
Neuronal Network Activation Induced by Forniceal Deep Brain Stimulation in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
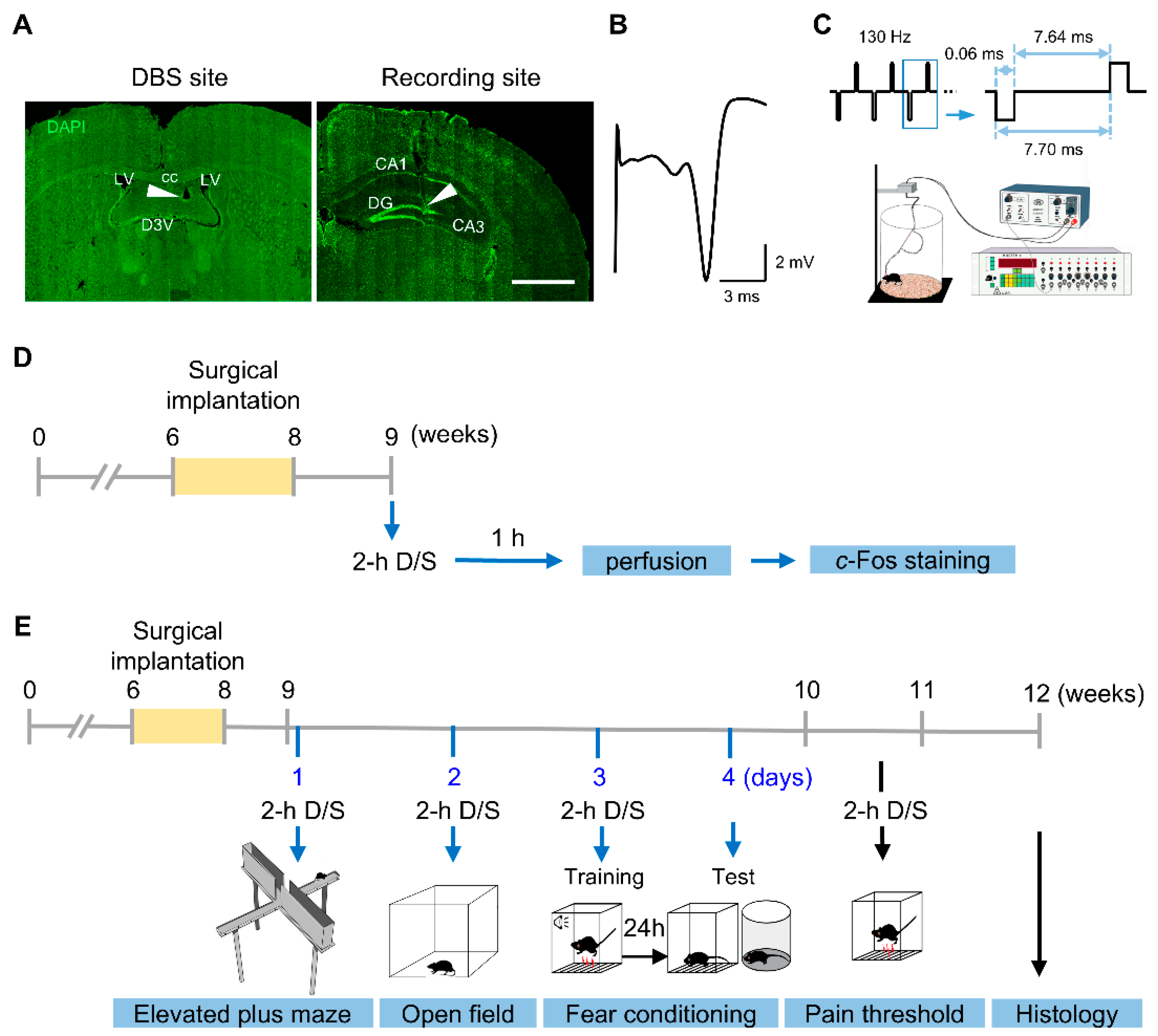
2. Materials and Methods
2.1. Animals
2.2. Surgery and DBS
2.3. Immunohistochemical Staining
2.4. Imaging and Quantification
2.5. Behavioral Assays
2.6. Histology
2.7. Statistical Analyses
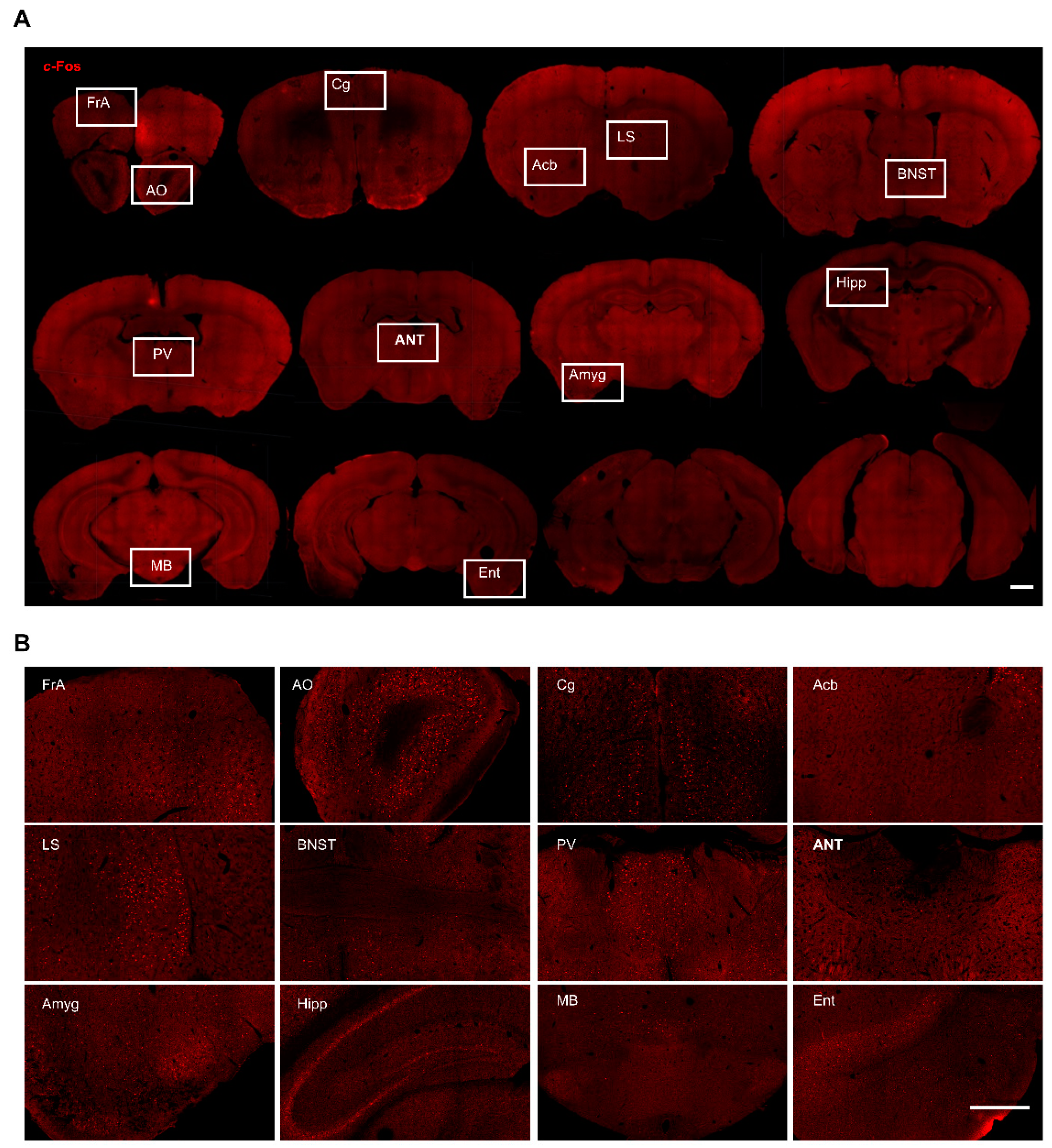
3. Results
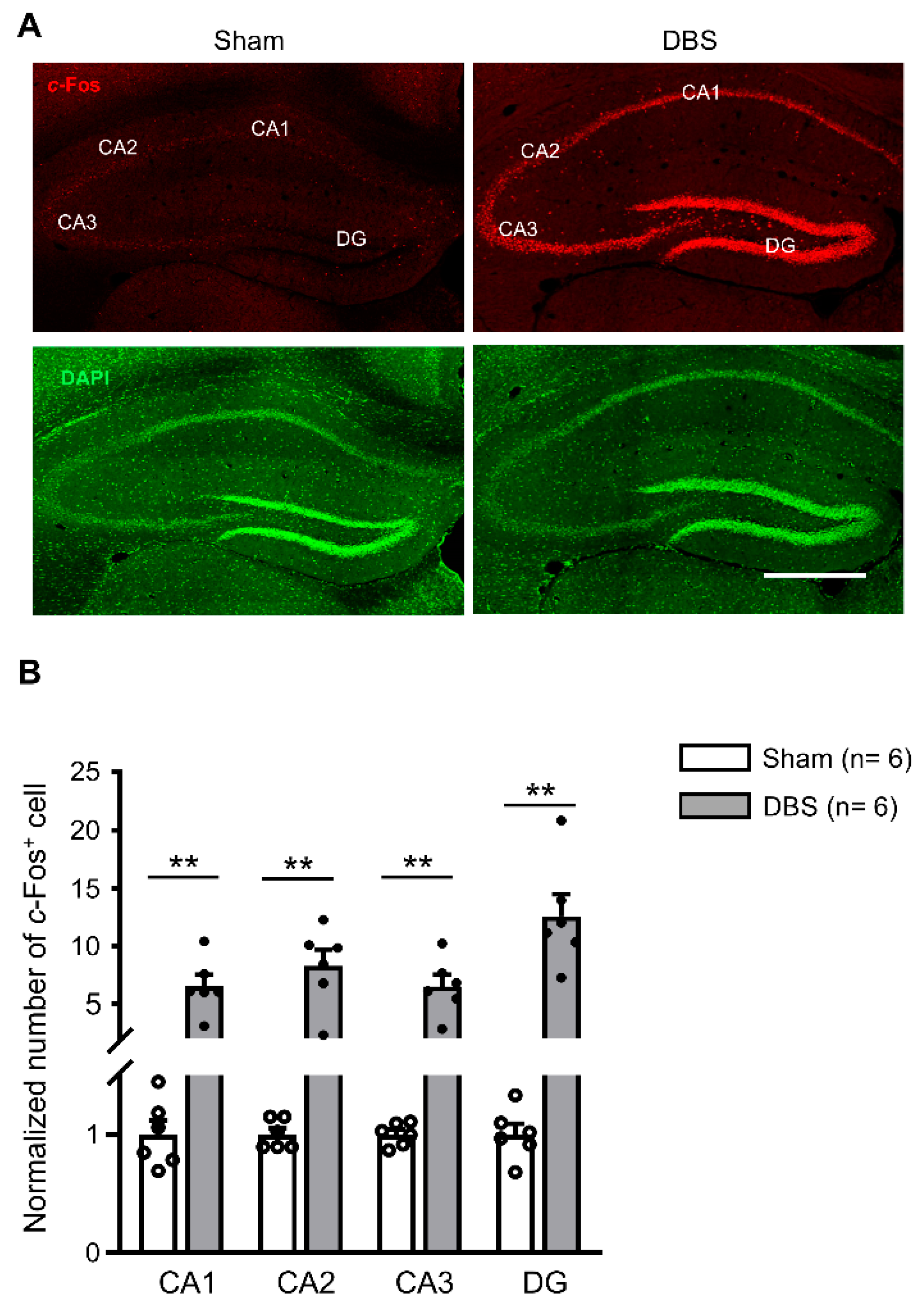
3.1. c-Fos Expression in the Hippocampus
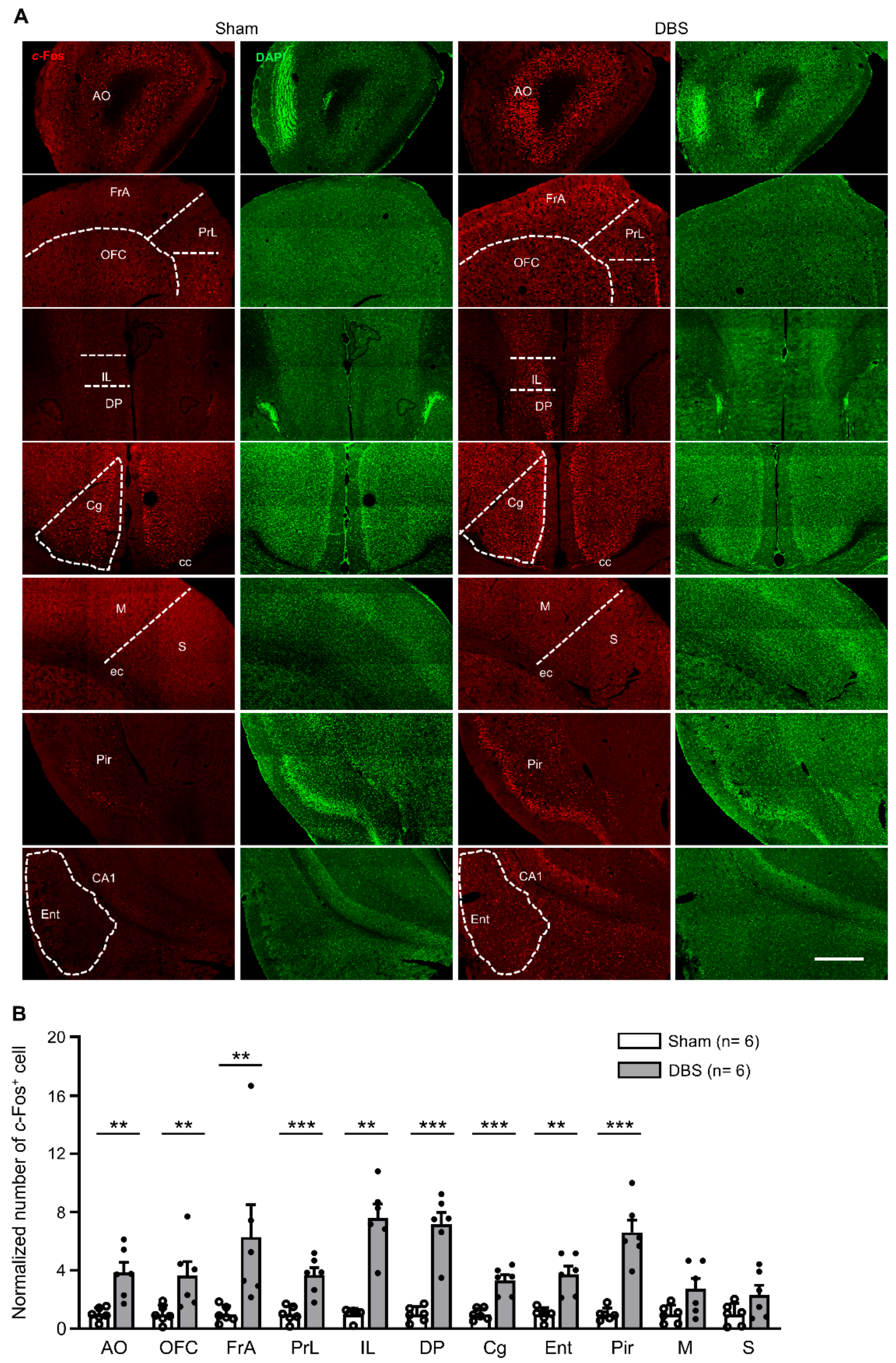
3.2. c-Fos Expression in the Cortices
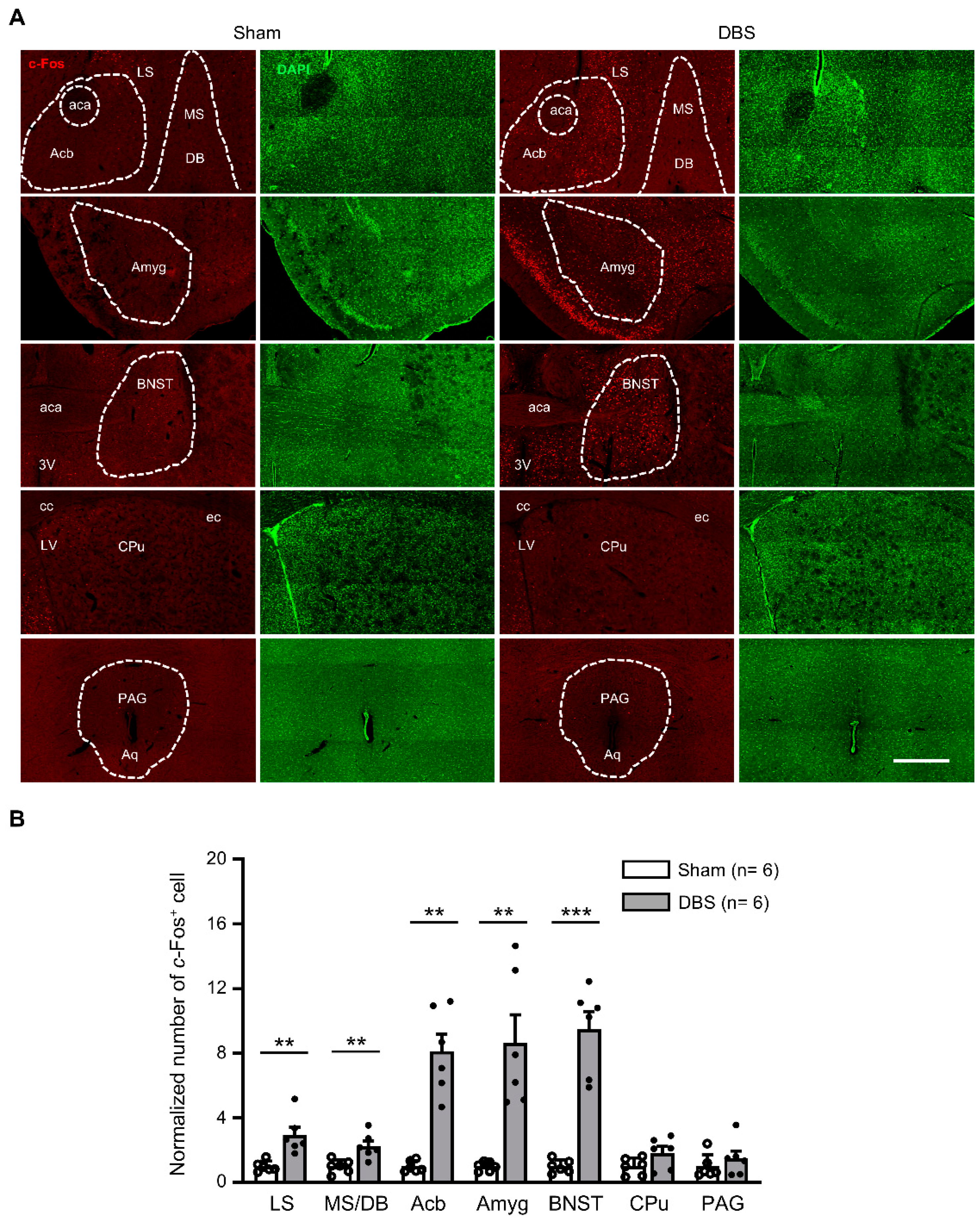
3.3. c-Fos Expression in the Subcortical Regions
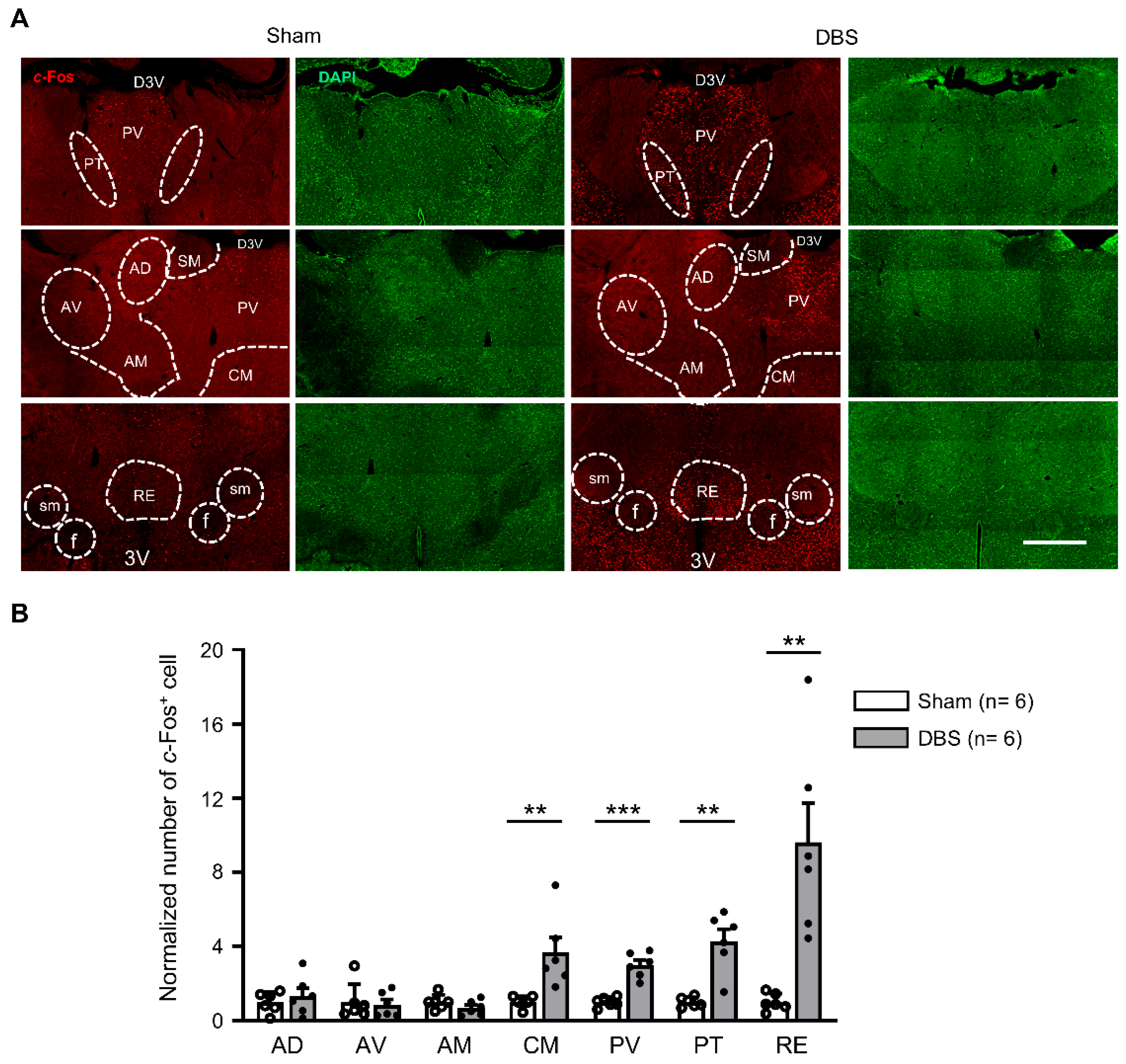
3.4. c-Fos Expression in the Thalamic Nuclei
3.5. c-Fos Expression in the Hypothalamic Nuclei
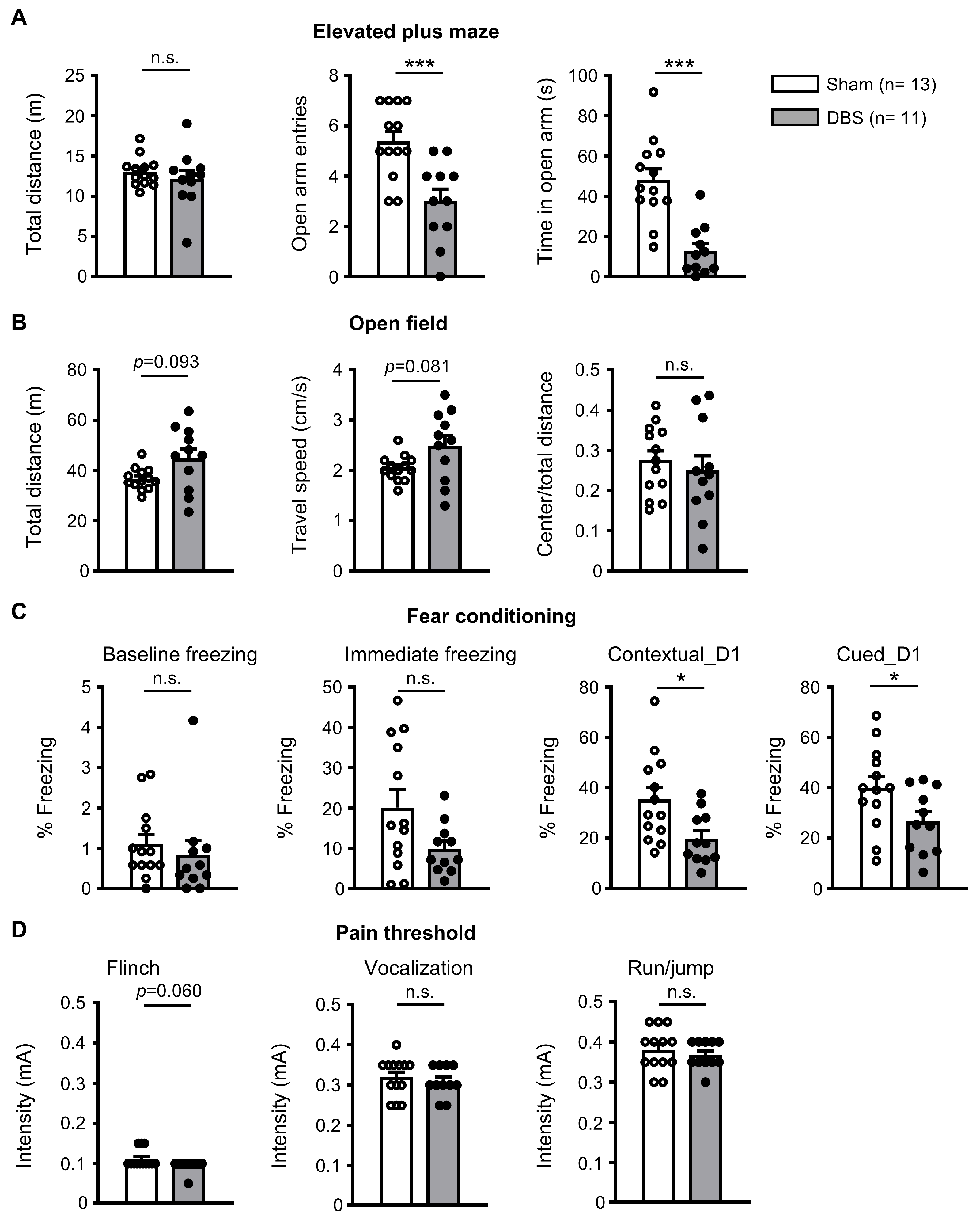
3.6. Behavioral Effect of Acute Forniceal DBS
4. Discussion
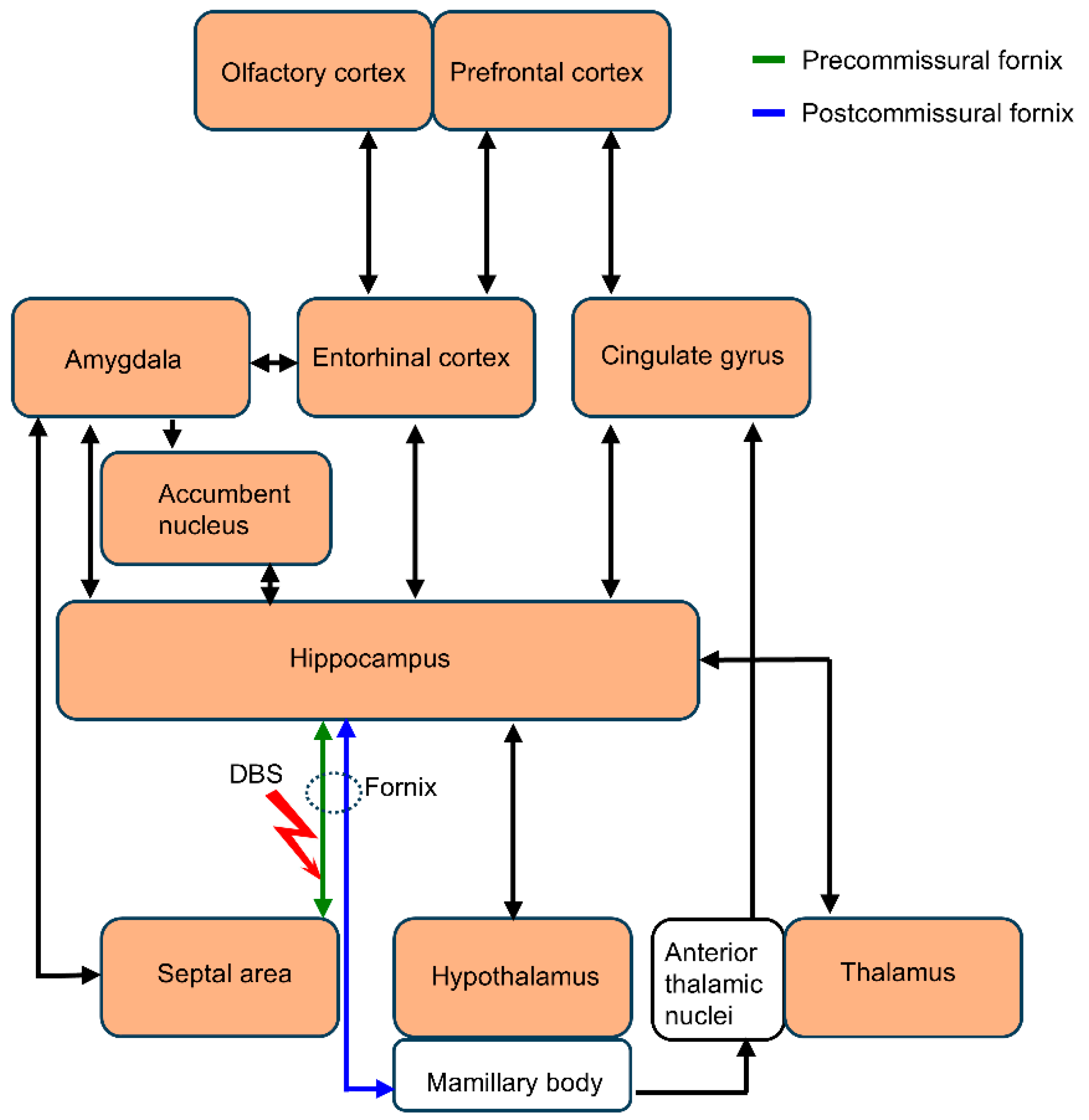
4.1. Hippocampal Circuit
4.2. Cortico-Hippocampal Circuit
4.3. Septo-Hippocampal Circuit
4.4. Hippocampal-Hypothalamic Circuit
4.5. Hippocampal–Thalamic Circuit
4.6. Hippocampal-Amygdala Circuit
4.7. Hippocampal–Accumbens Circuit
4.8. Hippocampal-Striatal Circuit
4.9. Pain Descending Modulation Circuit
4.10. The Effect of Acute Forniceal DBS on Emotion and Memory
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavallieri, F.; Mulroy, E.; Moro, E. The history of deep brain stimulation. Park. Relat. Disord. 2023, 121, 105980. [Google Scholar] [CrossRef]
- Senova, S.; Fomenko, A.; Gondard, E.; Lozano, A.M. Anatomy and function of the fornix in the context of its potential as a therapeutic target. J. Neurol. Neurosurg. Psychiatry 2020, 91, 547–559. [Google Scholar] [CrossRef]
- Hescham, S.; Lim, L.W.; Jahanshahi, A.; Steinbusch, H.W.; Prickaerts, J.; Blokland, A.; Temel, Y. Deep brain stimulation of the forniceal area enhances memory functions in experimental dementia: The role of stimulation parameters. Brain Stimul. 2013, 6, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Laxton, A.W.; Lozano, A.M. Deep brain stimulation for the treatment of Alzheimer disease and dementias. World Neurosurg. 2013, 80, S28.e1–S28.e8. [Google Scholar] [CrossRef]
- Laxton, A.W.; Tang-Wai, D.F.; McAndrews, M.P.; Zumsteg, D.; Wennberg, R.; Keren, R.; Wherrett, J.; Naglie, G.; Hamani, C.; Smith, G.S.; et al. A phase I trial of deep brain stimulation of memory circuits in Alzheimer’s disease. Ann. Neurol. 2010, 68, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Lozano, A.M.; Fosdick, L.; Chakravarty, M.M.; Leoutsakos, J.-M.; Munro, C.; Oh, E.; Drake, K.E.; Lyman, C.H.; Rosenberg, P.B.; Anderson, W.S.; et al. A Phase II Study of Fornix Deep Brain Stimulation in Mild Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 54, 777–787. [Google Scholar] [CrossRef]
- Hao, S.; Tang, B.; Wu, Z.; Ure, K.; Sun, Y.; Tao, H.; Gao, Y.; Patel, A.J.; Curry, D.J.; Samaco, R.C.; et al. Forniceal deep brain stimulation rescues hippocampal memory in Rett syndrome mice. Nature 2015, 526, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Wang, Q.; Tang, B.; Wu, Z.; Yang, T.; Tang, J. CDKL5 deficiency augments inhibitory input into the dentate gyrus that can be reversed by deep brain stimulation. J. Neurosci. 2021, 41, 9031–9046. [Google Scholar] [CrossRef]
- Shirvalkar, P.R.; Rapp, P.R.; Shapiro, M.L. Bidirectional changes to hippocampal theta–gamma comodulation predict memory for recent spatial episodes. Proc. Natl. Acad. Sci. USA 2010, 107, 7054–7059. [Google Scholar] [CrossRef]
- Pohodich, A.E.; Yalamanchili, H.; Raman, A.T.; Wan, Y.-W.; Gundry, M.; Hao, S.; Jin, H.; Tang, J.; Liu, Z.; Zoghbi, H.Y. Forniceal deep brain stimulation induces gene expression and splicing changes that promote neurogenesis and plasticity. eLife 2018, 7, e34031. [Google Scholar] [CrossRef]
- Lu, H.; Ash, R.T.; He, L.; Kee, S.E.; Wang, W.; Yu, D.; Hao, S.; Meng, X.; Ure, K.; Ito-Ishida, A.; et al. Loss and Gain of MeCP2 Cause Similar Hippocampal Circuit Dysfunction that Is Rescued by Deep Brain Stimulation in a Rett Syndrome Mouse Model. Neuron 2016, 91, 739–747. [Google Scholar] [CrossRef]
- Lowet, E.; Kondabolu, K.; Zhou, S.; Mount, R.A.; Wang, Y.; Ravasio, C.R.; Han, X. Deep brain stimulation creates informational lesion through membrane depolarization in mouse hippocampus. Nat. Commun. 2022, 13, 7709. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.; Ammari, R.; Bioulac, B.; Garcia, L. Latest view on the mechanism of action of deep brain stimulation. Mov. Disord. 2008, 23, 2111–2121. [Google Scholar] [CrossRef]
- VanElzakker, M.; Fevurly, R.D.; Breindel, T.; Spencer, R.L. Environmental novelty is associated with a selective increase in Fos expression in the output elements of the hippocampal formation and the perirhinal cortex. Learn. Mem. 2008, 15, 899–908. [Google Scholar] [CrossRef]
- Dragunow, M.; Faull, R. The use of c-fos as a metabolic marker in neuronal pathway tracing. J. Neurosci. Methods 1989, 29, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tang, B.; Tang, J. Protocol for deep brain stimulation in the fimbria-fornix of freely moving mice. STAR Protoc. 2022, 3, 101054. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tang, B.; Hao, S.; Wu, Z.; Yang, T.; Tang, J. Forniceal deep brain stimulation in a mouse model of Rett syndrome increases neurogenesis and hippocampal memory beyond the treatment period. Brain Stimul. 2023, 16, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.B.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic Press: London, UK, 1997. [Google Scholar]
- Lee, I.; Jerman, T.S.; Kesner, R.P. Disruption of delayed memory for a sequence of spatial locations following CA1- or CA3-lesions of the dorsal hippocampus. Neurobiol. Learn. Mem. 2005, 84, 138–147. [Google Scholar] [CrossRef]
- Farovik, A.; Dupont, L.M.; Eichenbaum, H. Distinct roles for dorsal CA3 and CA1 in memory for sequential nonspatial events. Learn. Mem. 2010, 17, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Maren, S. Differential roles for hippocampal areas CA1 and CA3 in the contextual encoding and retrieval of extinguished fear. Learn. Mem. 2008, 15, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Tsien, J.Z.; Huerta, P.T.; Tonegawa, S. The essential role of hippocampal CA1 NMDA receptor–Dependent synaptic plasticity in spatial memory. Cell 1996, 87, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Leutgeb, J.K.; Leutgeb, S.; Moser, M.-B.; Moser, E.I. Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science 2007, 315, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Nakashiba, T.; Young, J.Z.; McHugh, T.J.; Buhl, D.L.; Tonegawa, S. Transgenic inhibition of synaptic transmission reveals role of CA3 output in hippocampal learning. Science 2008, 319, 1260–1264. [Google Scholar] [CrossRef]
- Nakazawa, K.; Quirk, M.C.; Chitwood, R.A.; Watanabe, M.; Yeckel, M.F.; Sun, L.D.; Kato, A.; Carr, C.A.; Johnston, D.; Wilson, M.A.; et al. Requirement for hippocampal CA3 NMDA receptors in associative memory recall. Science 2002, 297, 211–218. [Google Scholar] [CrossRef] [PubMed]
- McHugh, T.J.; Jones, M.W.; Quinn, J.J.; Balthasar, N.; Coppari, R.; Elmquist, J.K.; Lowell, B.B.; Fanselow, M.S.; Wilson, M.A.; Tonegawa, S. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science 2007, 317, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Lehr, A.B.; Kumar, A.; Tetzlaff, C.; Hafting, T.; Fyhn, M.; Stöber, T.M. CA2 beyond social memory: Evidence for a fundamental role in hippocampal information processing. Neurosci. Biobehav. Rev. 2021, 126, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Sweet, J.A.; Eakin, K.C.; Munyon, C.N.; Miller, J.P. Improved learning and memory with theta-burst stimulation of the fornix in rat model of traumatic brain injury. Hippocampus 2014, 24, 1592–1600. [Google Scholar] [CrossRef] [PubMed]
- Stypulkowski, P.H.; Stanslaski, S.R.; Giftakis, J.E. Modulation of hippocampal activity with fornix Deep Brain Stimulation. Brain Stimul. 2017, 10, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Witter, M.P.; Doan, T.P.; Jacobsen, B.; Nilssen, E.S.; Ohara, S. Architecture of the Entorhinal Cortex A Review of Entorhinal Anatomy in Rodents with Some Comparative Notes. Front. Syst. Neurosci. 2017, 11, 46. [Google Scholar] [CrossRef]
- Morrissey, M.D.; Takehara-Nishiuchi, K. Diversity of mnemonic function within the entorhinal cortex: A meta-analysis of rodent behavioral studies. Neurobiol. Learn. Mem. 2014, 115, 95–107. [Google Scholar] [CrossRef]
- Kerr, K.M.; Agster, K.L.; Furtak, S.C.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region: The lateral and medial entorhinal areas. Hippocampus 2007, 17, 697–708. [Google Scholar] [CrossRef]
- Agster, K.L.; Burwell, R.D. Cortical efferents of the perirhinal, postrhinal, and entorhinal cortices of the rat. Hippocampus 2009, 19, 1159–1186. [Google Scholar] [CrossRef] [PubMed]
- Le Merre, P.; Ährlund-Richter, S.; Carlén, M. The mouse prefrontal cortex: Unity in diversity. Neuron 2021, 109, 1925–1944. [Google Scholar] [CrossRef] [PubMed]
- Mathiasen, M.L.; Aggleton, J.P.; Witter, M.P. Projections of the insular cortex to orbitofrontal and medial prefrontal cortex: A tracing study in the rat. Front. Neuroanat. 2023, 17, 1131167. [Google Scholar] [CrossRef] [PubMed]
- Ragozzino, M.E.; Detrick, S.; Kesner, R.P. Involvement of the prelimbic-infralimbic areas of the rodent prefrontal cortex in behavioral flexibility for place and response learning. J. Neurosci. 1999, 19, 4585–4594. [Google Scholar] [CrossRef] [PubMed]
- Takada, M. Neuroanatomy of Frontal Association Cortex. Brain Nerve 2016, 68, 1253–1261. [Google Scholar]
- McDonald, A.J. Organization of amygdaloid projections to the mediodorsal thalamus and prefrontal cortex: A fluorescence retrograde transport study in the rat. J. Comp. Neurol. 1987, 262, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Condé, F.; Maire-Lepoivre, E.; Audinat, E.; Crépel, F. Afferent connections of the medial frontal cortex of the rat. II. Cortical and subcortical afferents. J. Comp. Neurol. 1995, 352, 567–593. [Google Scholar] [CrossRef]
- Nakayama, D.; Baraki, Z.; Onoue, K.; Ikegaya, Y.; Matsuki, N.; Nomura, H. Frontal association cortex is engaged in stimulus integration during associative learning. Curr. Biol. 2015, 25, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Vanderwolf, C. Hippocampal activity, olfaction, and sniffing: An olfactory input to the dentate gyrus. Brain Res. 1992, 593, 197–208. [Google Scholar] [CrossRef]
- van Groen, T.; Wyss, J.M. Extrinsic projections from area CA1 of the rat hippocampus: Olfactory, cortical, subcortical, and bilateral hippocampal formation projections. J. Comp. Neurol. 1990, 302, 515–528. [Google Scholar] [CrossRef]
- Li, R.; Zhang, C.; Rao, Y.; Yuan, T.-F. Deep brain stimulation of fornix for memory improvement in Alzheimer’s disease: A critical review. Ageing Res. Rev. 2022, 79, 101668. [Google Scholar] [CrossRef] [PubMed]
- Khakpai, F.; Nasehi, M.; Haeri-Rohani, A.; Eidi, A.; Zarrindast, M.R. Septo-hippocampo-septal loop and memory formation. Basic Clin. Neurosci. 2013, 4, 5–23. [Google Scholar]
- Khakpai, F.; Zarrindast, M.R.; Nasehi, M.; Haeri-Rohani, A.; Eidi, A. The role of glutamatergic pathway between septum and hippocampus in the memory formation. EXCLI J. 2013, 12, 41–51. [Google Scholar]
- Risold, P.; Swanson, L. Connections of the rat lateral septal complex. Brain Res. Rev. 1997, 24, 115–195. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, T.P.; Chambers, R.; Russell, D.S. Regulation of affect by the lateral septum: Implications for neuropsychiatry. Brain Res. Rev. 2004, 46, 71–117. [Google Scholar] [CrossRef] [PubMed]
- Rizzi-Wise, C.A.; Wang, D.V. Putting Together Pieces of the Lateral Septum: Multifaceted Functions and Its Neural Pathways. eNeuro 2021, 8, ENEURO.0315-21.2021. [Google Scholar] [CrossRef] [PubMed]
- Thinschmidt, J.S.; Frazier, C.J.; King, M.A.; Meyer, E.M.; Papke, R.L. Medial septal/diagonal band cells express multiple functional nicotinic receptor subtypes that are correlated with firing frequency. Neurosci. Lett. 2005, 389, 163–168. [Google Scholar] [CrossRef]
- Alonso, A.; Köhler, C. Evidence for separate projections of hippocampal pyramidal and non-pyramidal neurons to different parts of the septum in the rat brain. Neurosci. Lett. 1982, 31, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Takács, V.T.; Freund, T.F.; Gulyás, A.I. Types and synaptic connections of hippocampal inhibitory neurons reciprocally connected with the medial septum. Eur. J. Neurosci. 2008, 28, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Toth, K.; Borhegyi, Z.; Freund, T. Postsynaptic targets of GABAergic hippocampal neurons in the medial septum-diagonal band of broca complex. J. Neurosci. 1993, 13, 3712–3724. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Remy, S. Septo-hippocampal interaction. Cell Tissue Res. 2018, 373, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Chartock, H.E. Interactions of septal evoked responses to stimulation of the fornix and medial forebrain bundle. Brain Res. 1979, 172, 439–451. [Google Scholar] [CrossRef] [PubMed]
- McLennan, H.; Miller, J.J. The hippocampal control of neuronal discharges in the septum of the rat. J. Physiol. 1974, 237, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, D.A.; Reinoso-Suarez, F. The topographic organization of hypothalamic and brain stem projections to the hippocampus. Brain Res. Bull. 1978, 3, 373–389. [Google Scholar] [CrossRef]
- Stanfield, B.B.; Cowan, W.M. An EM autoradiographic study of the hypothalamo-hippocampal projection. Brain Res. 1984, 309, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Petrovich, G.D. Lateral Hypothalamus as a Motivation-Cognition Interface in the Control of Feeding Behavior. Front. Syst. Neurosci. 2018, 12, 14. [Google Scholar] [CrossRef]
- Bang, J.Y.; Sunstrum, J.K.; Garand, D.; Parfitt, G.M.; Woodin, M.; Inoue, W.; Kim, J. Hippocampal-hypothalamic circuit controls context-dependent innate defensive responses. eLife 2022, 11, e74736. [Google Scholar] [CrossRef]
- Yan, J.-J.; Ding, X.-J.; He, T.; Chen, A.-X.; Zhang, W.; Yu, Z.-X.; Cheng, X.-Y.; Wei, C.-Y.; Hu, Q.-D.; Liu, X.-Y.; et al. A circuit from the ventral subiculum to anterior hypothalamic nucleus GABAergic neurons essential for anxiety-like behavioral avoidance. Nat. Commun. 2022, 13, 7464. [Google Scholar] [CrossRef]
- Bang, J.Y.; Zhao, J.; Rahman, M.; St-Cyr, S.; McGowan, P.O.; Kim, J.C. Hippocampus-Anterior Hypothalamic Circuit Modulates Stress-Induced Endocrine and Behavioral Response. Front. Neural Circuits 2022, 16, 894722. [Google Scholar] [CrossRef]
- Paolini, A.; McKenzie, J. Lesions in the magnocellular preoptic nucleus decrease olfactory investigation in rats. Behav. Brain Res. 1996, 81, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.J.; Fleming, A.S.; Lee, A.; Magnusson, J.E. The effects of olfactory and somatosensory desensitization on Fos-like immunoreactivity in the brains of pup-exposed postpartum rats. Behav. Neurosci. 1996, 110, 134–153. [Google Scholar] [CrossRef]
- Fewell, G.D.; Meredith, M. Experience facilitates vomeronasal and olfactory influence on Fos expression in medial preoptic area during pheromone exposure or mating in male hamsters. Brain Res. 2002, 941, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.-X.; McNaughton, N. The supramammillary area: Its organization, functions and relationship to the hippocampus. Prog. Neurobiol. 2004, 74, 127–166. [Google Scholar] [CrossRef] [PubMed]
- Vann, S.; Brown, M.; Aggleton, J. Fos expression in the rostral thalamic nuclei and associated cortical regions in response to different spatial memory tests. Neuroscience 2000, 101, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Vann, S.D.; Brown, M.W.; Erichsen, J.T.; Aggleton, J.P. Using fos imaging in the rat to reveal the anatomical extent of the disruptive effects of fornix lesions. J. Neurosci. 2000, 20, 8144–8152. [Google Scholar] [CrossRef]
- Chen, S.; He, L.; Huang, A.J.Y.; Boehringer, R.; Robert, V.; Wintzer, M.E.; Polygalov, D.; Weitemier, A.Z.; Tao, Y.; Gu, M.; et al. A hypothalamic novelty signal modulates hippocampal memory. Nature 2020, 586, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.-X.; McNaughton, N. The medial supramammillary nucleus, spatial learning and the frequency of hippocampal theta activity. Brain Res. 1997, 764, 101–108. [Google Scholar] [CrossRef]
- Li, Y.; Bao, H.; Luo, Y.; Yoan, C.; Sullivan, H.A.; Quintanilla, L.; Wickersham, I.; Lazarus, M.; Shih, Y.-Y.I.; Song, J. Supramammillary nucleus synchronizes with dentate gyrus to regulate spatial memory retrieval through glutamate release. eLife 2020, 9, e53129. [Google Scholar] [CrossRef]
- Li, Y.-D.; Luo, Y.-J.; Chen, Z.-K.; Quintanilla, L.; Cherasse, Y.; Zhang, L.; Lazarus, M.; Huang, Z.-L.; Song, J. Hypothalamic modulation of adult hippocampal neurogenesis in mice confers activity-dependent regulation of memory and anxiety-like behavior. Nat. Neurosci. 2022, 25, 630–645. [Google Scholar] [CrossRef]
- Jang, S.H.; Yeo, S.S. The differences of the precommissural and postcommissural fornix in the hippocampal location: A diffusion tensor tractography study. Neuroradiology 2017, 59, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.C.; Reddy, V.; Mayes, D.A. Neuroanatomy, Mammillary Bodies. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Henderson, J.; Greene, E. Behavioral effects of lesions of precommissural and postcommissural fornix. Brain Res. Bull. 1977, 2, 123–129. [Google Scholar] [CrossRef]
- Thomas, G.J. Delayed alternation in rats after pre- or postcommissural fornicotomy. J. Comp. Physiol. Psychol. 1978, 92, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Dillingham, C.M.; Frizzati, A.; Nelson, A.J.; Vann, S.D. How do mammillary body inputs contribute to anterior thalamic function? Neurosci. Biobehav. Rev. 2015, 54, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Shweman, S.M. Thalamus. Scholarpedia 2006, 1, 1583. [Google Scholar]
- Nelson, A.J.D. The anterior thalamic nuclei and cognition: A role beyond space? Neurosci. Biobehav. Rev. 2021, 126, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.; Moritz, C.; Quigley, M.; Cordes, D.; Haughton, V.; Meyerand, E. Functional Connectivity in the thalamus and hippocampus studied with functional MR imaging. AJNR Am. J. Neuroradiol. 2000, 21, 1397–1401. [Google Scholar] [PubMed]
- Vertes, R.P.; Linley, S.B.; Hoover, W.B. Limbic circuitry of the midline thalamus. Neurosci. Biobehav. Rev. 2015, 54, 89–107. [Google Scholar] [CrossRef]
- Scheel, N.; Wulff, P.; Malsen, J.G.d.M. Afferent connections of the thalamic nucleus reuniens in the mouse. J. Comp. Neurol. 2020, 528, 1189–1202. [Google Scholar] [CrossRef]
- Vertes, R.; Hoover, W.; Rodriguez, J. Projections of the central medial nucleus of the thalamus in the rat: Node in cortical, striatal and limbic forebrain circuitry. Neuroscience 2012, 219, 120–136. [Google Scholar] [CrossRef] [PubMed]
- Saunders, R.C.; Rosene, D.L.; Van Hoesen, G.W. Comparison of the efferents of the amygdala and the hippocampal formation in the rhesus monkey: II. Reciprocal and non-reciprocal connections. J. Comp. Neurol. 1988, 271, 185–207. [Google Scholar] [CrossRef]
- Benes, F.M.; Berretta, S. Amygdalo-entorhinal inputs to the hippocampal formation in relation to schizophrenia. Ann. New York Acad. Sci. 2000, 911, 293–304. [Google Scholar] [CrossRef]
- McGaugh, J.L. Memory consolidation and the amygdala: A systems perspective. Trends Neurosci. 2002, 25, 456–461. [Google Scholar] [CrossRef]
- Phelps, E.A. Human emotion and memory: Interactions of the amygdala and hippocampal complex. Curr. Opin. Neurobiol. 2004, 14, 198–202. [Google Scholar] [CrossRef]
- Fastenrath, M.; Coynel, D.; Spalek, K.; Milnik, A.; Gschwind, L.; Roozendaal, B.; Papassotiropoulos, A.; de Quervain, D.J. Dynamic modulation of amygdala–hippocampal connectivity by emotional arousal. J. Neurosci. 2014, 34, 13935–13947. [Google Scholar] [CrossRef] [PubMed]
- Lebow, M.A.; Chen, A. Overshadowed by the amygdala: The bed nucleus of the stria terminalis emerges as key to psychiatric disorders. Mol. Psychiatry 2016, 21, 450–463. [Google Scholar] [CrossRef]
- Goode, T.D.; Maren, S. Role of the bed nucleus of the stria terminalis in aversive learning and memory. Learn. Mem. 2017, 24, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Goode, T.D.; Ressler, R.L.; Acca, G.M.; Miles, O.W.; Maren, S.; States, U. Bed nucleus of the stria terminalis regulates fear to unpredictable threat signals. eLife 2019, 8, e46525. [Google Scholar] [CrossRef] [PubMed]
- Lingg, R.T.; Johnson, S.B.; Emmons, E.B.; Anderson, R.M.; Romig-Martin, S.A.; Narayanan, N.S.; McGaugh, J.L.; LaLumiere, R.T.; Radley, J.J. Bed nuclei of the stria terminalis modulate memory consolidation via glucocorticoid-dependent and independent circuits. Proc. Natl. Acad. Sci. USA 2020, 117, 8104–8114. [Google Scholar] [CrossRef]
- van Huijstee, A.N.; Mansvelder, H.D. Glutamatergic synaptic plasticity in the mesocorticolimbic system in addiction. Front. Cell Neurosci. 2014, 8, 466. [Google Scholar] [CrossRef] [PubMed]
- LeGates, T.A.; Kvarta, M.D.; Tooley, J.R.; Francis, T.C.; Lobo, M.K.; Creed, M.C.; Thompson, S.M. Reward behaviour is regulated by the strength of hippocampus–nucleus accumbens synapses. Nature 2018, 564, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B. The nucleus accumbens: An interface between cognition, emotion, and action. Annu. Rev. Psychol. 2015, 66, 25–52. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.K.; Kim, J.P.; Settell, M.L.; Han, S.R.; Blaha, C.D.; Min, H.-K.; Lee, K.H. Fornix deep brain stimulation circuit effect is dependent on major excitatory transmission via the nucleus accumbens. NeuroImage 2016, 128, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Lee, S.-Y.; Cho, H.-U.; Oh, Y.; Kim, I.Y.; Lee, K.H.; Jang, D.P.; Min, H.-K. Fornix Stimulation Induces Metabolic Activity and Dopaminergic Response in the Nucleus Accumbens. Front. Neurosci. 2019, 13, 1109. [Google Scholar] [CrossRef] [PubMed]
- Grahn, J.A.; Parkinson, J.A.; Owen, A.M. The cognitive functions of the caudate nucleus. Prog. Neurobiol. 2008, 86, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Budai, D.; Harasawa, I.; Fields, H.L. Midbrain periaqueductal gray (PAG) inhibits nociceptive inputs to sacral dorsal horn nociceptive neurons through α2-adrenergic receptors. J. Neurophysiol. 1998, 80, 2244–2254. [Google Scholar] [CrossRef] [PubMed]
- Yeh, L.-F.; Ozawa, T.; Johansen, J.P. Functional organization of the midbrain periaqueductal gray for regulating aversive memory formation. Mol. Brain 2021, 14, 136. [Google Scholar] [CrossRef]
- Herry, C.; Johansen, J.P. Encoding of fear learning and memory in distributed neuronal circuits. Nat. Neurosci. 2014, 17, 1644–1654. [Google Scholar] [CrossRef]
- Ghasemi, M.; Navidhamidi, M.; Rezaei, F.; Azizikia, A.; Mehranfard, N. Anxiety and hippocampal neuronal activity: Relationship and potential mechanisms. Cogn. Affect. Behav. Neurosci. 2022, 22, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.I.; Ressler, K.J.; Binder, E.; Nemeroff, C.B. The neurobiology of anxiety disorders: Brain imaging, genetics, and psychoneuroendocrinology. Clin. Lab. Med. 2010, 30, 865–891. [Google Scholar] [CrossRef]
- Liberzon, I.; Duval, E.; Javanbakht, A. Neural circuits in anxiety and stress disorders: A focused review. Ther. Clin. Risk Manag. 2015, 11, 115–126. [Google Scholar] [CrossRef]
- Maloney, E.A.; Sattizahn, J.R.; Beilock, S.L. Anxiety and cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 403–411. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, B.; Wu, Z.; Wang, Q.; Tang, J. Neuronal Network Activation Induced by Forniceal Deep Brain Stimulation in Mice. Genes 2025, 16, 210. https://doi.org/10.3390/genes16020210
Tang B, Wu Z, Wang Q, Tang J. Neuronal Network Activation Induced by Forniceal Deep Brain Stimulation in Mice. Genes. 2025; 16(2):210. https://doi.org/10.3390/genes16020210
Chicago/Turabian StyleTang, Bin, Zhenyu Wu, Qi Wang, and Jianrong Tang. 2025. "Neuronal Network Activation Induced by Forniceal Deep Brain Stimulation in Mice" Genes 16, no. 2: 210. https://doi.org/10.3390/genes16020210
APA StyleTang, B., Wu, Z., Wang, Q., & Tang, J. (2025). Neuronal Network Activation Induced by Forniceal Deep Brain Stimulation in Mice. Genes, 16(2), 210. https://doi.org/10.3390/genes16020210