1. Introduction
Sex identification in dioecious plants, at an early developmental pre-flowering stage, is challenging; yet of great economical and biological importance, especially for species producing high-added-value products, such as the mastic tree (
Pistachia lentiscus L. var.
chia).
P. lentiscus L. is widely distributed in the maquis communities of the Mediterranean basin and the economically important variety
chia is grown in the island of Chios in the northern Aegean Sea. Mastic gum or Chios mastiha, is the resinous sap extracted from the tree trunk and the main branches [
1] as a reaction of the of the male mastic trees to wounding. Chios mastiha, which is known for its therapeutic properties, has been identified as a Protected Designation of Origin (PDO) product by the European Commission, 1997 [
2] and in 2014 the know-how of cultivating mastic trees in Chios was inscribed in the Representative List of the Intangible Cultural Heritage of Humanity (UNESCO), 2014 [
3].
Substantial advancements have been made mainly in
Pistacia vera, using conventional breeding and selection methods. However, there is limited scientific research on Pistacia wild species and specifically on
P. lentiscus [
4], albeit the great economic importance of
P. lentiscus var.
chia for production of the mastic gum, which is typically propagated vegetatively by hardwood shoot cuttings [
5] and breeding efforts are still in their infancy. Two major limiting factors of breeding capacity in the perennial
Pistacia species are the required long breeding cycles, and their dioecious nature, which entail additional costs, labor, time, and land for preserving and testing the breeding populations [
6,
7]. The use of various DNA-based molecular markers developed for
P. vera for the study of intra- and interspecific phylogeny, and marker-assisted breeding have only recently begun to be applied in
P. lentiscus mainly for male cultivar fingerprinting, genotype identification, and genetic diversity analysis in wild genotypes of
P. lentiscus.
A major breeding target in
P. lentiscus var.
chia would be the maximum productivity of male trees for high-quality mastic gum. The male mastic trees can reach full sexual maturity after 5–6 years, whilst mastic production reaches a maximum yield of 1 kg at 12–15 years [
8]. As such, it is essential for the mastic tree breeders, growers, and producers to distinguish the male trees in early growth stages. Therefore, early-stage sex identification is crucial given the importance of distinguishing the male seedlings in mass propagation systems, which produce higher quantities and better-quality of mastic compared to the female trees. Thus far, sex identification in
Pistacia species is unattainable during the long pre-reproductive phase of the trees, due to the lack of sex-related morphological methods at this developmental stage [
9,
10,
11]. The application of appropriate species-specific molecular markers could be an effective molecular tool and facilitate sex identification in mastic trees at the pre-reproductive phase.
Research on the sex determination mechanism in
P. vera L. has shown that the development of opposite sex primordia is initiated but then arrested in the early stages [
12]. Based on sex-associated loci identified in pistachio, a ZZ/ZW sex determination system has been reported, in which the females are the heterogametic sex [
6,
10,
13,
14]. Furthermore, there is evidence that the heterogametic sex chromosome system is emerging in the early stage of differentiation given that the heterozygous SNPs markers are present in pistachio females [
10]. Interestingly and despite the dioecious nature of
Pistacia species, monoecious instances with either male and female inflorescences on different branches, or on the same branch or mixed inflorescences and bisexual flowers, have also been previously observed in wild species [
15], such as in
Pistacia chinensis Bunge [
16],
Pistacia atlantica Desf. [
17,
18], and
Pistacia terebinthus [
19]. Similar cases have also been observed in
P. lentiscus L. trees on the island of Chios (Chios Mastic Gum Growers Association).
Sex identification in
Pistacia species has largely focused in
Pistacia vera using various sex-linked DNA markers, such as different Random Amplified Polymorphic DNA (RAPD) markers in
P. vera [
9,
20,
21],
Pistacia eurycarpa and
P. atlantica [
17]. More specifically, the OPO-08 decameric Operon primer (RAPD), successfully distinguished the female individuals in
P. vera, resulting in a 945 bp amplification product [
20]. However, in other instances, sex discrimination with this RAPD marker was not possible in
P. vera [
22]. In a later study, Yakubov et al. [
23] transformed the OPO-08 RAPD primer into Sequence Characterized Amplified Region (SCAR) marker that enabled the discrimination between male and female individuals in different
P. vera genotypes [
23] and in other
Pistacia species [
9,
21]. Based on these studies, the low frequency of sex-linked bands observed indicates that the genetic loci involved in sex determination are small and probably involve a single or very few genes [
9,
20]. Testing the SCAR marker in
P. vera gave false-negative and false-positive results in females and males, respectively, and was not effective in distinguishing sex in other
Pistacia species [
7]. Effective sex identification of female and male individual
P. vera cultivars has been also achieved using Inter-Simple Sequence Repeat (ISSR) markers [
24]. Sex-linked polymorphic EST-SSR markers were also effective in eliminating males in
P. vera yet were not able to successfully identify the sex in other Pistacia species [
6], except for the transcriptome-based EST-SSR marker identified by Chang et al. [
25], which was effective in distinguishing between male and female
P. chinensis.
The development of species-specific markers for sex identification in
P. lentiscus var.
chia is still lacking, especially at the juvenile period. The use of ISSR and RAPD markers for genetic diversity studies in
P. lentiscus L. have also shown a great potential for sex identification, whereas SSRs were not as effective in distinguishing male from female individuals [
26]. Specifically, there is evidence that male and female plants were distinguished using RAPD and ISSR markers in
P. lentiscus L. (12 females vs. 12 males) [
26] and
P. lentiscus L. var.
chia genotypes, (one female vs. 10 males) [
27]; 10 females vs. 10 males) [
28], however these studies used only a limited number of male and female individuals, thus further work is required to be able to confidently differentiate male from female individuals in
P. lentiscus and especially in the variety
chia.
To achieve the maximum economic benefits from the cultivation of P. lentiscus var. chia, it is necessary to distinguish males from females at the seedling stage through early sex diagnosis tools. Herein, genetic markers such as ISSRs, RAPDs and SCARs, which have shown promising results in other closely related Pistacia species, were used to assess the efficacy in sex differentiation between female and male mastic trees. These markers were further used to assess potential links between the sex identity and the genetic diversity of different mastic tree cultivars. Additionally, transcriptome-based markers were developed as an alternative for early-stage sex diagnosis in P. lentiscus var. chia using male, female, and monoecious individuals. The results of this study are promising as a desirable tool for accelerating the selection process for male seedlings at an early stage and reducing the costs of and time required in breeding programs.
4. Discussion
P. lentiscus var.
chia is of high economic importance for the pharmaceutical properties of the mastic gum, which is produced by the male trees in the island of Chios. However, the male mastic trees can reach full sexual maturity after approximately six years, when mastic production begins. Thus, distinguishing the male trees in early growth stages is important aim for the mastic tree breeding attempts, considering that genetically assisted breeding for productivity prerequisites sex identification of trees prior to the development of productivity associated genetic markers. Sex identification in Pistacia species has mainly focused on
P. vera L. and other related species, such as
P. eurycarpa and
P. atlantica [
6,
7,
9,
14,
17,
20,
21,
43] using various sex-linked DNA markers. Despite the importance of
P. lentiscus var.
chia, the genome of this species is still largely unknown, and thus far the use of
P. vera as reference genome is available. Limited studies have also been performed mainly for genotyping in
P. lentiscus genotypes with the use of ISSR, RAPD and SSR markers [
26] and
P. lentiscus var.
chia using ISSR and RAPD markers [
27,
28]. To our knowledge, this is the first report regarding: (i) transcriptome-based marker coupled with HRM analysis specifically developed for this species and especially for the
chia variety, using male and female populations along with identified monoecious individuals, to demonstrate a cost-effective method for sex identification that can be applied with minimum infrastructure and (ii) effective sex differentiation in
P. lentiscus var.
chia with 100% accuracy using a combination of different sex-linked ISSR, RAPD and SCAR markers. Additionally, investigating whether sex identity is associated with genetic variations and population structure we were able to examine the genetic relations amongst phenotypic cultivars of
P. lentiscus var.
chia.
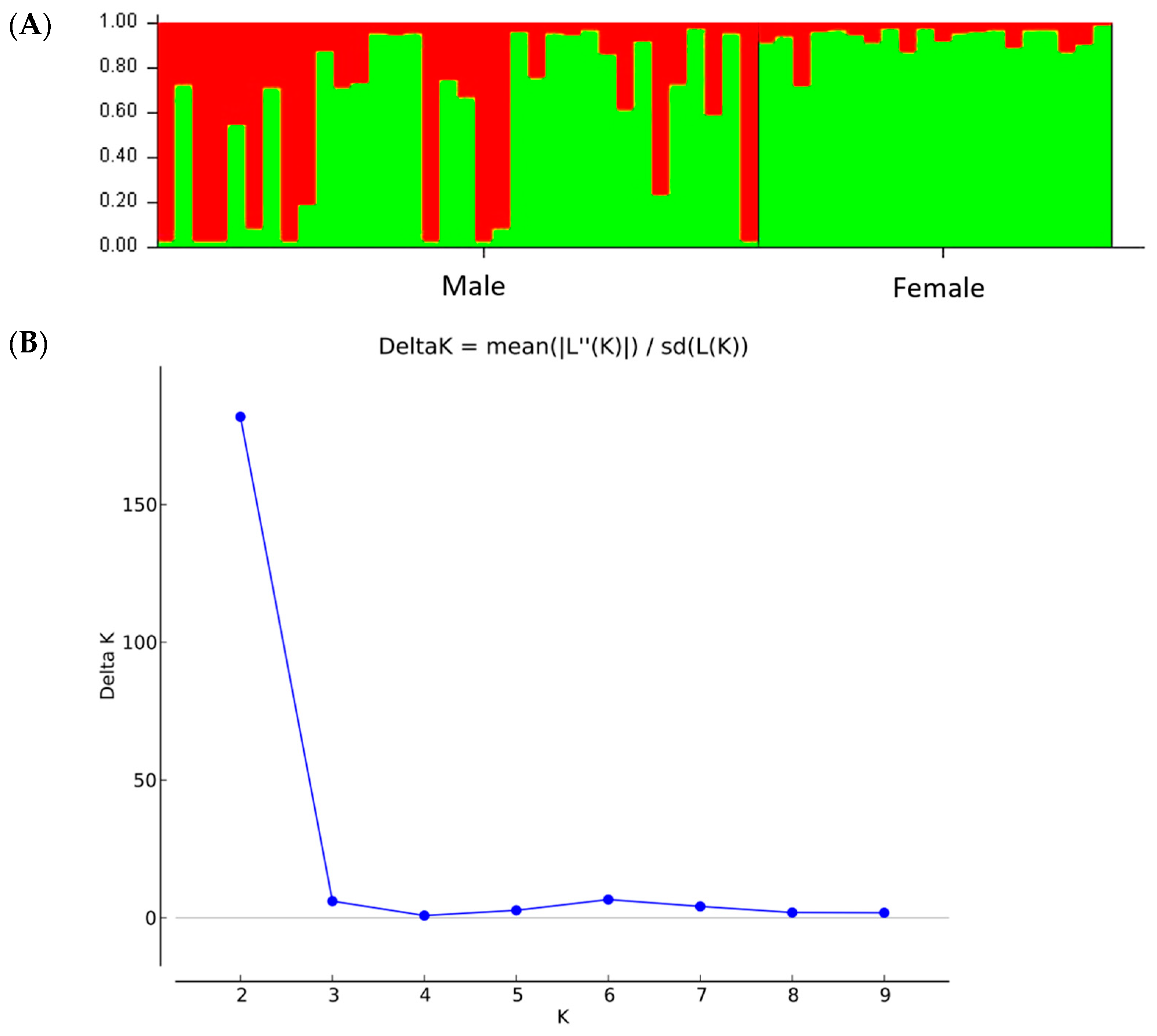
Our results indicated 100% accuracy on sex differentiation in
P. lentiscus var.
chia genotypes based on a combination of one SCAR and nine ISSR markers. These markers have been previously used either in
P. vera, such as the SCO-08 [
23], (AC)
8CG and (AC)
8TA [
44], or in
P. lentiscus var.
chia, such as the UBC811, UBC827, UBC834 UBC841, UBC842, and UBC856 [
27,
28] along with the UBC880, which we also included in this study. Our results are in accordance with the effective sex separation on
P. lentiscus genotypes based on ISSR markers [
26]. The 169 loci for a Φ
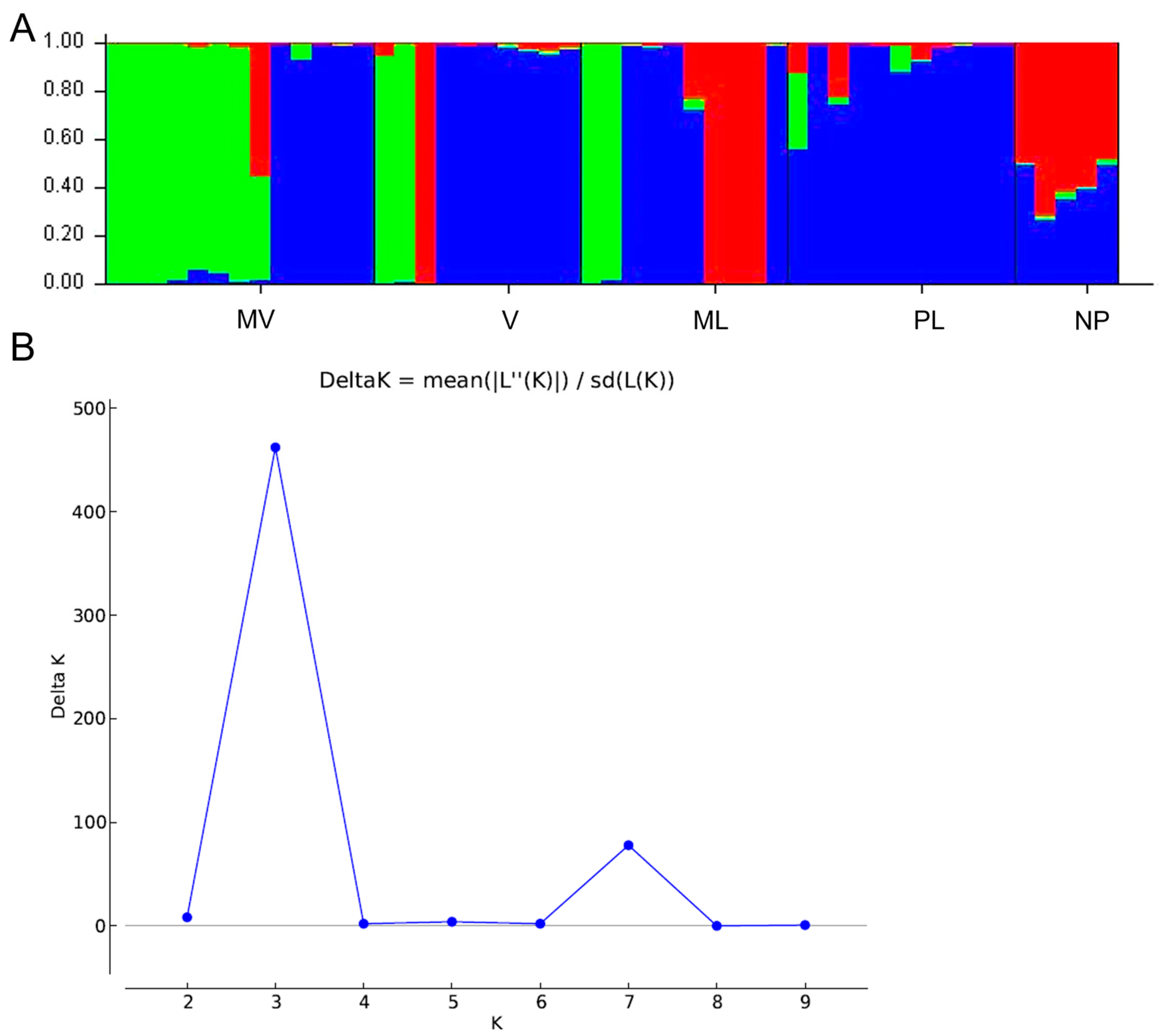
ST at 0.263 were more than adequate for estimating population structure [
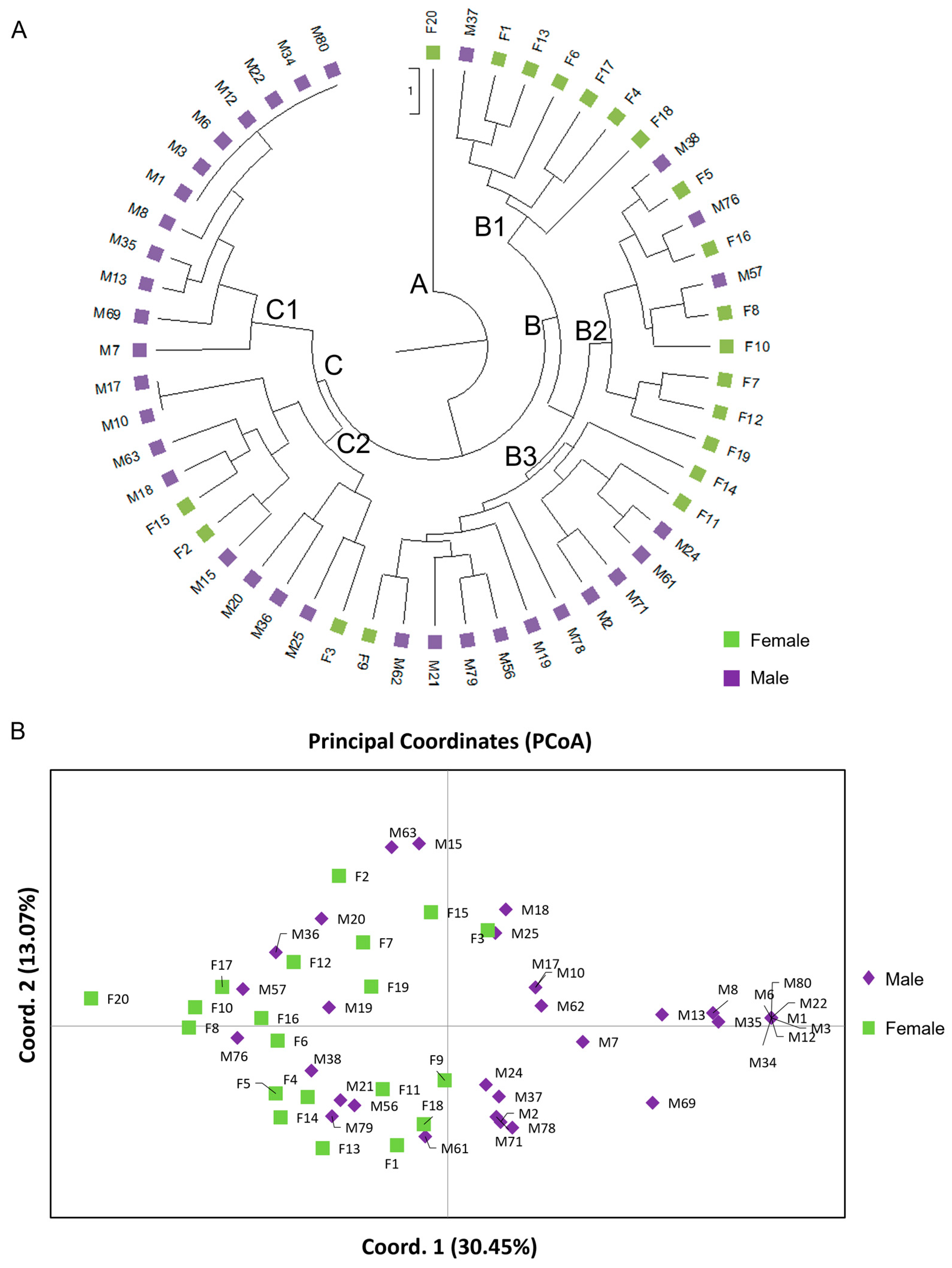
45]. The population structure analysis was in accordance with the PCoA and the UPGMA dendrogram demonstrated a clear separation of male and female genotypes. Interestingly, the non-productive males formed a distinct group, closer related to the male individuals. The male individuals showed higher genetic diversity compared to the females, which exhibited reduced intrapopulation variability. Comparable results were also demonstrated in genetic diversity studies in different male cultivars and female
P. lentiscus var.
chia genotypes based on ISSR markers [
27,
28].
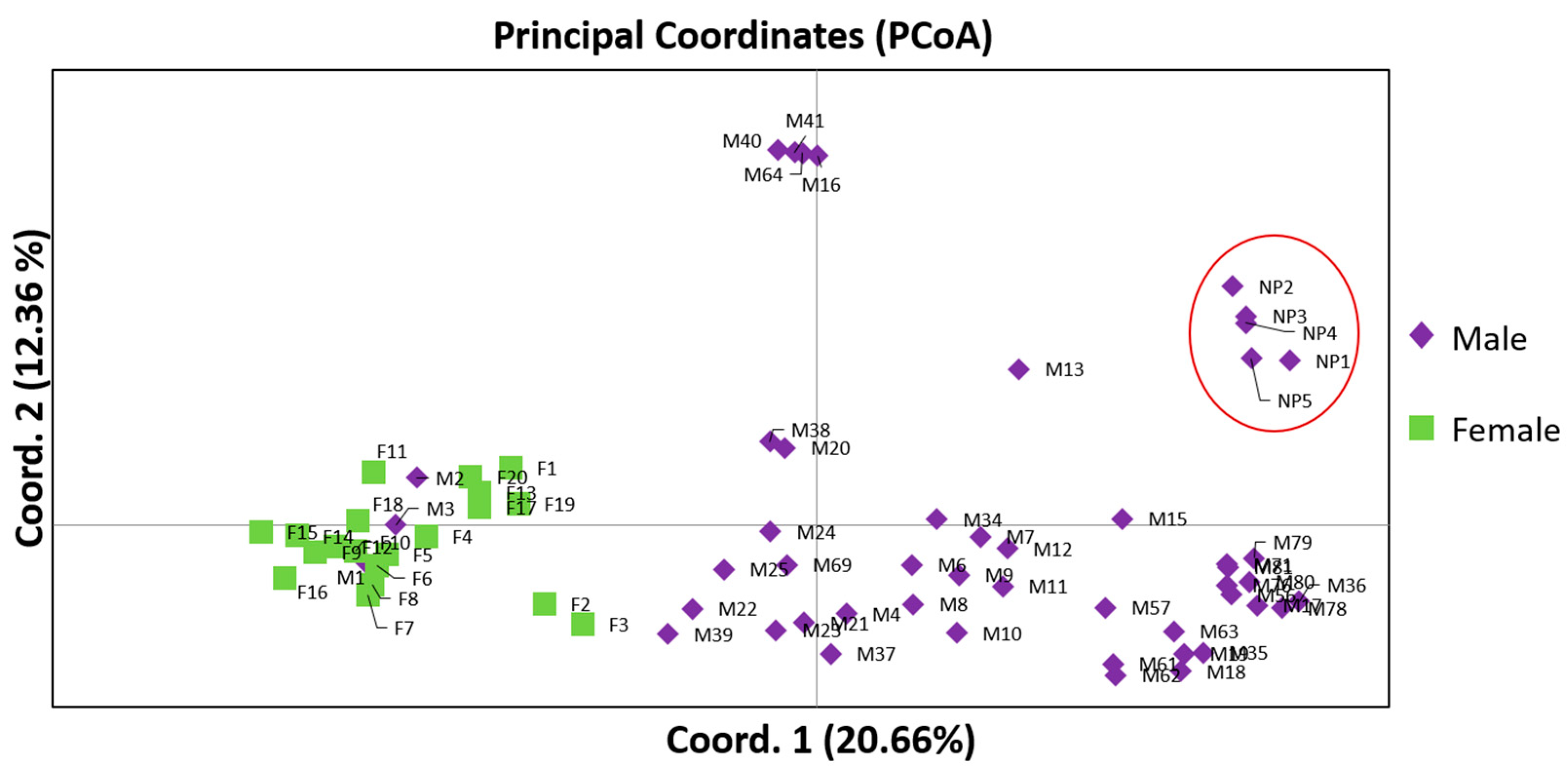
Sex-related differences between male and female individuals masked the genetic diversity among 20 females and the 49 males previously assigned to the 5 phenotypic cultivars,
Mavroschinos (MV),
Votomos (V),
Maroulitis (ML), and
Platiphyllos (PL). This effect has also been observed in
P. vera, 20 male and 20 female genotypes where the genetic distance was found to be mainly affected by the sex type rather than the intrapopulation distances [
24]. Herein, the female individuals showed closer genetic association to most of the males assigned to
Mavroschinos along with two males of the
Votomos and
Maroulitis cultivars, whereas the males of
Platiplyllos were further distant. This can be attributed to the higher diversity demonstrated by the
Mavroschinos,
Maroulitis and
Votomos cultivars compared to
Platiplyllos. Additionally, the non-productive (NP) individuals demonstrated low genetic diversity and were again clustered separately from the male population and not assigned to any phenotypic cultivar. Comparatively to our results, studies on genetic diversity of the same morphological cultivars confirm the existence of genetic heterogeneity [
27,
28], which is possibly the effect of either vegetative propagation or intercrossing indicating hybridization.
Based on genetic structure analysis, three main genetic groups were formed, (i) Mavroschinos, which seems to share genetic material with males of the Votomos (M2 and M8) and Maroulitis (M9 and M10) cultivars, possibly a result of mis-assigned individuals to these cultivars, (ii) Platiphyllos, which was the most homogeneous cultivar and (iii) Maroulitis with a cluster including M40, M41, M64 and M16, which has been possibly mis-assigned as Votomos. Furthermore, Votomos and Platiphyllos seem to interact and be genetically related, which could possibly indicate that these genotypes may have derived from vegetative propagation or may have a common genetic background. The non-productive population was also more homogeneous and presented a moderate level of admixture indicating a degree of intercrossing between Maroulitis and Platiphyllos and possibly Mavroschinos. Interestingly, Mavroschinos and Maroulitis presented three and one unique bands, respectively. The unique specific bands amplified in the two cultivars could potentially help in certification of these cultivars, yet further research, in a large germplasm collection, is required for validation of reliable markers.
Sex-linked marker identification has been mainly reported in
P. vera, mainly given the existence of molecular tools and the high-quality genome assembly [
14], yet not with 100% accuracy in other related species [
6,
25]. Research on marker development for sex identification in Pistacia genus has mainly focused on simple sequence repeat (SSR) in
P. vera and their transferability in other Pistacia species [
46], expressed sequence tag-derived (EST)-SSR in
P. chinensis [
25] and single nucleotide polymorphism in
P. vera (SNP) [
7]. Herein, we report the first EST-SSR and DEG-based markers’ design on male, female and monoecious
P. lentiscus var.
chia genotypes and their application using RT-PCR coupled with high-resolution melting (HRM) analysis for a cost-effective approach. Our results showed that the genetic diversity of male and female populations was relatively high and that male populations demonstrated higher levels of admixture. Interestingly, most of the non-productive male individuals were grouped with the females, whereas monoecious individuals were grouped with the males. Monoecious trees possibly originated from female trees [
15], given that in the ZW/ZZ sex-determination system the females are the heterozygotes [
14]. Studies on monoecious occurrence have shown that gender types (male, female or monoecious) in
P. chinensis Bunge were unstable in successive years [
16], and this could potentially be attributed to sexual plasticity as a response to environmental stresses and sex determination mechanism [
15].
The designed novel primers did not overlap with any of the primers reported in literature [
10,
15,
20,
47]. The eight most informative markers comprised six EST-SSRs and two DEG-based markers. More specifically, the six EST-SSRs based novel markers developed in the present study, were all found on DNA binding proteins, that are known for their relation to sex determination and flowering pathways in plants. Several MYB family members have been shown to play a role in the regulation of several cellular processes, as well as in responses to biotic and abiotic stress [
48], and in sex determination [
30]. Interestingly, the association of MYB with the SWI/SNF ATP-dependent chromatin remodeling complex could also indicate a possible temperature dependency of the sex determination in mastic trees [
49]. The
SUF4 is a suppressor of
FRI4 and a transcription factor (TF) required for delayed flowering in winter-annual Arabidopsis and has been characterized as a differentiation and developmental protein, as well as a DNA-binding TF with an activity in histone H3-K4 methylation [
50]. The
SPL9 are Squamosa Promoter Binding-Like Proteins and SBP TF, that have been connected to early flowering and other developmental functions [
51]. The
ELF6 is an Early Flowering (6) TF of the jumonji (jmj) protein family, which is involved in histone demethylation, resetting the embryo epigenetic memory [
50]. Lastly, the
Rpr2 is a DOF-zinc finger DNA-binding domain with a TF function involved in flowering regulation and responses to biotic and abiotic stresses [
52].
The two DEG-based markers that showed significant polymorphism information content (PIC) values, ACS1 and ACS2, were both found on a homologous to the acetyl-CoA synthetase (
ACS) gene, a gene that was found to be significantly downregulated in productive male individuals. ACS is peculiar in a sense that it is not a DNA binding protein, and it is mostly related to metabolism and not to sex differentiation. ACS in plants has been shown to be a component of a two-enzyme system that plants use to maintain acetate homeostasis [
53]. Our findings regarding ACS could indicate an acetate homeostasis dependency of sex determination that is visible on a transcription level. Interestingly, acetyl-coA has been found to play a significant role in many regulatory processes associated with acetylation of controlling components (such as histone acetylation or N-terminal and/or amino acid side-chain acetylation). Hence, the post-translational acetylation mechanism may enable plants to respond to shifting environmental and developmental conditions, which could affect the sex determination whilst controlling the acetyl-CoA and acetate balance [
53]. Regarding the lack of visible sequence differences between the sexes for ACS1 on a transcript level it might indicate a differential mRNA maturation, that could affect the half-life of the mRNA and thus, its detection and translation.
The percentage of true positives for sex identification was more than 80% for males and 70% for females. It is noteworthy that sex-linked SNP loci markers were unable to successfully identify the sex in wild
Pistacia species, including
P. lentiscus [
6,
7]. Additionally, the transferability of sex-linked markers identified in
P. vera was found to be the lowest in
P. lentiscus L. [
46], which could be due to the larger genetic distance between the two species [
11,
54]. This could be an effect of the genetic or epigenetic control of sex regulation that may alter the related gene(s) [
55]. Therefore, the novel nature and the ability of the transcriptome-based markers to accurately assign the true positive male and female trees could provide insights in the sex determination mechanisms of
P. lentiscus.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}