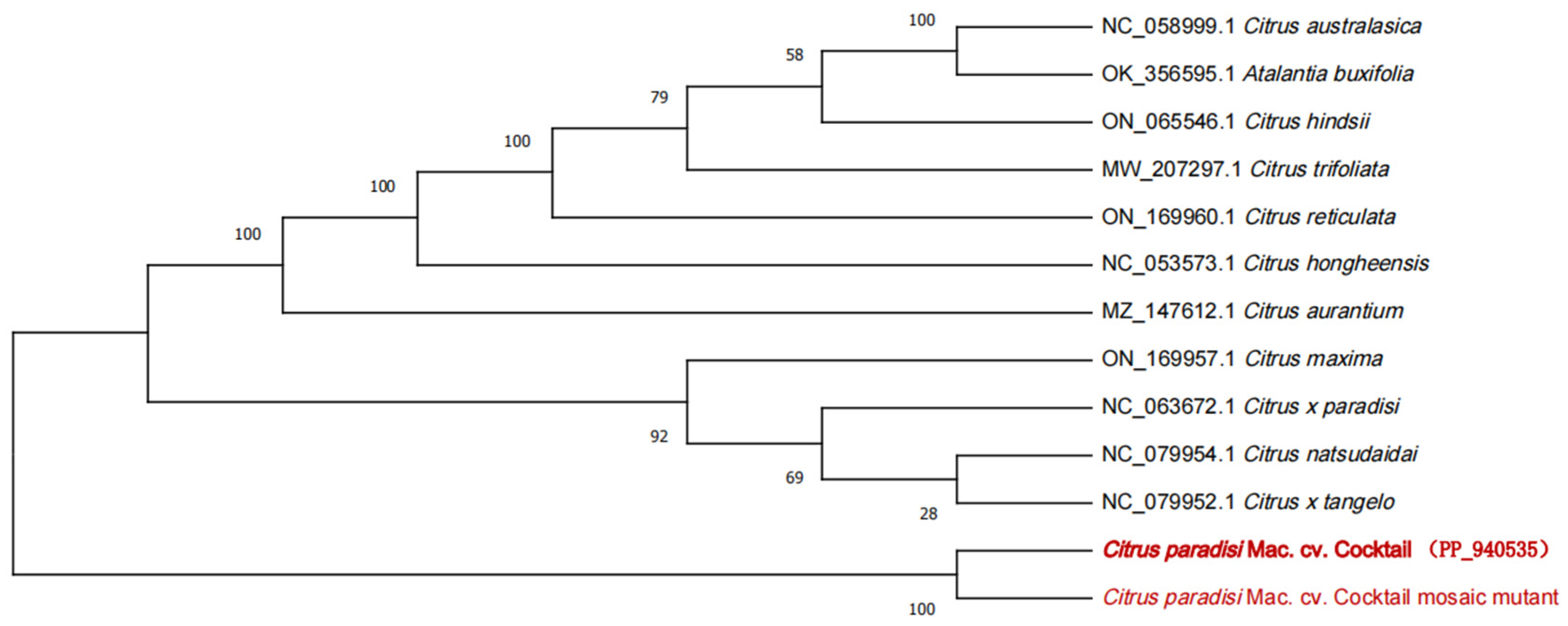
3.7. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xi, W.P.; Zhang, G.W.; Jiang, D.; Zhou, Z.Q. Phenolic compositions and antioxidant activities of grapefruit (Citrus paradisi Macfadyen) varieties cultivated in China. Int. J. Food Sci. Nutr. 2015, 66, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Xu, C.N.; Gao, X.Y.; Zhang, W.Q.; Yao, Z.L.; Wang, T.Y.; Feng, X.J.; Wang, Y. Comparative study on secondary metabolites from different citrus varieties in the production area of Zhejiang. Front. Nutr. 2023, 10, 1159676. [Google Scholar] [CrossRef] [PubMed]
- Castro-Vazquez, L.; Alañón, M.E.; Rodríguez-Robledo, V.; Pérez-Coello, M.S.; Hermosín-Gutierrez, I.; Díaz-Maroto, M.C.; Jordán, J.; Galindo, M.F.; Arroyo-Jiménez, M.d.M.; Delic, D. Bioactive Flavonoids, Antioxidant Behaviour, and Cytoprotective Effects of Dried Grapefruit Peels (Citrus paradisi Macf.). Oxidative Med. Cell. Longev. 2016, 2016, 8915729. [Google Scholar] [CrossRef] [PubMed]
- Arab, M.M.; Brown, P.J.; Abdollahi-Arpanahi, R.; Sohrabi, S.S.; Askari, H.; Aliniaeifard, S.; Mokhtassi-Bidgoli, A.; Mesgaran, M.B.; Leslie, C.A.; Marrano, A.; et al. Genome-wide association analysis and pathway enrichment provide insights into the genetic basis of photosynthetic responses to drought stress in Persian walnut. Hortic. Res. 2022, 9, uhac124. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Y.; Lv, J.; Xu, J.; Zhu, S.; Li, M.; Chen, N. Development of Chloroplast Genomic Resources for Oryza Species Discrimination. Front. Plant Sci. 2017, 8, 1854. [Google Scholar] [CrossRef]
- Ivanova, Z.; Sablok, G.; Daskalova, E.; Zahmanova, G.; Apostolova, E.; Yahubyan, G.; Baev, V. Chloroplast Genome Analysis of Resurrection Tertiary Relict Haberlea rhodopensis Highlights Genes Important for Desiccation Stress Response. Front. Plant Sci. 2017, 8, 204. [Google Scholar] [CrossRef]
- Roy, P.S.; Rao, G.J.N.; Jena, S.; Samal, R.; Patnaik, A.; Patnaik, S.S.C.; Jambhulkar, N.N.; Sharma, S.; Mohapatra, T. Nuclear and Chloroplast DNA Variation Provides Insights into Population Structure and Multiple Origin of Native Aromatic Rices of Odisha, India. PLoS ONE 2016, 11, e0162268. [Google Scholar] [CrossRef]
- Song, W.C.; Chen, Z.M.; He, L.; Feng, Q.; Zhang, H.R.; Du, G.L.; Shi, C.; Wang, S. Comparative Chloroplast Genome Analysis of Wax Gourd (Benincasa hispida) with Three Benincaseae Species, Revealing Evolutionary Dynamic Patterns and Phylogenetic Implications. Genes 2022, 13, 461. [Google Scholar] [CrossRef]
- Shi, W.B.; Huang, Y.H.; Hu, S.Q.; Wang, H.; Lin, Q.H.; Zhang, Z.R.; Shi, C.; Wang, S. Exploring the chloroplast genomics, comparative analysis, evolution, and phylogenetic relationships of Phylica pubescens (Rhamnaceae) in the Cape Flora. S. Afr. J. Bot. 2024, 164, 374–385. [Google Scholar] [CrossRef]
- Shi, W.B.; Song, W.C.; Liu, J.; Shi, C.; Wang, S. Comparative chloroplast genome analysis of Citrus (Rutaceae) species: Insights into genomic characterization, phylogenetic relationships, and discrimination of subgenera. Sci. Hortic. 2023, 313, 14. [Google Scholar] [CrossRef]
- Hu, J.; Yao, J.; Lu, J.; Liu, W.; Zhao, Z.; Li, Y.; Jiang, L.; Zha, L. The complete chloroplast genome sequences of nine melon varieties (Cucumis melo L.): Lights into comparative analysis and phylogenetic relationships. Front. Genet. 2024, 15, 1417266. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Tian, C.Y.; Hu, H.H.; Yang, Y.T.; Ma, H.L.; Liu, Q.; Liu, L.M.; Li, Z.Y.; Wu, Z.N. Characterization and Comparative Analysis of Complete Chloroplast Genomes of Four Bromus (Poaceae, Bromeae) Species. Genes 2024, 15, 815. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Duan, B.Z.; Zhou, Z.Y.; Fang, H.; Yang, M.H.; Xia, C.L.; Zhou, Y.; Wang, J. Comparative analysis of medicinal plants Scutellaria baicalensis and common adulterants based on chloroplast genome sequencing. BMC Genom. 2024, 25, 39. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zhao, Y.; Wang, X.R.; Jian, X.Y. Characterization of the chloroplast genome of Gleditsia species and comparative analysis. Sci. Rep. 2024, 14, 4262. [Google Scholar] [CrossRef]
- Shi, W.B.; Song, W.C.; Zhao, Y.Q.; Shi, C.; Wang, S. Complete chloroplast genomes of four Atalantia (Rutaceae) species: Insights into comparative analysis, phylogenetic relationships, and divergence time estimation. Plant Syst. Evol. 2023, 309, 17. [Google Scholar] [CrossRef]
- Kan, J.; Nie, L.; Wang, M.; Tiwari, R.; Tembrock, L.R.; Wang, J. The Mendelian pea pan-plastome: Insights into genomic structure, evolutionary history, and genetic diversity of an essential food crop. Genom. Commun. 2024, 1, e004. [Google Scholar] [CrossRef]
- Moosavi, S.J.; Mueller, M.; Gailing, O. Development of New Chloroplast Microsatellites for Pinus gerardiana and their Application in Genetic Diversity Analyses. Ecol. Evol. 2025, 15, e71185. [Google Scholar] [CrossRef]
- Ren, W.C.; Liu, C.; Yan, S.; Jiang, Z.H.; Wang, T.H.; Wang, Z.; Zhang, M.Q.; Liu, M.Q.; Sun, J.I.; Gao, J.H.; et al. Structural Characterization of the Acer ukurunduense Chloroplast Genome Relative to Related Species in the Acer Genus. Front. Genet. 2022, 13, 15. [Google Scholar] [CrossRef]
- Lin, J.X.; Lin, Z.Y.; Chen, Y.Q.; Xu, H.B. The complete chloroplast genome sequence of Lemna turionifera (Araceae). Mitochondrial DNA Part B—Resour. 2024, 9, 971–975. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; dePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Ebert, D.; Peakall, R.O.D. Chloroplast simple sequence repeats (cpSSRs): Technical resources and recommendations for expanding cpSSR discovery and applications to a wide array of plant species. Mol. Ecol. Resour. 2009, 9, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Beier, S.; Thiel, T.; Muench, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Parvathy, S.T.; Udayasuriyan, V.; Bhadana, V. Codon usage bias. Mol. Biol. Rep. 2021, 49, 539–565. [Google Scholar] [CrossRef]
- Sharp, P.M.; Tuohy, T.M.F.; Mosurski, K.R. Codon usage in yeast: Cluster analysis clearly differentiates highly and lowly expressed genes. Nucleic Acids Res. 1986, 14, 5125–5143. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Li, H.E.; Guo, Q.Q.; Xu, L.; Gao, H.D.; Liu, L.; Zhou, X.Y. CPJSdraw: Analysis and visualization of junction sites of chloroplast genomes. PeerJ 2023, 11, e15326. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11 Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Lin, H. The complete chloroplast genome and phylogenetic analysis of Citrus clementina (Rutaceae). Mitochondrial DNA Part B 2021, 6, 2926–2927. [Google Scholar] [CrossRef]
- Zhu, Y.Y.; Zhang, X.W.; Yan, S.F.; Feng, C.; Wang, D.F.; Yang, W.; Daud, M.K.; Xiang, J.Q.; Mei, L. SSR identification and phylogenetic analysis in four plant species based on complete chloroplast genome sequences. Plasmid 2023, 125, 102670. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Sharma, A.; Sharma, V.; Gaikwad, P.N.; Sidhu, G.S.; Kaur, G.; Kaur, N.; Jindal, T.; Chhuneja, P.; Rattanpal, H.S. Comprehensive genome-wide identification and transferability of chromosome-specific highly variable microsatellite markers from citrus species. Sci. Rep. 2023, 13, 10919. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.G.; Wu, H.K.; Luo, Y.; Xi, W.P.; Zhou, Z.Q. Comparative analysis of chloroplast genomes of the genus Citrus and its close relatives. Mitochondrial DNA Part A 2017, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Kong, W.-P.; Zhang, M.-M.; Hou, D. The complete chloroplast genome of Cucumis anguria var. anguria (Cucurbitaceae) and its phylogenetic implication. Mitochondrial DNA Part B 2020, 5, 654–655. [Google Scholar] [CrossRef]
- Park, I.; Yang, S.; Choi, G.; Kim, W.; Moon, B. The Complete Chloroplast Genome Sequences of Aconitum pseudolaeve and Aconitum longecassidatum, and Development of Molecular Markers for Distinguishing Species in the Aconitum Subgenus Lycoctonum. Molecules 2017, 22, 2012. [Google Scholar] [CrossRef]
- Zalapa, J.E.; Cuevas, H.; Zhu, H.Y.; Steffan, S.; Senalik, D.; Zeldin, E.; McCown, B.; Harbut, R.; Simon, P. Using Next-Generation Sequencing Approaches to Isolate Simple Sequence Repeat (SSR) Loci in the Plant Sciences. Am. J. Bot. 2012, 99, 193–208. [Google Scholar] [CrossRef]
- Yi, S.; Li, Y.; Wang, W. Selection shapes the patterns of codon usage in three closely related species of genus Misgurnus. Genomics 2018, 110, 134–142. [Google Scholar] [CrossRef]
- Wu, Z.; Liao, R.; Yang, T.; Dong, X.; Lan, D.; Qin, R.; Liu, H. Analysis of six chloroplast genomes provides insight into the evolution of Chrysosplenium (Saxifragaceae). BMC Genom. 2020, 21, 621. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, P.; Zeng, Q.; Zeng, R.; Hu, W.; Sun, L.; Liu, S.; Luan, F.; Zhu, Q. Comparative chloroplast genome analysis of seven extant Citrullus species insight into genetic variation, phylogenetic relationships, and selective pressure. Sci. Rep. 2023, 13, 6779. [Google Scholar] [CrossRef]
- Andrade-Marcial, M.; Pacheco-Arjona, R.; Hernandez-Castellano, S.; Che-Aguilar, L.; De-la-Pena, C. Transcriptome analysis reveals molecular mechanisms underlying chloroplast biogenesis in albino Agave angustifolia plantlets. Physiol. Plant. 2024, 176, e14289. [Google Scholar] [CrossRef]
- Li, N.N.; Lu, J.L.; Li, Q.S.; Zheng, X.Q.; Wang, X.C.; Wang, L.; Wang, Y.C.; Ding, C.Q.; Liang, Y.R.; Yang, Y.J. Dissection of Chemical Composition and Associated Gene Expression in the Pigment-Deficient Tea Cultivar ‘Xiaoxueya’ Reveals an Albino Phenotype and Metabolite Formation. Front. Plant Sci. 2019, 10, 1543. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSR Type (Number of Copies) | SSR Sequence | Number of Copies | Total Number | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 03 | 04 | 05 | 06 | 07 | 08 | 09 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | |||
| Mononucleotide (78) | A/T | - | - | - | - | - | - | - | 18 | 16 | 14 | 10 | 6 | 6 | - | 3 | 3 | - | - | 1 | 77 |
| C/G | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Trinucleotide (1) | AAT/ATT | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Tetranucleotide (2) | AAAT/ATTT | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Pentanucleotide (1) | AAAAT/ATTTT | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Amino Acid | Codon | Number | RSCU | Amino Acid | Codon | Number | RSCU |
|---|---|---|---|---|---|---|---|
| Ter | UAA | 1111 | 1.1929 | Ter | UAA | 1158 | 1.1634 |
| UAG | 750 | 0.8053 | UAG | 839 | 0.8429 | ||
| UGA | 933 | 1.0018 | UGA | 989 | 0.9936 | ||
| Ala | GCA | 490 | 1.0925 | Ala | GCA | 470 | 1.1171 |
| GCC | 430 | 0.9588 | GCC | 386 | 0.9174 | ||
| GCG | 318 | 0.7090 | GCG | 310 | 0.7368 | ||
| GCU | 556 | 1.2397 | GCU | 517 | 1.2288 | ||
| Cys | UGC | 456 | 0.8057 | Cys | UGC | 486 | 0.8541 |
| UGU | 676 | 1.1943 | UGU | 652 | 1.1459 | ||
| Asp | GAC | 409 | 0.5550 | Asp | GAC | 450 | 0.5810 |
| GAU | 1065 | 1.4450 | GAU | 1099 | 1.4190 | ||
| Glu | GAA | 1432 | 1.3978 | Glu | GAA | 1429 | 1.3707 |
| GAG | 617 | 0.6022 | GAG | 656 | 0.6293 | ||
| Phe | UUC | 1435 | 0.7990 | Phe | UUC | 1481 | 0.7884 |
| UUU | 2157 | 1.2010 | UUU | 2276 | 1.2116 | ||
| Gly | GGA | 899 | 1.3565 | Gly | GGA | 910 | 1.4577 |
| GGC | 401 | 0.6051 | GGC | 363 | 0.5815 | ||
| GGG | 714 | 1.0773 | GGG | 625 | 1.0012 | ||
| GGU | 637 | 0.9611 | GGU | 599 | 0.9596 | ||
| His | CAC | 410 | 0.6274 | His | CAC | 425 | 0.6574 |
| CAU | 897 | 1.3726 | CAU | 868 | 1.3426 | ||
| Ile | AUA | 1441 | 0.9763 | Ile | AUA | 1347 | 0.9596 |
| AUC | 1139 | 0.7717 | AUC | 1053 | 0.7502 | ||
| AUU | 1848 | 1.2520 | AUU | 1811 | 1.2902 | ||
| Lys | AAA | 2274 | 1.3256 | Lys | AAA | 2200 | 1.3652 |
| AAG | 1157 | 0.6744 | AAG | 1023 | 0.6348 | ||
| Leu | CUA | 737 | 0.8600 | Leu | CUA | 802 | 0.9086 |
| CUC | 627 | 0.7316 | CUC | 687 | 0.7783 | ||
| CUG | 520 | 0.6068 | CUG | 463 | 0.5245 | ||
| CUU | 1051 | 1.2264 | CUU | 1099 | 1.2451 | ||
| UUA | 1103 | 1.2870 | UUA | 1143 | 1.2949 | ||
| UUG | 1104 | 1.2882 | UUG | 1102 | 1.2485 | ||
| Met | AUG | 893 | 1.0000 | Met | AUG | 807 | 1.0000 |
| Asn | AAC | 801 | 0.6185 | Asn | AAC | 824 | 0.6317 |
| AAU | 1789 | 1.3815 | AAU | 1785 | 1.3683 | ||
| Pro | CCA | 739 | 1.1934 | Pro | CCA | 777 | 1.1702 |
| CCC | 654 | 1.0561 | CCC | 696 | 1.0482 | ||
| CCG | 432 | 0.6976 | CCG | 487 | 0.7334 | ||
| CCU | 652 | 1.0529 | CCU | 696 | 1.0482 | ||
| Gln | CAA | 1163 | 1.4046 | Gln | CAA | 1048 | 1.4067 |
| CAG | 493 | 0.5954 | CAG | 442 | 0.5933 | ||
| Arg | AGA | 1175 | 1.8977 | Arg | AGA | 1114 | 1.8723 |
| AGG | 676 | 1.0918 | AGG | 606 | 1.0185 | ||
| CGA | 698 | 1.1273 | CGA | 655 | 1.1008 | ||
| CGC | 298 | 0.4813 | CGC | 318 | 0.5345 | ||
| CGG | 466 | 0.7526 | CGG | 463 | 0.7782 | ||
| CGU | 402 | 0.6493 | CGU | 414 | 0.6958 | ||
| Ser | AGC | 532 | 0.6829 | Ser | AGC | 520 | 0.6266 |
| AGU | 695 | 0.8922 | AGU | 657 | 0.7917 | ||
| UCA | 880 | 1.1297 | UCA | 1022 | 1.2316 | ||
| UCC | 810 | 1.0398 | UCC | 931 | 1.1219 | ||
| UCG | 694 | 0.8909 | UCG | 689 | 0.8303 | ||
| UCU | 1063 | 1.3646 | UCU | 1160 | 1.3979 | ||
| Thr | ACA | 704 | 1.1887 | Thr | ACA | 657 | 1.1521 |
| ACC | 554 | 0.9354 | ACC | 600 | 1.0522 | ||
| ACG | 422 | 0.7125 | ACG | 380 | 0.6664 | ||
| ACU | 689 | 1.1634 | ACU | 644 | 1.1293 | ||
| Val | GUA | 716 | 1.2079 | Val | GUA | 705 | 1.1634 |
| GUC | 467 | 0.7879 | GUC | 484 | 0.7987 | ||
| GUG | 409 | 0.6900 | GUG | 441 | 0.7277 | ||
| GUU | 779 | 1.3142 | GUU | 794 | 1.3102 | ||
| Trp | UGG | 757 | 1.0000 | Trp | UGG | 754 | 1.0000 |
| Tyr | UAC | 702 | 0.6689 | Tyr | UAC | 711 | 0.6749 |
| UAU | 1397 | 1.3311 | UAU | 1396 | 1.3251 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Zhu, Y.; Zheng, S.; Wang, J.; Cheng, H.; Chen, H.; Zhu, W. Comparative Genomic and Phylogenetic Analysis of Chloroplasts in Citrus paradisi Mac.cv. Cocktail. Genes 2025, 16, 544. https://doi.org/10.3390/genes16050544
Wu Q, Zhu Y, Zheng S, Wang J, Cheng H, Chen H, Zhu W. Comparative Genomic and Phylogenetic Analysis of Chloroplasts in Citrus paradisi Mac.cv. Cocktail. Genes. 2025; 16(5):544. https://doi.org/10.3390/genes16050544
Chicago/Turabian StyleWu, Qun, Yun Zhu, Shipei Zheng, Jiajun Wang, Huilin Cheng, Haimin Chen, and Weidong Zhu. 2025. "Comparative Genomic and Phylogenetic Analysis of Chloroplasts in Citrus paradisi Mac.cv. Cocktail" Genes 16, no. 5: 544. https://doi.org/10.3390/genes16050544
APA StyleWu, Q., Zhu, Y., Zheng, S., Wang, J., Cheng, H., Chen, H., & Zhu, W. (2025). Comparative Genomic and Phylogenetic Analysis of Chloroplasts in Citrus paradisi Mac.cv. Cocktail. Genes, 16(5), 544. https://doi.org/10.3390/genes16050544