
Central Roles of ZmNAC128 and ZmNAC130 in Nutrient Uptake and Storage during Maize Grain Filling


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. RNA Sequencing (RNA-Seq)
2.3. Quantification of Lipid Content in Mature Kernels
2.4. Analysis of Element Content in Mature Kernels
2.5. Determination of Phytohormone Content in 16-DAP Endosperm
2.6. RNA Extraction and Quantitative RT-PCR
2.7. Dual-Luciferase Reporter (DLR) Assay
2.8. Phylogenetic Analysis
2.9. Statistical Analysis
3. Results
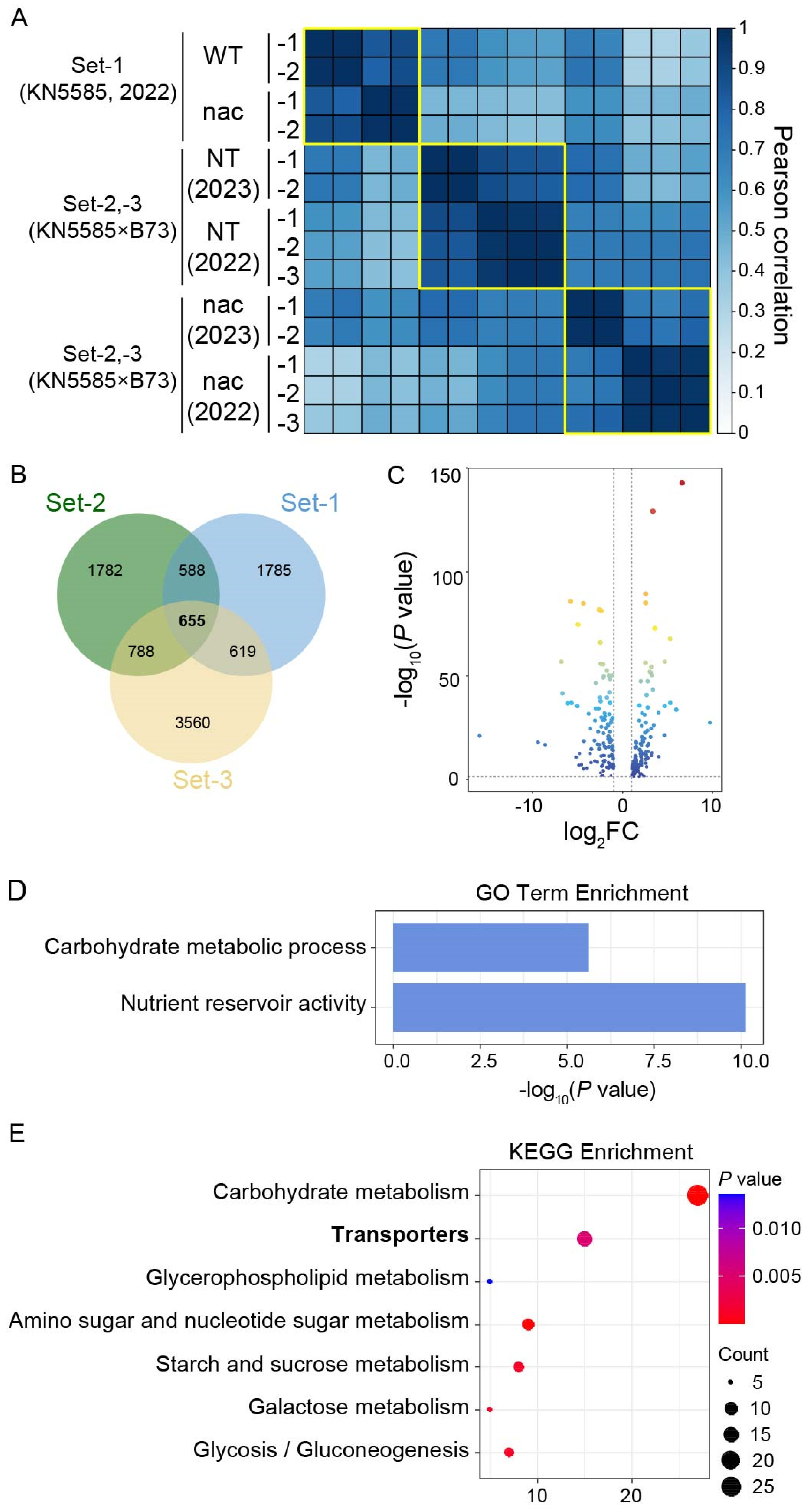
3.1. Generation and Analysis of Transcriptomes from Endosperms of zmnac128 zmnac130 Mutants across Different Genetic Backgrounds and Growing Years
3.2. Identifying the hdEGs across Different Genetic Backgrounds and Growing Years as Core Members in the Regulatory Network of ZmNAC128 and ZmNAC130
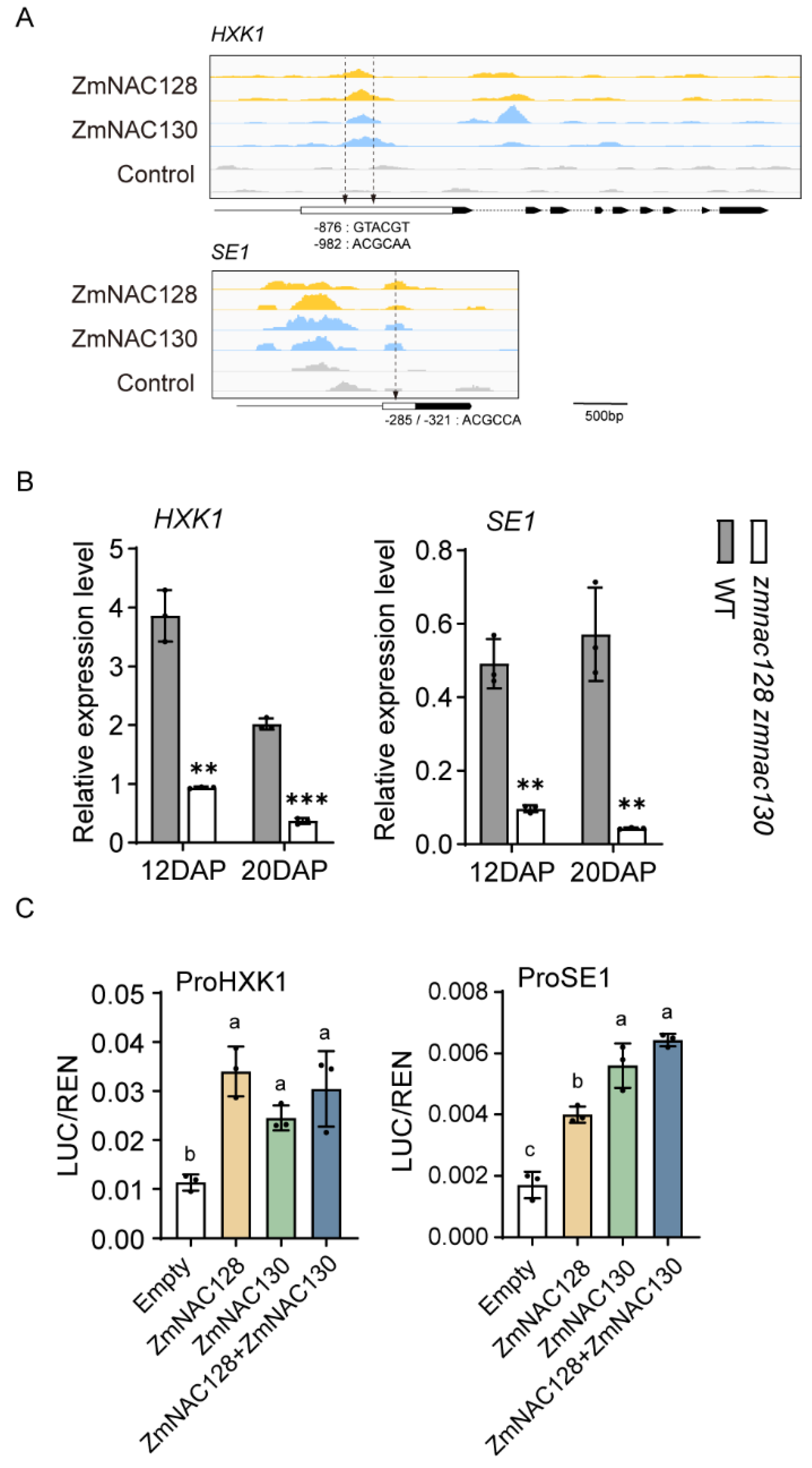
3.3. Identifying Starch Metabolism-Related Sugary Enhancer 1 (SE1) and Hexokinase 1 (HXK1) as Direct Targets of ZmNAC128 and ZmNAC130
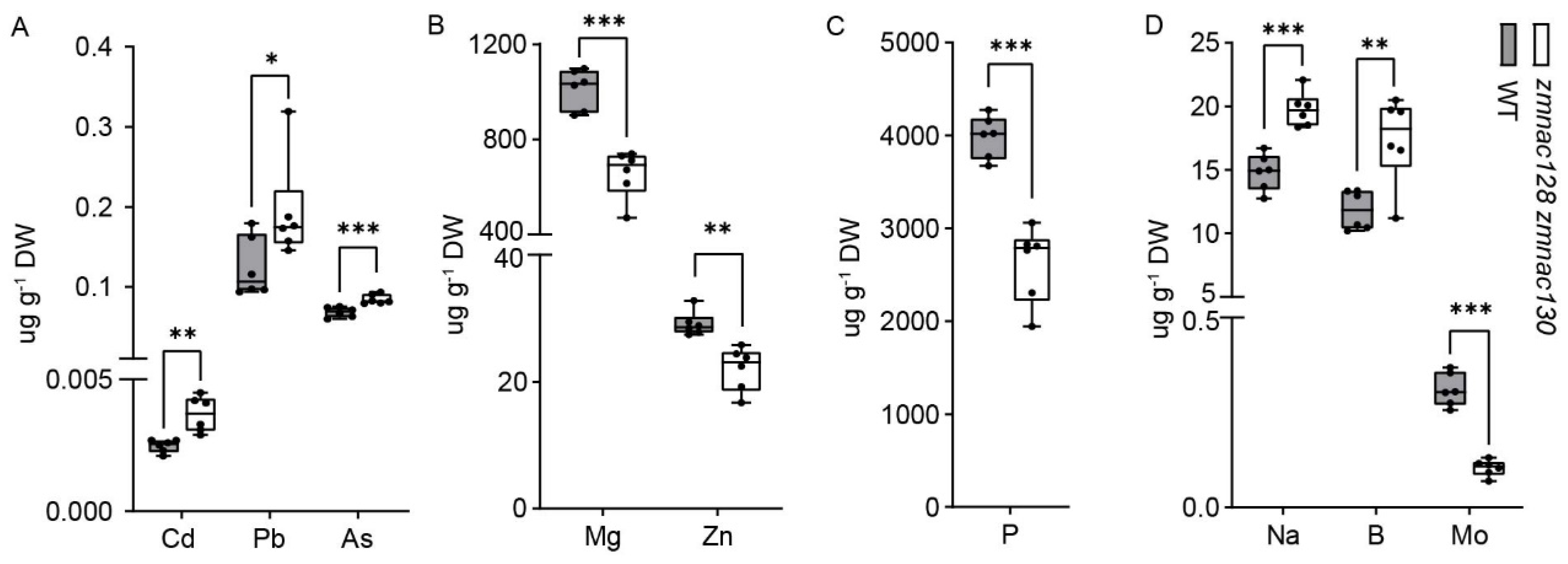
3.4. Regulatory Roles of ZmNAC128 and ZmNAC130 on Toxic HMM Accumulation and Biofortification in Maize Kernels
3.5. ZmNAC128 and ZmNAC130 Directly Regulate the Expression of Grain-Filling-Controlling P Transporter ZmPHO1;2
3.6. A Regulatory Mechanism of ZmNAC128 and ZmNAC130 in the Expression of the Endosperm-Specific Nitrate Transporter ZmNPF1.1
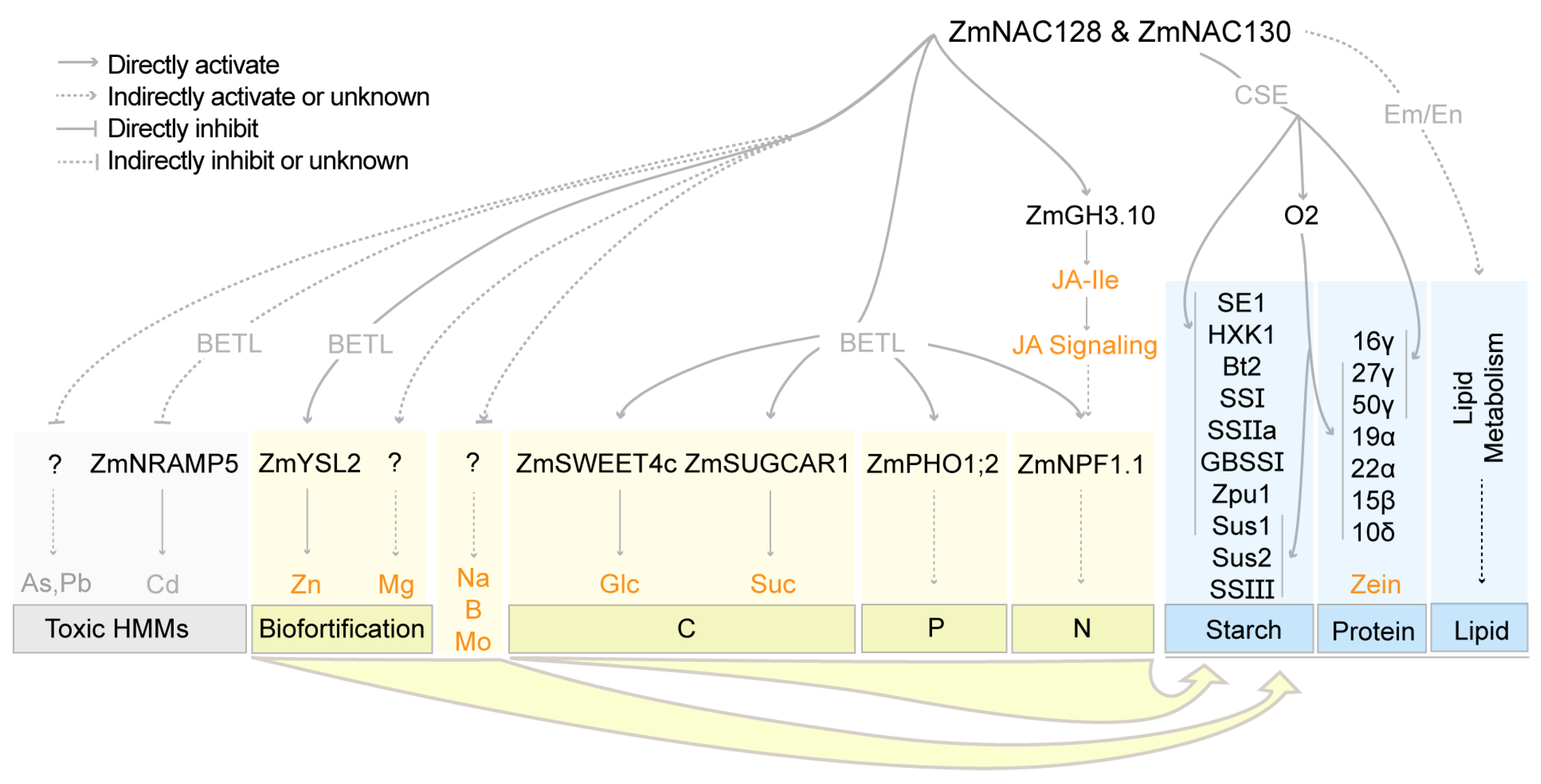
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sabelli, P.A.; Larkins, B.A. The development of endosperm in grasses. Plant Physiol. 2009, 149, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.E.; Ma, F. Determinants of Kernel Sink Strength; CABI: Wallingford, UK, 2017. [Google Scholar]
- Sosso, D.; Luo, D.P.; Li, Q.B.; Sasse, J.; Yang, J.L.; Gendrot, G.; Suzuki, M.; Koch, K.E.; McCarty, D.R.; Chourey, P.S.; et al. Seed filling in domesticated maize and rice depends on SWEET-mediated hexose transport. Nat. Genet. 2015, 47, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, J.; Yu, M.; Zhang, M.; Zhong, Y.; Wang, T.; Liu, P.; Song, W.; Zhao, H.; Fastner, A.; et al. The sugar transporter ZmSUGCAR1 of the Nitrate Transporter 1/Peptide Transporter family is critical for maize grain filling. Plant Cell 2022, 34, 4232–4254. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.F.; Leach, K.A.; Boyer, N.R.; Swyers, M.J.; Benitez-Alfonso, Y.; Skopelitis, T.; Luo, A.; Sylvester, A.; Jackson, D.; Braun, D.M. Sucrose Transporter ZmSut1 Expression and Localization Uncover New Insights into Sucrose Phloem Loading. Plant Physiol. 2016, 172, 1876–1898. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xiao, W.; Tian, L.; Guo, L.; Ma, G.; Ji, C.; Huang, Y.; Wang, H.; Wu, X.; Yang, T.; et al. Spatial transcriptomics uncover sucrose post-phloem transport during maize kernel development. Nat. Commun. 2023, 14, 7191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.H.; Taliercio, E.W.; Chourey, P.S. The Miniature1 seed locus of maize encodes a cell wall invertase required for normal development of endosperm and maternal cells in the pedicel. Plant Cell 1996, 8, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhang, L.; Gao, Q.F.; Wang, J.M.; Li, X.Y.; Wang, H.; Liu, Y.; Lin, H.; Liu, J.Y.; Wang, X.; et al. A plasma membrane transporter coordinates phosphate reallocation and grain filling in cereals. Nat. Genet. 2021, 53, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Nieves, A.L.; Salazar-Vidal, M.N.; Torres-Rodríguez, J.V.; Pérez-Vázquez, L.M.; Massange-Sánchez, J.A.; Gillmor, C.S.; Sawers, R.J.H. The pho1;2a′-m1.1 allele of Phosphate1 conditions misregulation of the phosphorus starvation response in maize (Zea mays ssp. mays L.). Plant Direct 2022, 6, e416. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.-F.; Chen, Y.-Y.; Ji, C.; Wang, Y.-L.; Huang, X.; Zhang, C.-Y.; Yang, J.; Song, T.; Wu, J.-C.; Guo, L.-X.; et al. A genome-wide association study identifies a transporter for zinc uploading to maize kernels. EMBO Rep. 2023, 24, e55542. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.; Boehlein, S. Starch Biosynthesis in Maize Endosperm; CABI: Wallingford, UK, 2017. [Google Scholar]
- Larkins, B.; YongRui, W.Y.W.; RenTao, S.R.S.; Messing, J. Maize Seed Storage Proteins; CABI: Wallingford, UK, 2017. [Google Scholar]
- Chen, J.; Zeng, B.; Zhang, M.; Xie, S.; Wang, G.; Hauck, A.; Lai, J. Dynamic transcriptome landscape of maize embryo and endosperm development. Plant Physiol. 2014, 166, 252–264. [Google Scholar] [CrossRef]
- Schmidt, R.J.; Burr, F.A.; Burr, B. Transposon Tagging and Molecular Analysis of the Maize Regulatory Locus Opaque-2. Science 1987, 238, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Burr, F.A.; Aukerman, M.J.; Burr, B. Maize Regulatory Gene Opaque-2 Encodes a Protein with a Leucine-Zipper Motif That Binds to Zein DNA. Proc. Natl. Acad. Sci. USA 1990, 87, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Li, C.B.; Qiao, Z.Y.; Qi, W.W.; Wang, Q.; Yuan, Y.; Yang, X.; Tang, Y.P.; Mei, B.; Lv, Y.D.; Zhao, H.; et al. Genome-Wide Characterization of cis-Acting DNA Targets Reveals the Transcriptional Regulatory Framework of Opaque2 in Maize. Plant Cell 2015, 27, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.P.; Li, G.S.; Ryu, C.H.; Ma, C.; Zhang, S.S.; Lloyd, A.; Hunter, B.G.; Larkins, B.A.; Drews, G.N.; Wang, X.F.; et al. Opaque-2 Regulates a Complex Gene Network Associated with Cell Differentiation and Storage Functions of Maize Endosperm. Plant Cell 2018, 30, 2425–2446. [Google Scholar] [CrossRef]
- VicenteCarbajosa, J.; Moose, S.P.; Parsons, R.L.; Schmidt, R.J. A maize zinc-finger protein binds the prolamin box in zein gene promoters and interacts with the basic leucine zipper transcriptional activator Opaque2. Proc. Natl. Acad. Sci. USA 1997, 94, 7685–7690. [Google Scholar] [CrossRef]
- Wu, Y.R.; Messing, J. Rapid Divergence of Prolamin Gene Promoters of Maize After Gene Amplification and Dispersal. Genetics 2012, 192, 507–519. [Google Scholar] [CrossRef]
- Pysh, L.D.; Aukerman, M.J.; Schmidt, R.J. OHP1—A Maize basic domain leucine zipper protein that interacts with Opaque2. Plant Cell 1993, 5, 227–236. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, J.; Wu, Y. Transcriptional Regulation of Zein Gene Expression in Maize through the Additive and Synergistic Action of Opaque2, Prolamine-Box Binding Factor, and O2 Heterodimerizing Proteins. Plant Cell 2015, 27, 1162–1172. [Google Scholar] [CrossRef]
- Gontarek, B.C.; Neelakandan, A.K.; Wu, H.; Becraft, P.W. NKD Transcription Factors Are Central Regulators of Maize Endosperm Development. Plant Cell 2016, 28, 2916–2936. [Google Scholar] [CrossRef]
- Qiao, Z.Y.; Qi, W.W.; Wang, Q.; Feng, Y.N.; Yang, Q.; Zhang, N.; Wang, S.S.; Tang, Y.P.; Song, R.T. ZmMADS47 Regulates Zein Gene Transcription through Interaction with Opaque2. PLoS Genet. 2016, 12, e1005991. [Google Scholar] [CrossRef]
- Li, C.; Yue, Y.; Chen, H.; Qi, W.; Song, R. The ZmbZIP22 Transcription Factor Regulates 27-kD γ-Zein Gene Transcription during Maize Endosperm Development. Plant Cell 2018, 30, 2402–2424. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, J.; Ji, C.; Wu, Y.; Messing, J. NAC-type transcription factors regulate accumulation of starch and protein in maize seeds. Proc. Natl. Acad. Sci. USA 2019, 116, 11223–11228. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Yu, H.; He, J.; Peng, D.; Zhu, P.; Pan, S.; Wu, X.; Wang, J.; Ji, C.; Chao, Z.; et al. The transcription factors ZmNAC128 and ZmNAC130 coordinate with Opaque2 to promote endosperm filling in maize. Plant Cell 2023, 35, 4066–4090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, X.; Yang, J.; Messing, J.; Wu, Y. Maize endosperm-specific transcription factors O2 and PBF network the regulation of protein and starch synthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 10842–10847. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Qi, W.; Lv, Y.; Yan, S.; Xu, L.; Yang, W.; Yuan, Y.; Chen, Y.; Zhao, H.; Song, R. OPAQUE11 Is a Central Hub of the Regulatory Network for Maize Endosperm Development and Nutrient Metabolism. Plant Cell 2018, 30, 375–396. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, L.; Ji, C.; Wang, H.; Wang, J.; Zheng, X.; Xiao, Q.; Wu, Y. The B3 domain-containing transcription factor ZmABI19 coordinates expression of key factors required for maize seed development and grain filling. Plant Cell 2021, 33, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, H.; Guo, L.; Wu, X.; Xiao, Q.; Wang, J.; Wang, Q.; Ma, G.; Wang, W.; Wu, Y. ABA-induced phosphorylation of basic leucine zipper 29, ABSCISIC ACID INSENSITIVE 19, and Opaque2 by SnRK2.2 enhances gene transactivation for endosperm filling in maize. Plant Cell 2022, 34, 1933–1956. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Galli, M.; Spears, C.J.; Zhan, J.; Liu, P.; Yadegari, R.; Dannenhoffer, J.M.; Gallavotti, A.; Becraft, P.W. NAKED ENDOSPERM1, NAKED ENDOSPERM2, and OPAQUE2 interact to regulate gene networks in maize endosperm development. Plant Cell 2023, 36, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, S.; Zhao, Q.; Lv, D.; Wang, B.; Xiao, K.; Zhu, J.; Li, S.; Yang, W.; Liu, X.; et al. ZmGRAS11, transactivated by Opaque2, positively regulates kernel size in maize. J. Integr. Plant Biol. 2021, 63, 2031–2037. [Google Scholar] [CrossRef]
- Chen, Q.; Guo, Y.; Zhang, J.; Zheng, N.; Wang, J.; Liu, Y.; Lu, J.; Zhen, S.; Du, X.; Li, L.; et al. RNA polymerase common subunit ZmRPABC5b is transcriptionally activated by Opaque2 and essential for endosperm development in maize. Nucleic Acids Res. 2023, 51, 7832–7850. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Lahner, B.; Gong, J.; Mahmoudian, M.; Smith, E.L.; Abid, K.B.; Rogers, E.E.; Guerinot, M.L.; Harper, J.F.; Ward, J.M.; McIntyre, L.; et al. Genomic scale profiling of nutrient and trace elements in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 1215–1221. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Finegan, C.; Boehlein, S.K.; Leach, K.A.; Madrid, G.; Hannah, L.C.; Koch, K.E.; Tracy, W.F.; Resende, M.F.R. Genetic Perturbation of the Starch Biosynthesis in Maize Endosperm Reveals Sugar-Responsive Gene Networks. Front. Plant Sci. 2022, 12, 800326. [Google Scholar] [CrossRef]
- Zhang, X.; Mogel, K.; Lor, V.S.; Hirsch, C.N.; De Vries, B.; Kaeppler, H.F.; Tracy, W.F.; Kaeppler, S.M. Maize sugary enhancer1 (se1) is a gene affecting endosperm starch metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 20776–20785. [Google Scholar] [CrossRef]
- Song, W.Y.; Yamaki, T.; Yamaji, N.; Ko, D.; Jung, K.H.; Fujii-Kashino, M.; An, G.; Martinoia, E.; Lee, Y.; Ma, J.F. A rice ABC transporter, OsABCC1, reduces arsenic accumulation in the grain. Proc. Natl. Acad. Sci. USA 2014, 111, 15699–15704. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Chen, M.; Zheng, G.; Peng, Y.; Shi, X.; Qin, P.; Xu, X.; Teng, S. Association Study Reveals Genetic Loci Responsible for Arsenic, Cadmium and Lead Accumulation in Rice Grain in Contaminated Farmlands. Front. Plant Sci. 2019, 10, 61. [Google Scholar] [CrossRef]
- Kar, D.; Pradhan, A.A.; Dutta, A.; Bhagavatula, L.; Datta, S. The Multidrug and Toxic Compound Extrusion (MATE) Family in Plants and Their Significance in Metal Transport. In Plant Metal and Metalloid Transporters; Kumar, K., Srivastava, S., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 151–177. [Google Scholar]
- Clemens, S.; Ma, J.F. Toxic Heavy Metal and Metalloid Accumulation in Crop Plants and Foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef]
- Jia, L.; Hu, D.; Wang, J.; Liang, Y.; Li, F.; Wang, Y.; Han, Y. Genome-Wide Identification and Functional Analysis of Nitrate Transporter Genes (NPF, NRT2 and NRT3) in Maize. Int. J. Mol. Sci. 2023, 24, 12941. [Google Scholar] [CrossRef]
- Delfin, J.C.; Kanno, Y.; Seo, M.; Kitaoka, N.; Matsuura, H.; Tohge, T.; Shimizu, T. AtGH3.10 is another jasmonic acid-amido synthetase in Arabidopsis thaliana. Plant J. 2022, 110, 1082–1096. [Google Scholar] [CrossRef]
- Feng, S.; Yue, R.; Tao, S.; Yang, Y.; Zhang, L.; Xu, M.; Wang, H.; Shen, C. Genome-wide identification, expression analysis of auxin-responsive GH3 family genes in maize (Zea mays L.) under abiotic stresses. J. Integr. Plant Biol. 2015, 57, 783–795. [Google Scholar] [CrossRef]
- Kemper, E.L.; Neto, G.C.; Papes, F.; Moraes, K.C.; Leite, A.; Arruda, P. The role of opaque2 in the control of lysine-degrading activities in developing maize endosperm. Plant Cell 1999, 11, 1981–1994. [Google Scholar] [CrossRef]
- Deng, Y.T.; Wang, J.C.; Zhang, Z.Y.; Wu, Y.R. Transactivation of Sus1 and Sus2 by Opaque2 is an essential supplement to sucrose synthase-mediated endosperm filling in maize. Plant Biotechnol. J. 2020, 18, 1897–1907. [Google Scholar] [CrossRef]
- Lappe, R.R.; Baier, J.W.; Boehlein, S.K.; Huffman, R.; Lin, Q.; Wattebled, F.; Settles, A.M.; Hannah, L.C.; Borisjuk, L.; Rolletschek, H.; et al. Functions of maize genes encoding pyruvate phosphate dikinase in developing endosperm. Proc. Natl. Acad. Sci. USA 2018, 115, E24–E33. [Google Scholar] [CrossRef]
- Ji, C.; Xu, L.; Li, Y.; Fu, Y.; Li, S.; Wang, Q.; Zeng, X.; Zhang, Z.; Zhang, Z.; Wang, W.; et al. The O2-ZmGRAS11 transcriptional regulatory network orchestrates the coordination of endosperm cell expansion and grain filling in maize. Mol. Plant 2022, 15, 468–487. [Google Scholar] [CrossRef]
- Yan, P.; Du, Q.; Chen, H.; Guo, Z.; Wang, Z.; Tang, J.; Li, W.X. Biofortification of iron content by regulating a NAC transcription factor in maize. Science 2023, 382, 1159–1165. [Google Scholar] [CrossRef]
- Gamuyao, R.; Chin, J.H.; Pariasca-Tanaka, J.; Pesaresi, P.; Catausan, S.; Dalid, C.; Slamet-Loedin, I.; Tecson-Mendoza, E.M.; Wissuwa, M.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–539. [Google Scholar] [CrossRef]
- Kettenburg, A.T.; Lopez, M.A.; Yogendra, K.; Prior, M.J.; Rose, T.; Bimson, S.; Heuer, S.; Roy, S.J.; Bailey-Serres, J. PHOSPHORUS-STARVATION TOLERANCE 1 (OsPSTOL1) is prevalent in upland rice and enhances root growth and hastens low phosphate signaling in wheat. Plant Cell Environ. 2023, 46, 2187–2205. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transporters | Putative Transport Function | Set-1 | Set-2 | Set-3 | ||||
|---|---|---|---|---|---|---|---|---|
| Gene Name | Gene ID | log2(FC) | p Value | log2(FC) | p Value | log2(FC) | p Value | |
| ZmNRAMP5 | Zm00001eb304610 | HMMs | 3.26 | 1.4 × 10−13 | 1.31 | 3.1 × 10−3 | 3.62 | 7.9 × 10−67 |
| ZmMRPA14 | Zm00001eb072510 | 5.31 | 2.6 × 10−68 | 2.84 | 6.5 × 10−11 | 3.77 | 6.6 × 10−73 | |
| ZmMATE11 | Zm00001eb048860 | 6.58 | 9.6 × 10−144 | 5.90 | 3.6 × 10−26 | 6.89 | 2.5 × 10−267 | |
| ZmMATE17 | Zm00001eb086100 | 4.65 | 1.9 × 10−57 | 3.29 | 8.7 × 10−6 | 3.38 | 5.0 × 10−115 | |
| ZmMATE30 | Zm00001eb006660 | 3.02 | 1.6 × 10−52 | 2.53 | 7.1 × 10−22 | 1.15 | 1.6 × 10−4 | |
| ZmMATE48 | Zm00001eb405390 | 1.95 | 3.9 × 10−14 | 1.18 | 3.4 × 10−3 | 1.37 | 3.3 × 10−24 | |
| ZmTIP1;1 | Zm00001eb003730 | Water | 2.18 | 2.7 × 10−19 | 1.69 | 8.7 × 10−4 | 1.45 | 1.1 × 10−37 |
| ZmSIP1-2 | Zm00001eb349840 | Water | 1.37 | 1.2 × 10−6 | 2.09 | 4.3 × 10−8 | 1.47 | 9.3 × 10−48 |
| ZmNCS2-2 | Zm00001eb042450 | - | 1.51 | 3.4 × 10−16 | 1.58 | 9.5 × 10−10 | 1.40 | 8.5 × 10−66 |
| ZmNPF1.1 | Zm00001eb152400 | Oligopeptide, phosphate ion | −2.35 | 1.7 × 10−30 | −2.26 | 2.1 × 10−5 | −1.57 | 1.2 × 10−14 |
| ZmCOPT2 | Zm00001eb151980 | Copper | −1.15 | 1.5 × 10−6 | −2.20 | 4.8 × 10−2 | −2.85 | 2.2 × 10−38 |
| ZmPIP2:2 | Zm00001eb096680 | Water | −4.33 | 9.7 × 10−6 | −3.34 | 3.9 × 10−12 | −1.44 | 2.5 × 10−32 |
| ZmTIP3-1 | Zm00001eb221090 | Water | −2.49 | 7.8 × 10−4 | −1.91 | 8.1 × 10−8 | −2.14 | 1.2 × 10−34 |
| cl34132_1a | Zm00001eb068600 | - | −2.12 | 1.3 × 10−50 | −2.11 | 3.5 × 10−12 | −1.13 | 1.8 × 10−13 |
| putative transporter | Zm00001eb323920 | Phosphate ion | −5.68 | 1.3 × 10−37 | −6.09 | 9.1 × 10−41 | −4.51 | 1.1 × 10−133 |
| ZmSWEET4c | Zm00001eb236810 | Glucose | −3.12 | 8.8 × 10−10 | −2.56 | 2.7 × 10−3 | −1.25 | 1.4 × 10−34 |
| ZmPHO1;2a | Zm00001eb191650 | Phosphate ion | −1.19 | 1.4 × 10−16 | −1.58 | 5.8 × 10−7 | −1.52 | 2.4 × 10−17 |
| ZmPHO1;2b1 | Zm00001eb258520 | Phosphate ion | −0.13 | 1.1 × 10−2 | −1.09 | 3.78 × 10−2 | −1.48 | 3.5 × 10−37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, D.; Pan, S.; Du, X.; Chen, E.; He, J.; Zhang, Z. Central Roles of ZmNAC128 and ZmNAC130 in Nutrient Uptake and Storage during Maize Grain Filling. Genes 2024, 15, 663. https://doi.org/10.3390/genes15060663
Peng D, Pan S, Du X, Chen E, He J, Zhang Z. Central Roles of ZmNAC128 and ZmNAC130 in Nutrient Uptake and Storage during Maize Grain Filling. Genes. 2024; 15(6):663. https://doi.org/10.3390/genes15060663
Chicago/Turabian StylePeng, Di, Shuxing Pan, Xin Du, Erwang Chen, Junjun He, and Zhiyong Zhang. 2024. "Central Roles of ZmNAC128 and ZmNAC130 in Nutrient Uptake and Storage during Maize Grain Filling" Genes 15, no. 6: 663. https://doi.org/10.3390/genes15060663
APA StylePeng, D., Pan, S., Du, X., Chen, E., He, J., & Zhang, Z. (2024). Central Roles of ZmNAC128 and ZmNAC130 in Nutrient Uptake and Storage during Maize Grain Filling. Genes, 15(6), 663. https://doi.org/10.3390/genes15060663