
Effects of Long-Term Cryopreservation on the Transcriptomes of Giant Grouper Sperm
 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sperm Collection
2.3. Sperm Cryopreservation and Thawing
2.4. RNA Extraction, cDNA Library Construction, and Sequencing
2.5. Data Analyses
2.6. Sample Time-Series Analysis
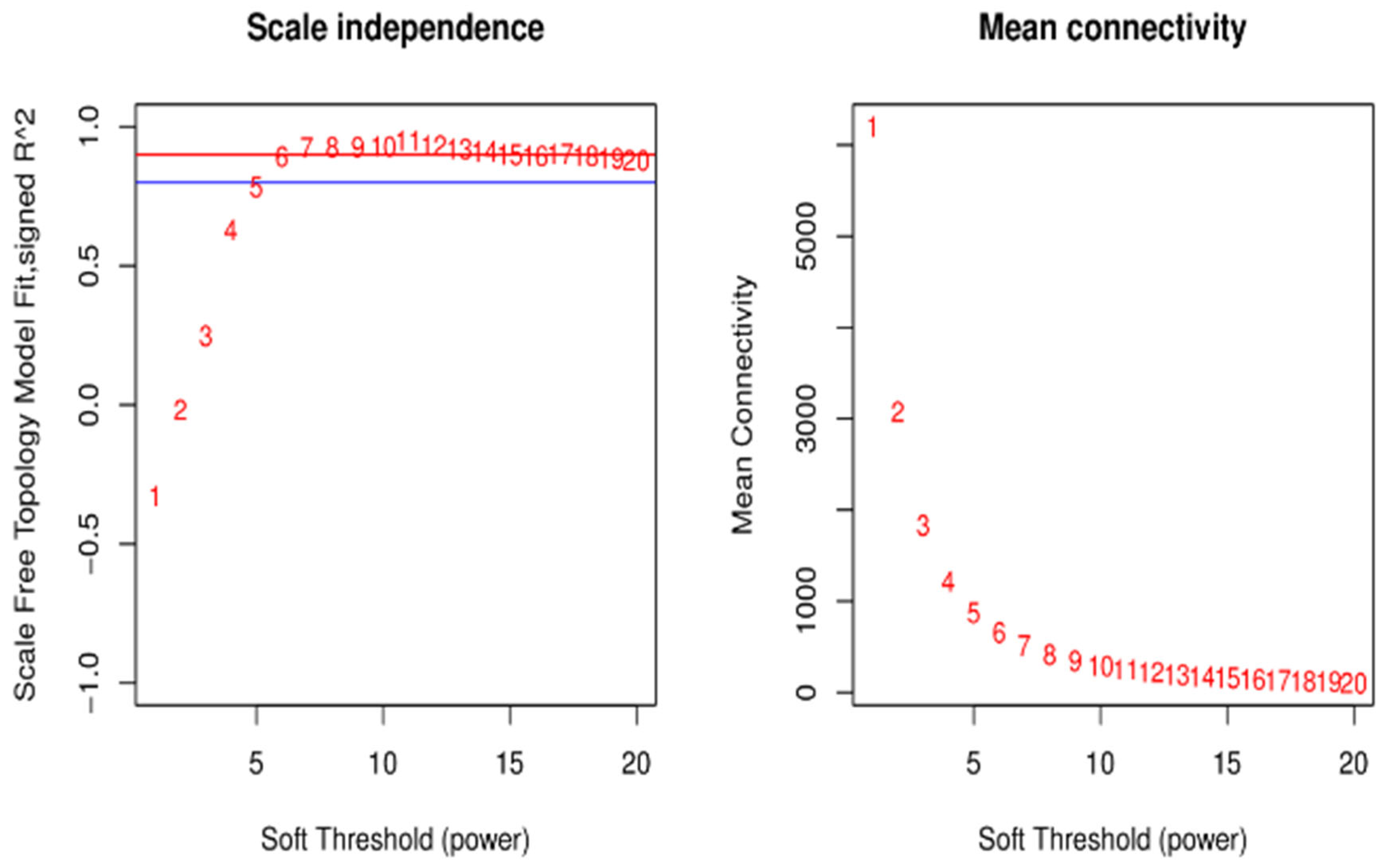
2.7. Weighted Gene Co-Expression Network (WGCNA) Construction
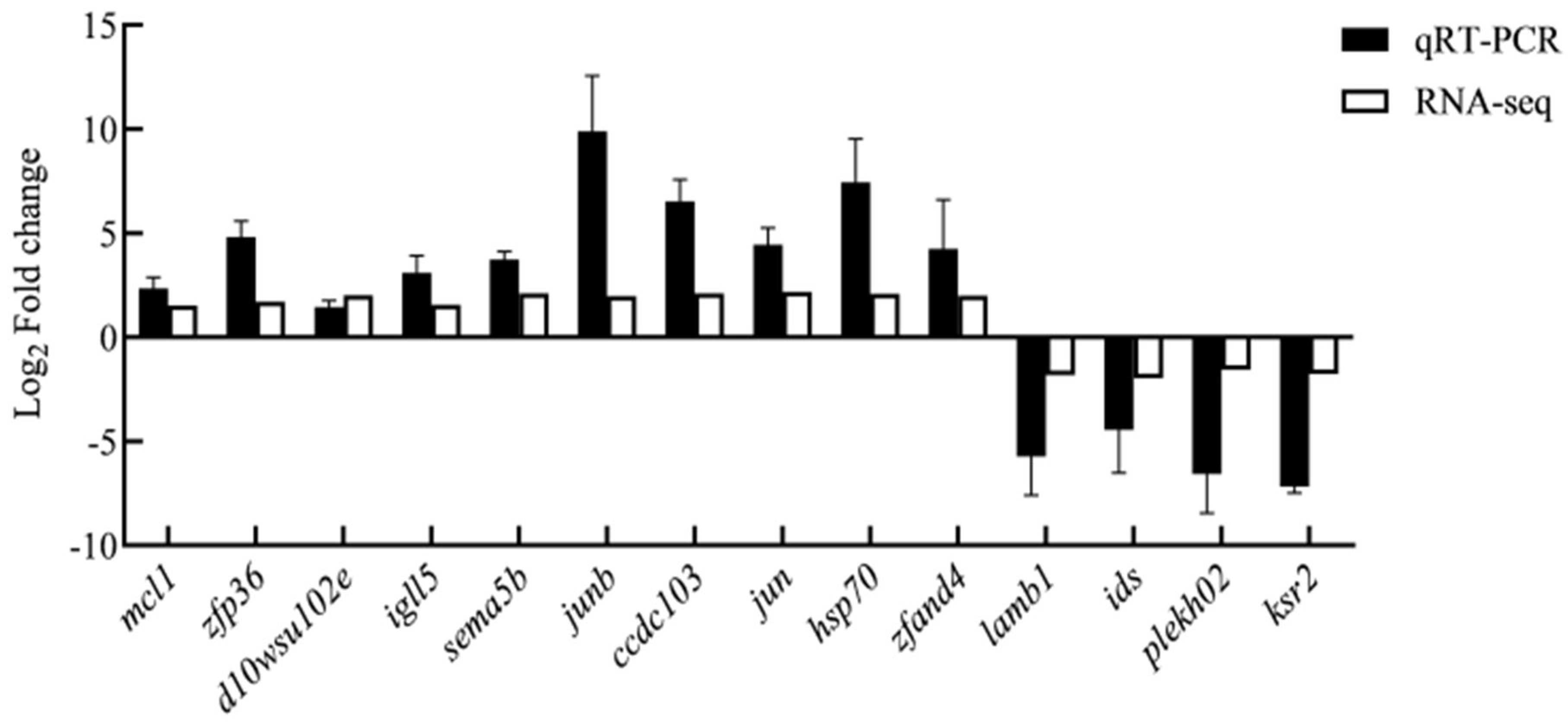
2.8. Experimental Validation Using qRT-PCR
3. Results
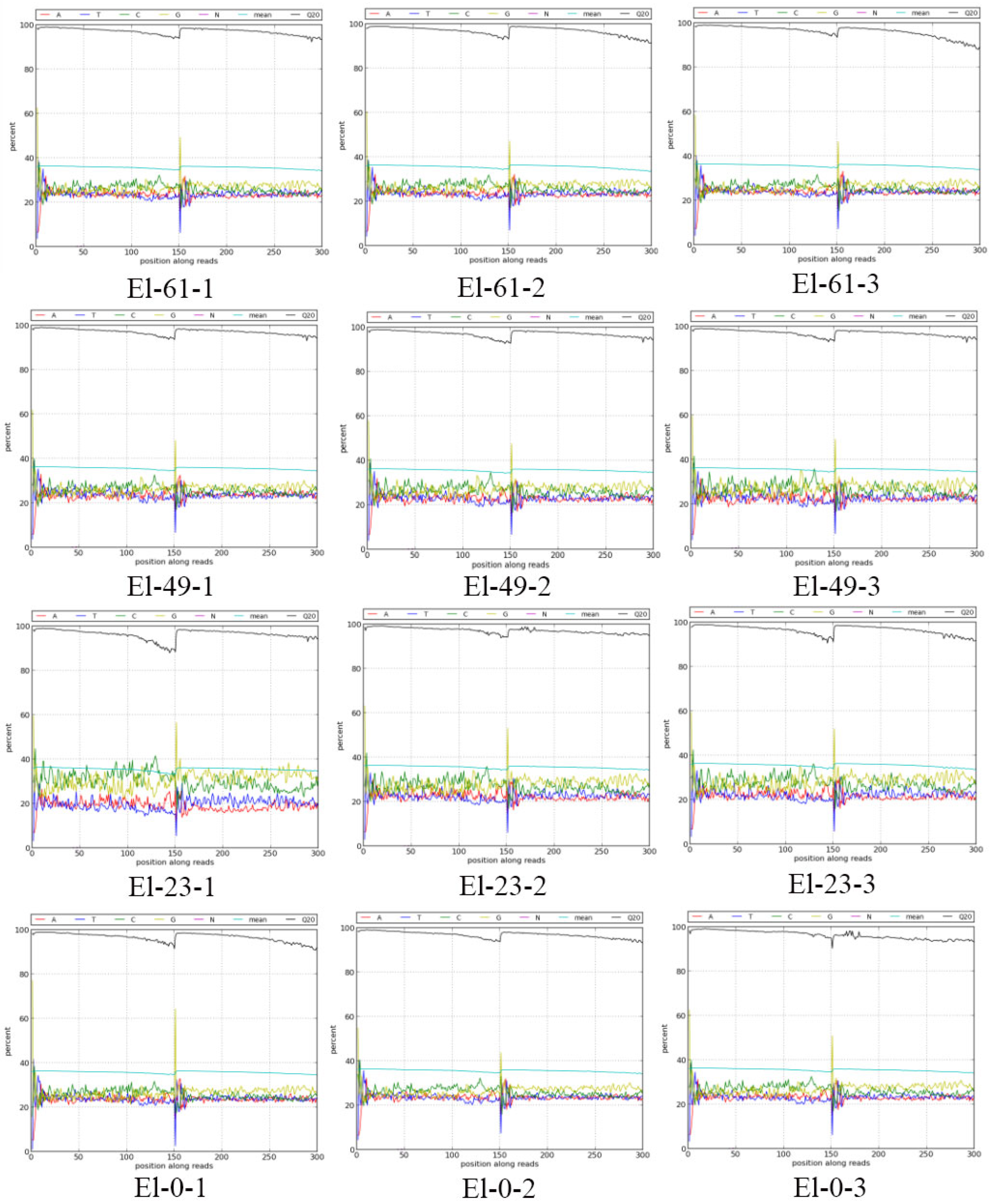
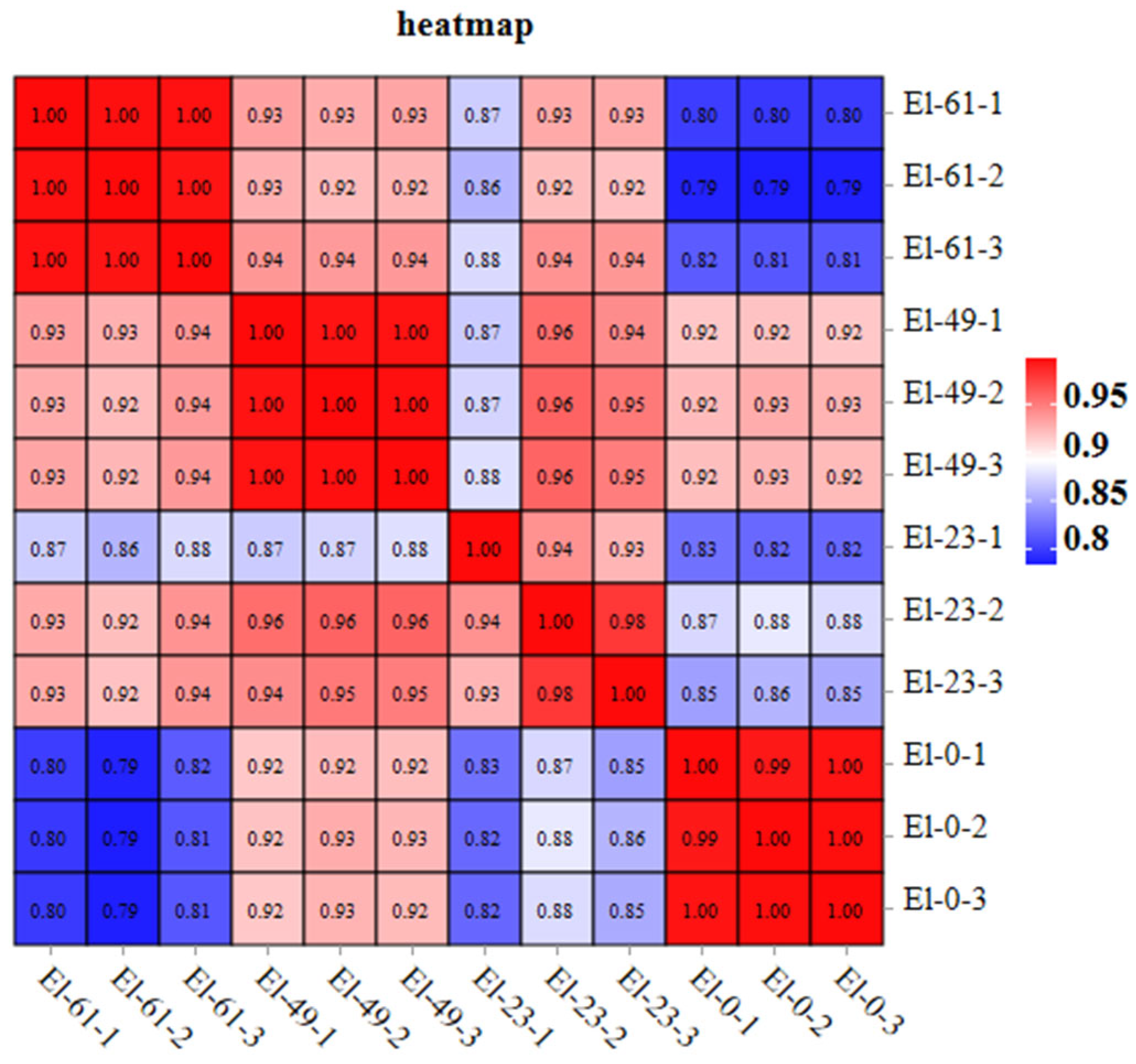
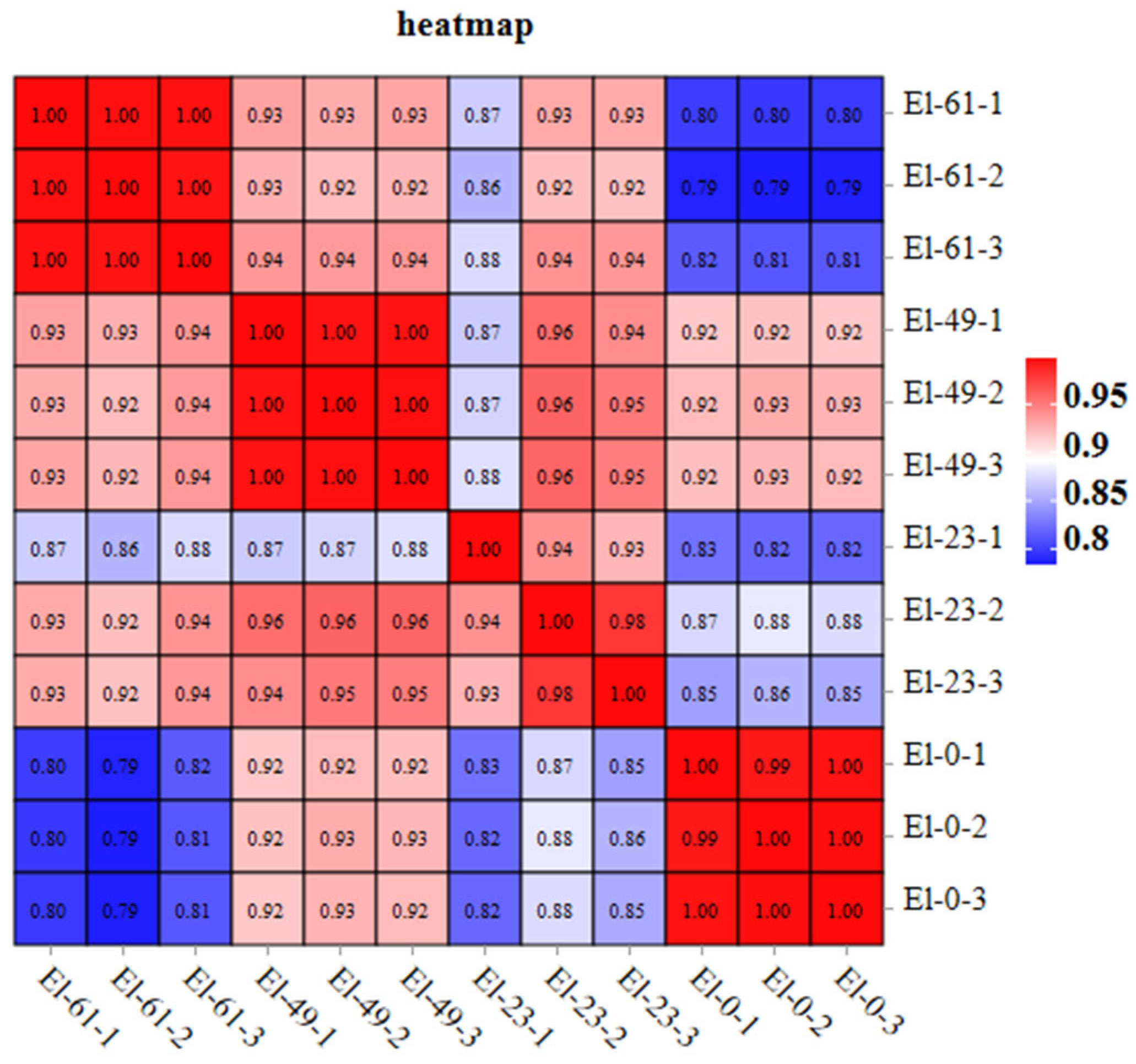
3.1. Transcriptome Sequencing and Assembly
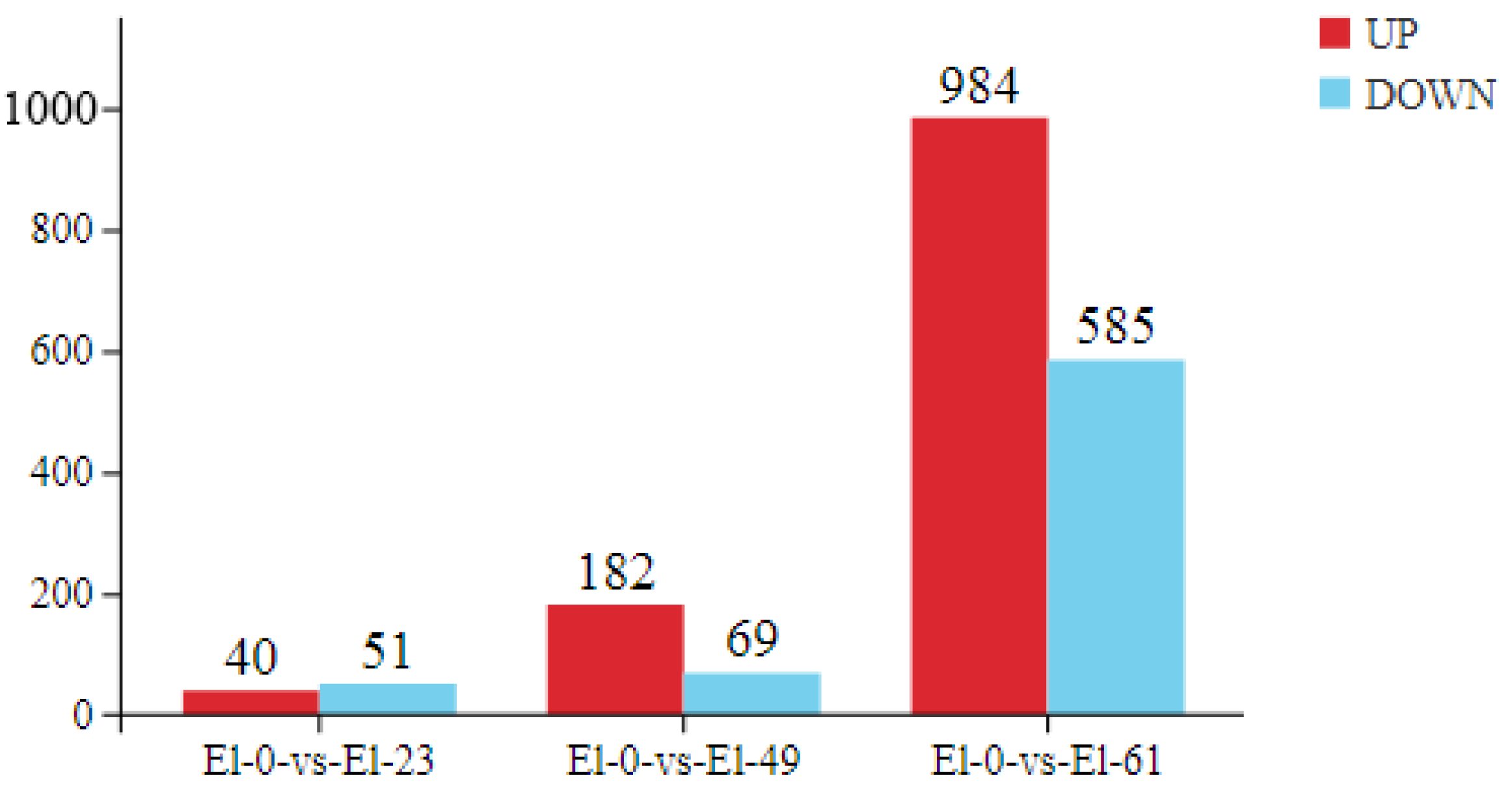
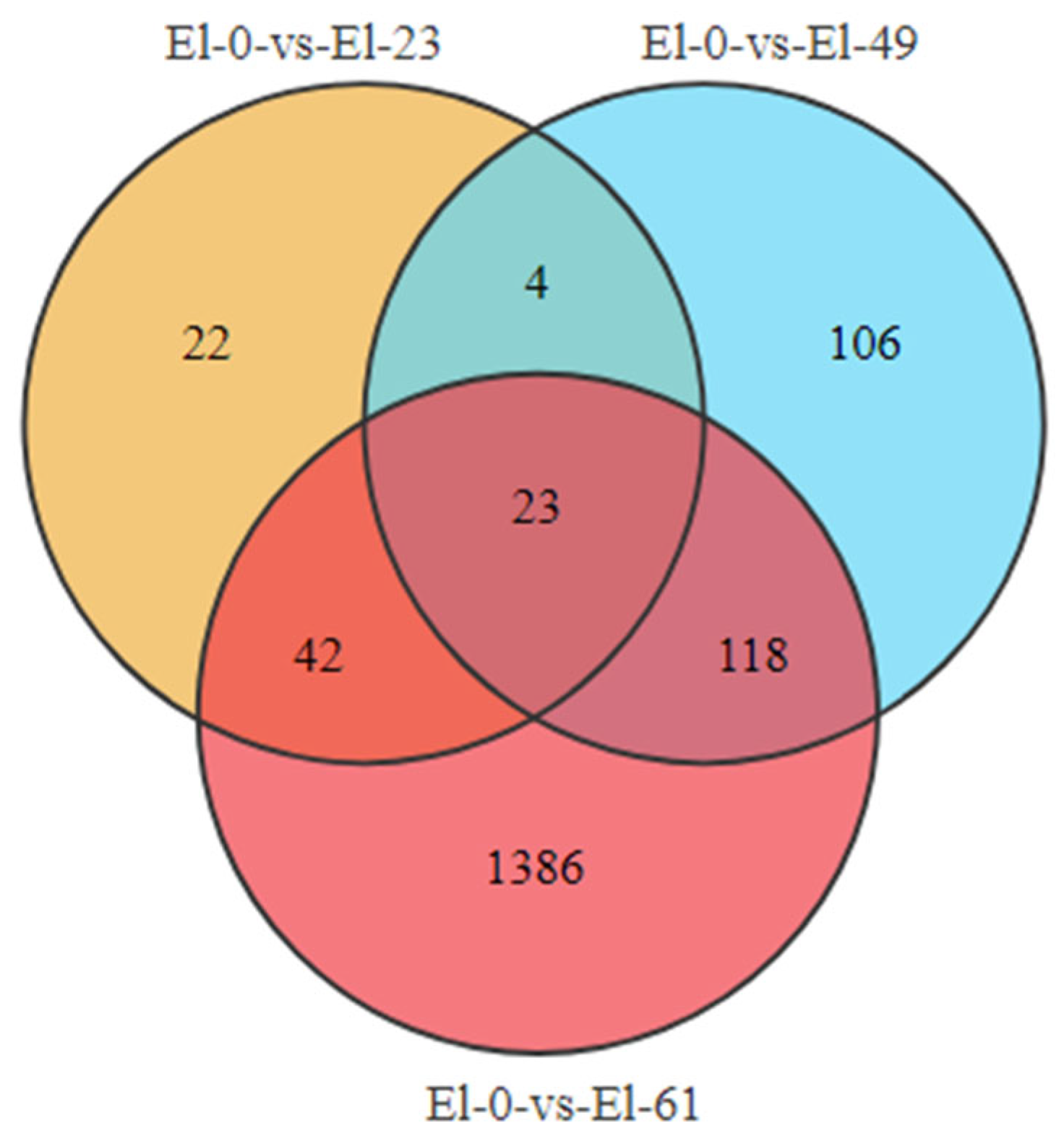
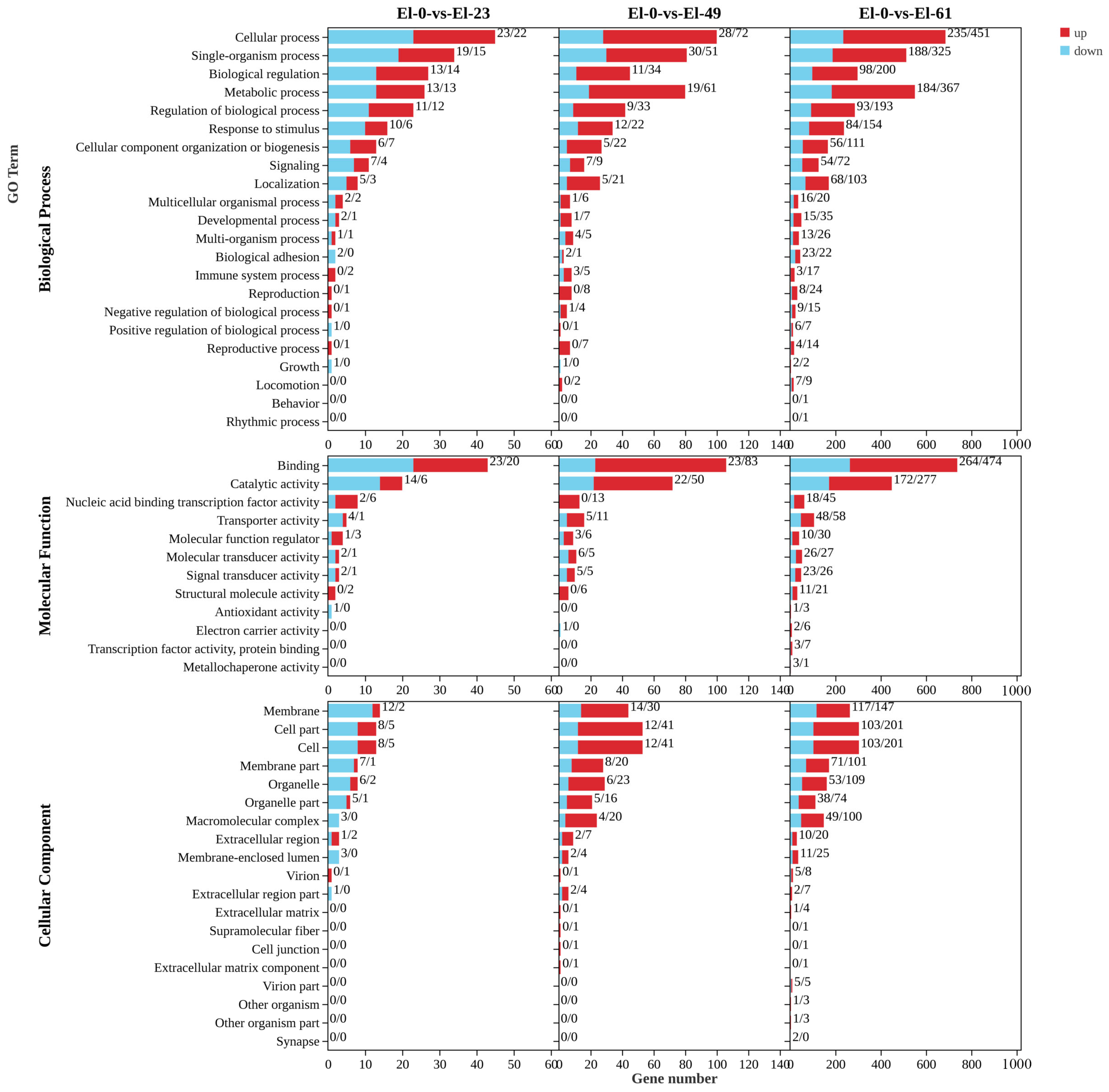
3.2. Analysis of Differentially Expressed Genes
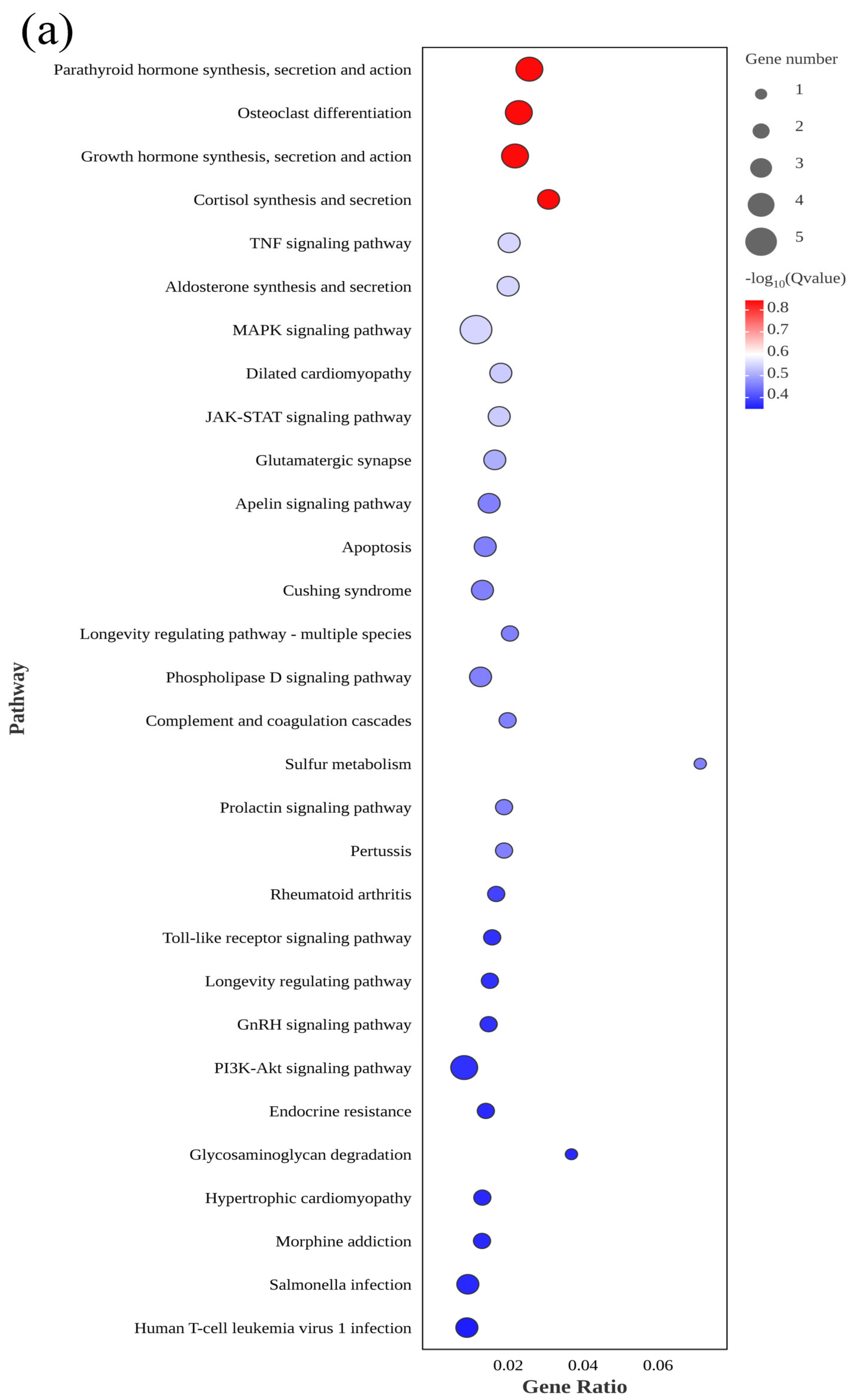
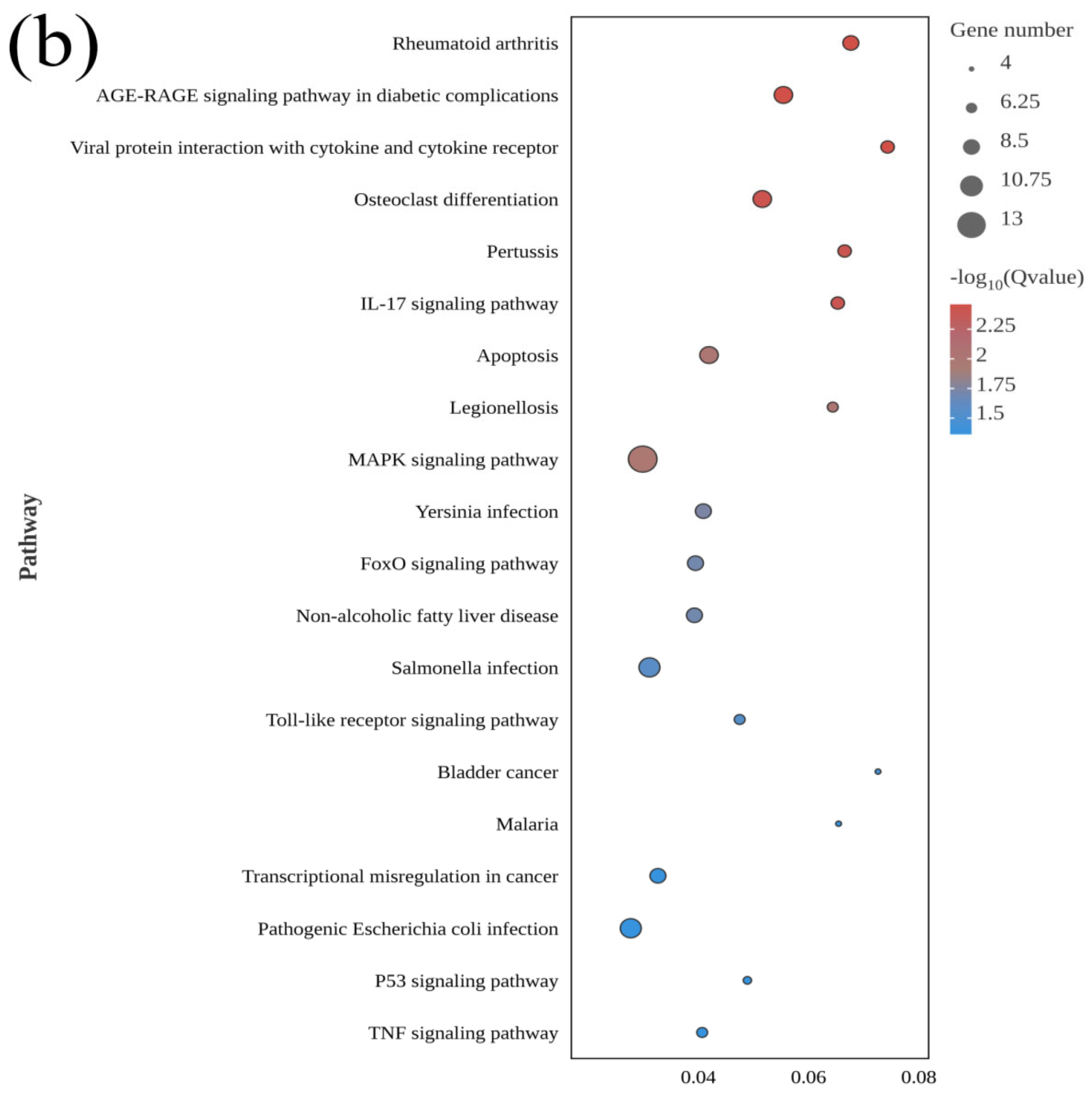
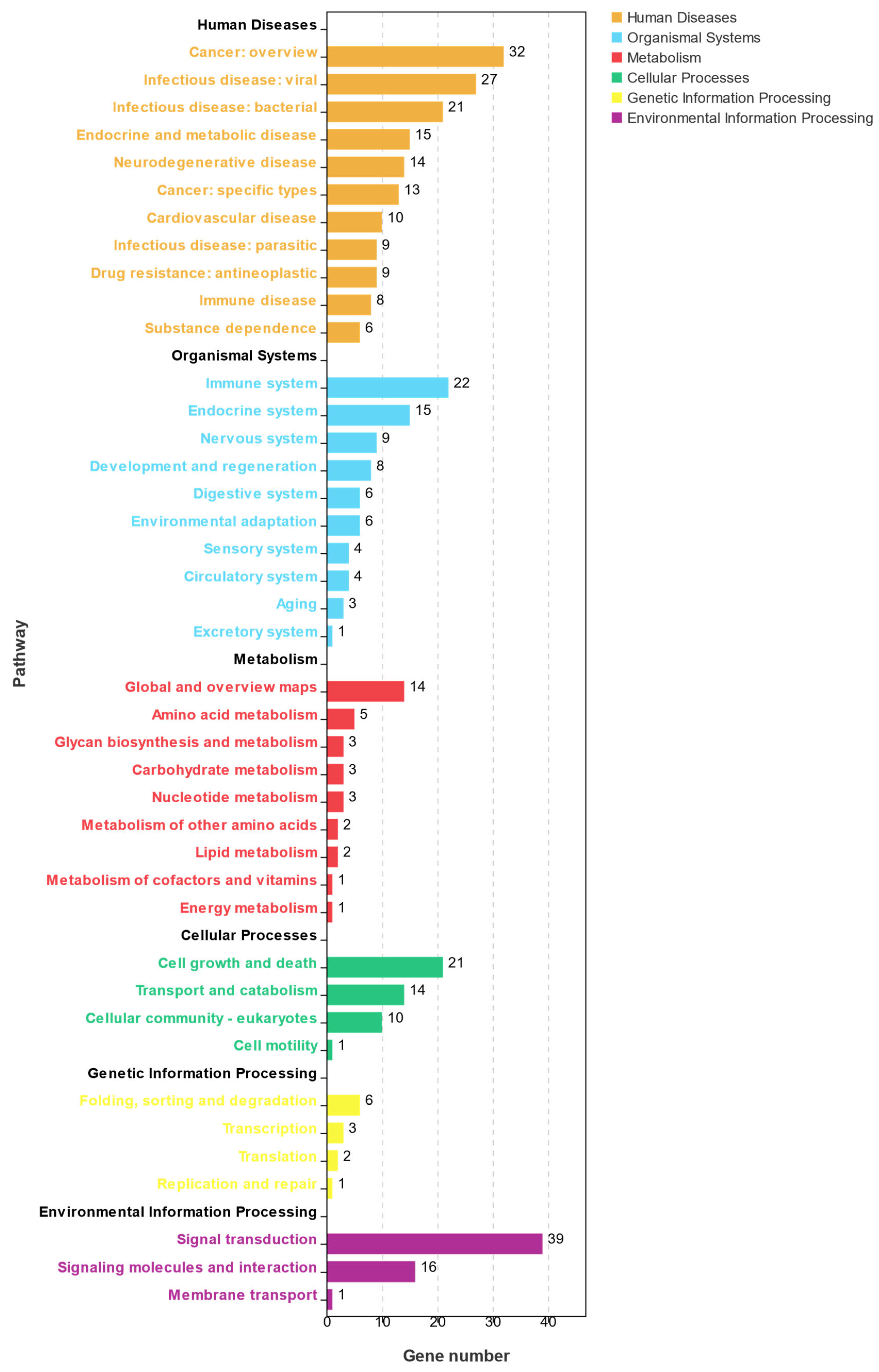
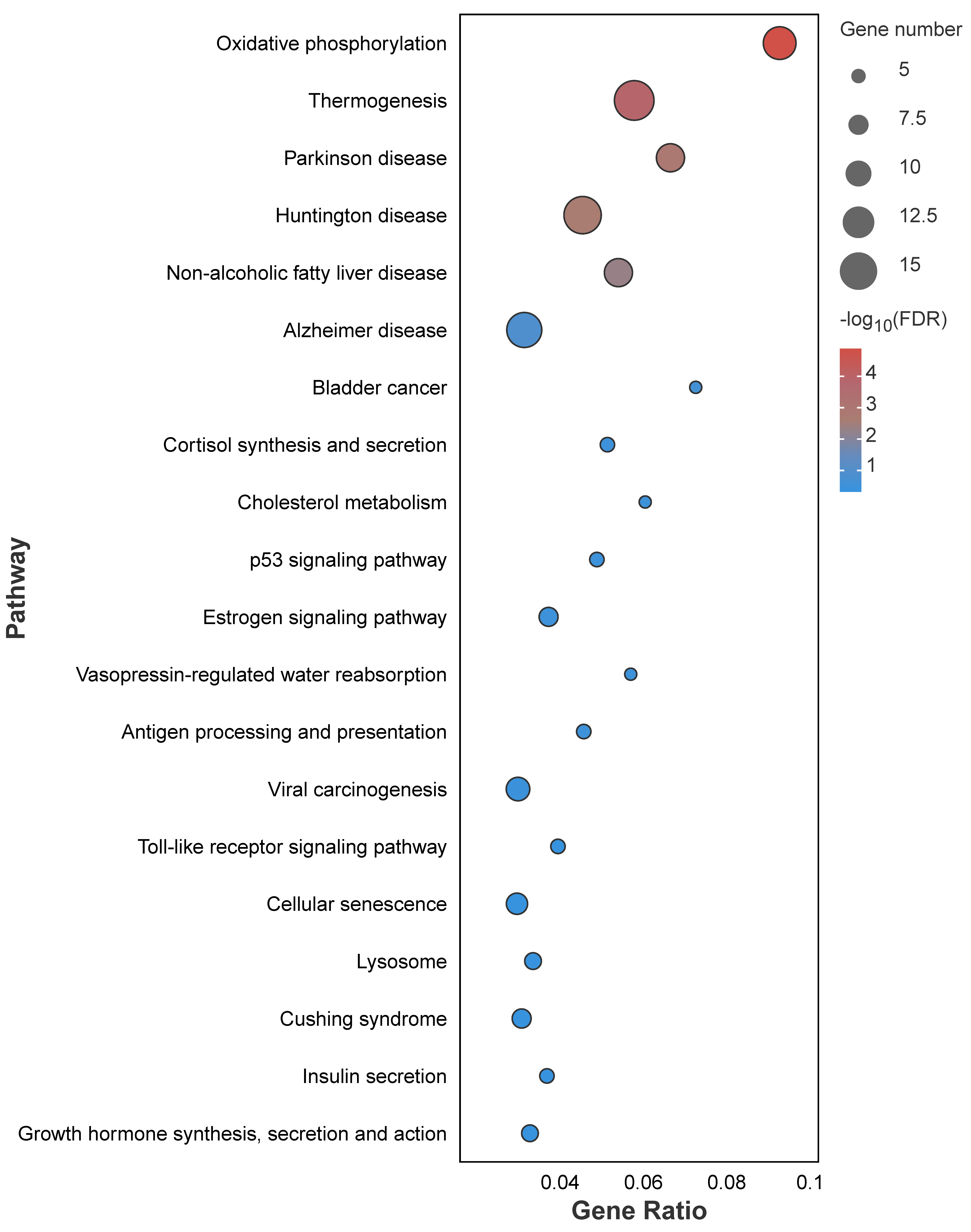
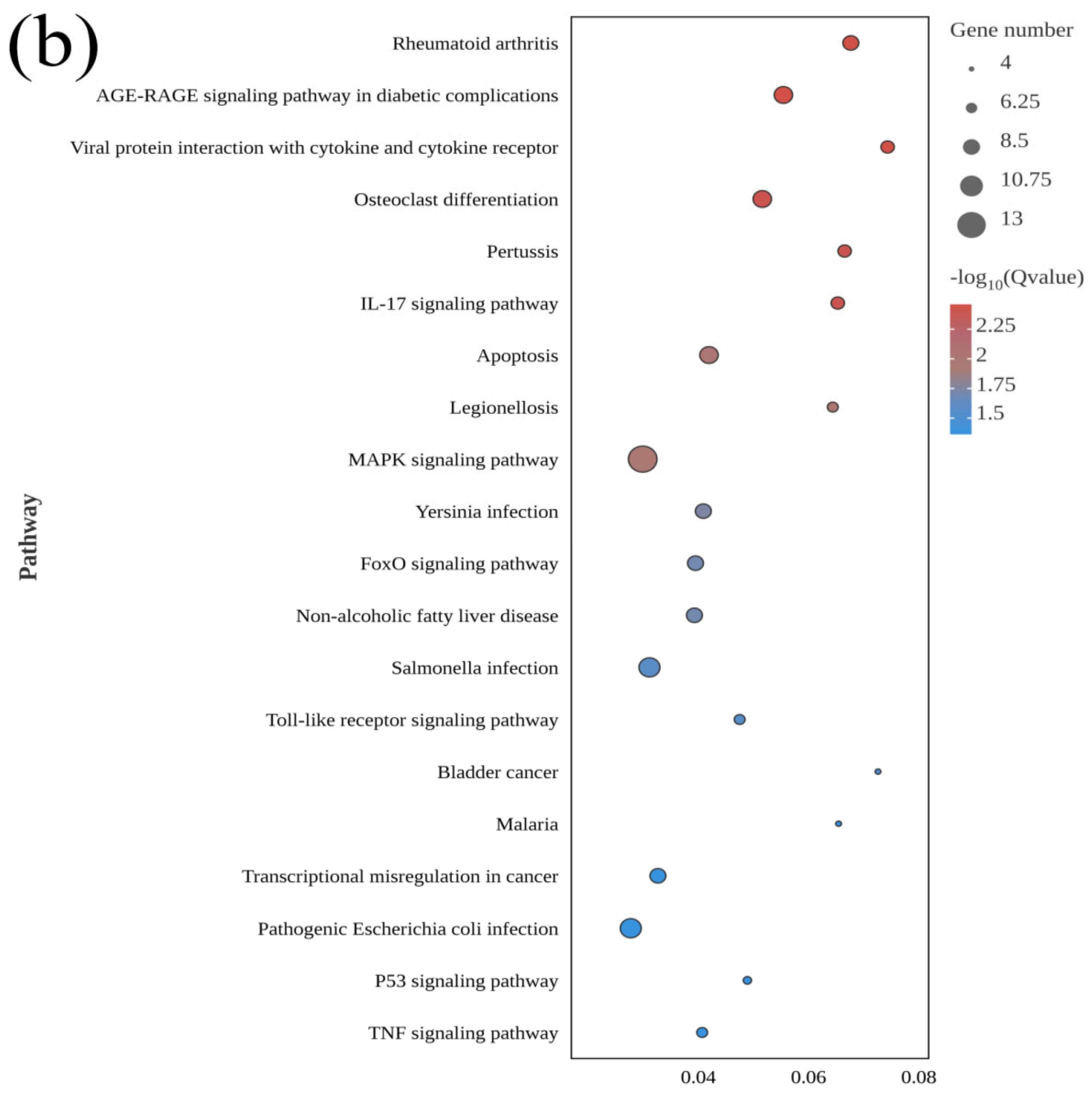
3.3. GO and KEGG Analyses of DEGs
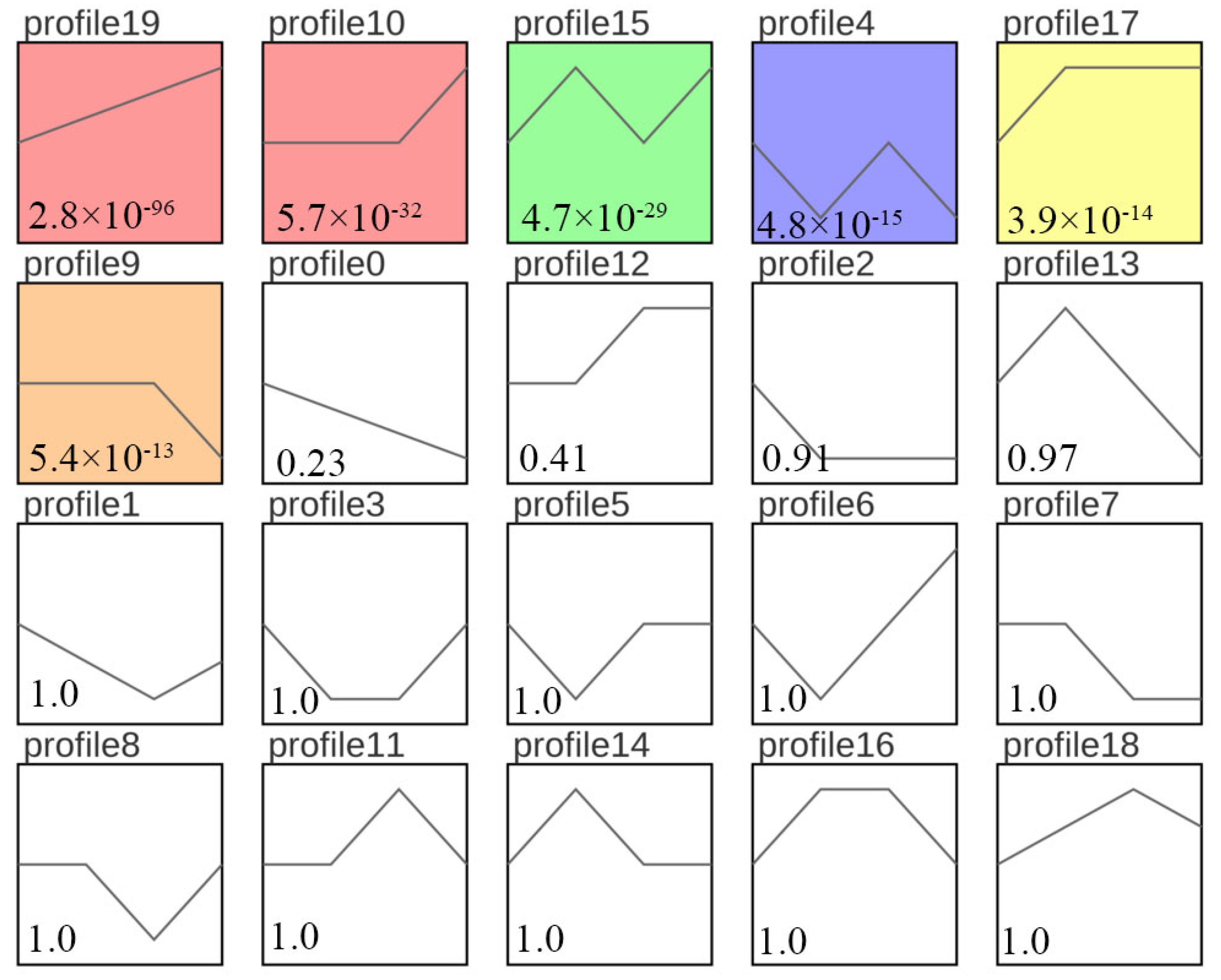
3.4. Sample Time-Series Analysis of DEGs
3.5. Construction of the Co-Expression Network (the WGCNA)
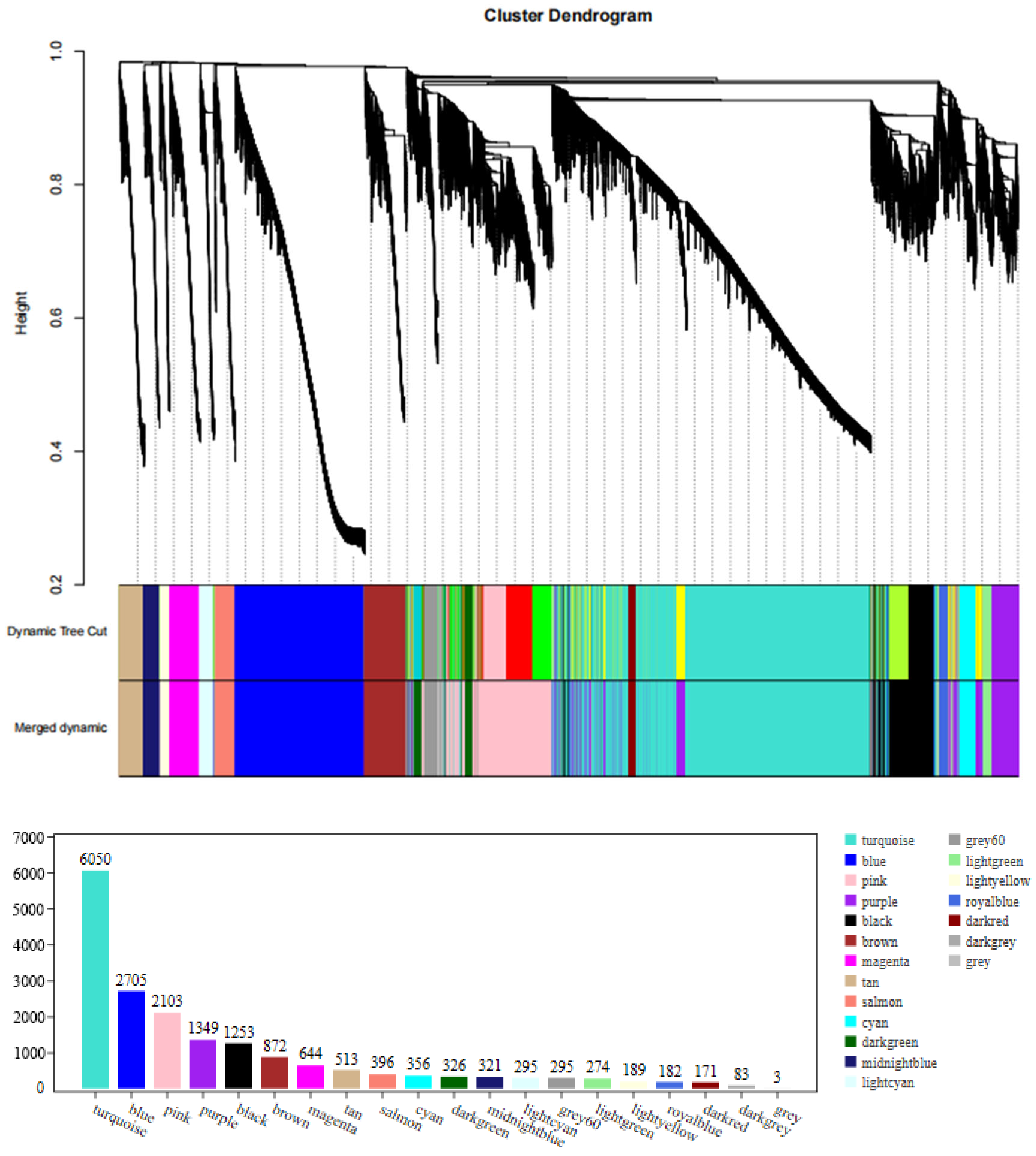
3.5.1. Clustering and Module Cutting of the Co-Expression Networks
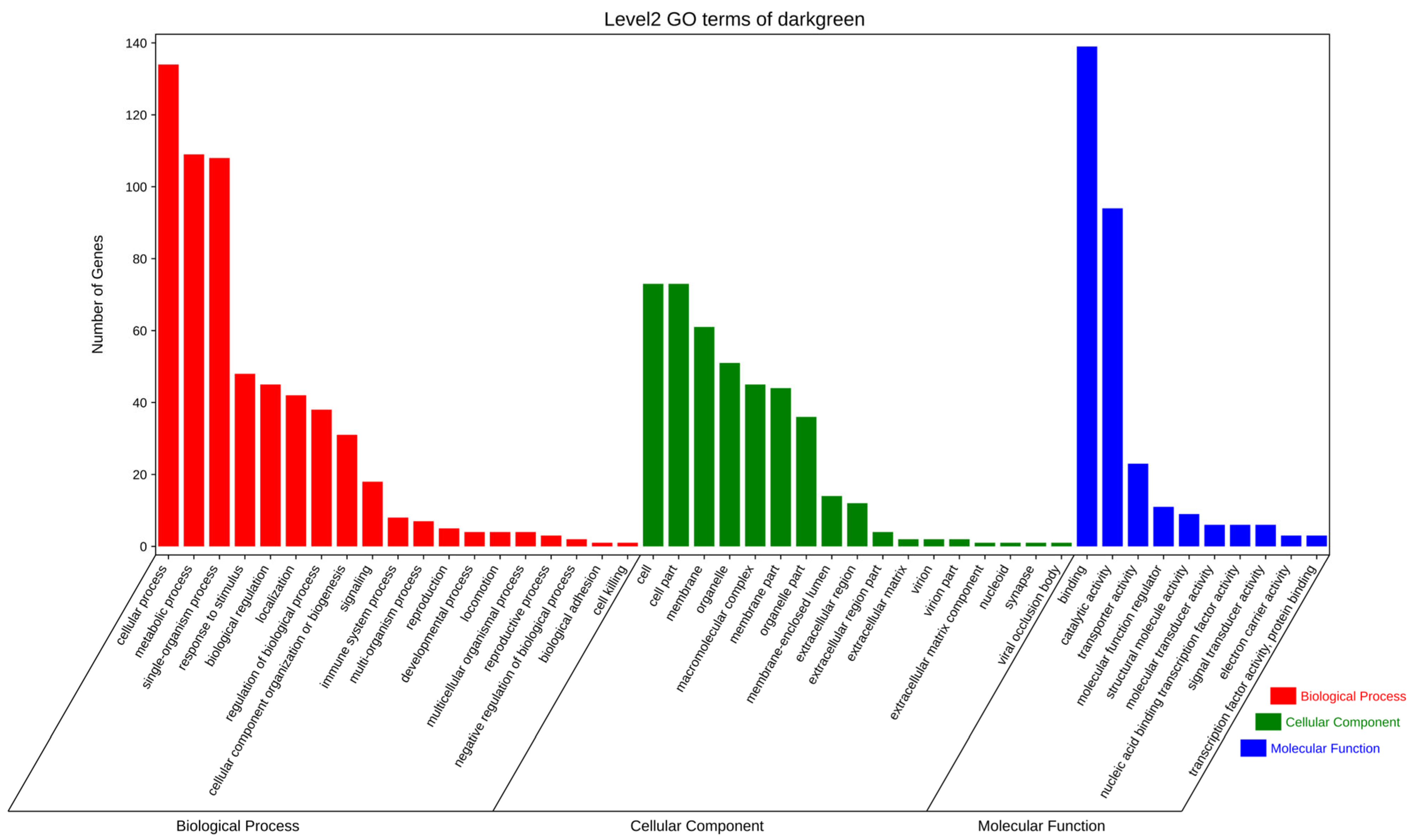
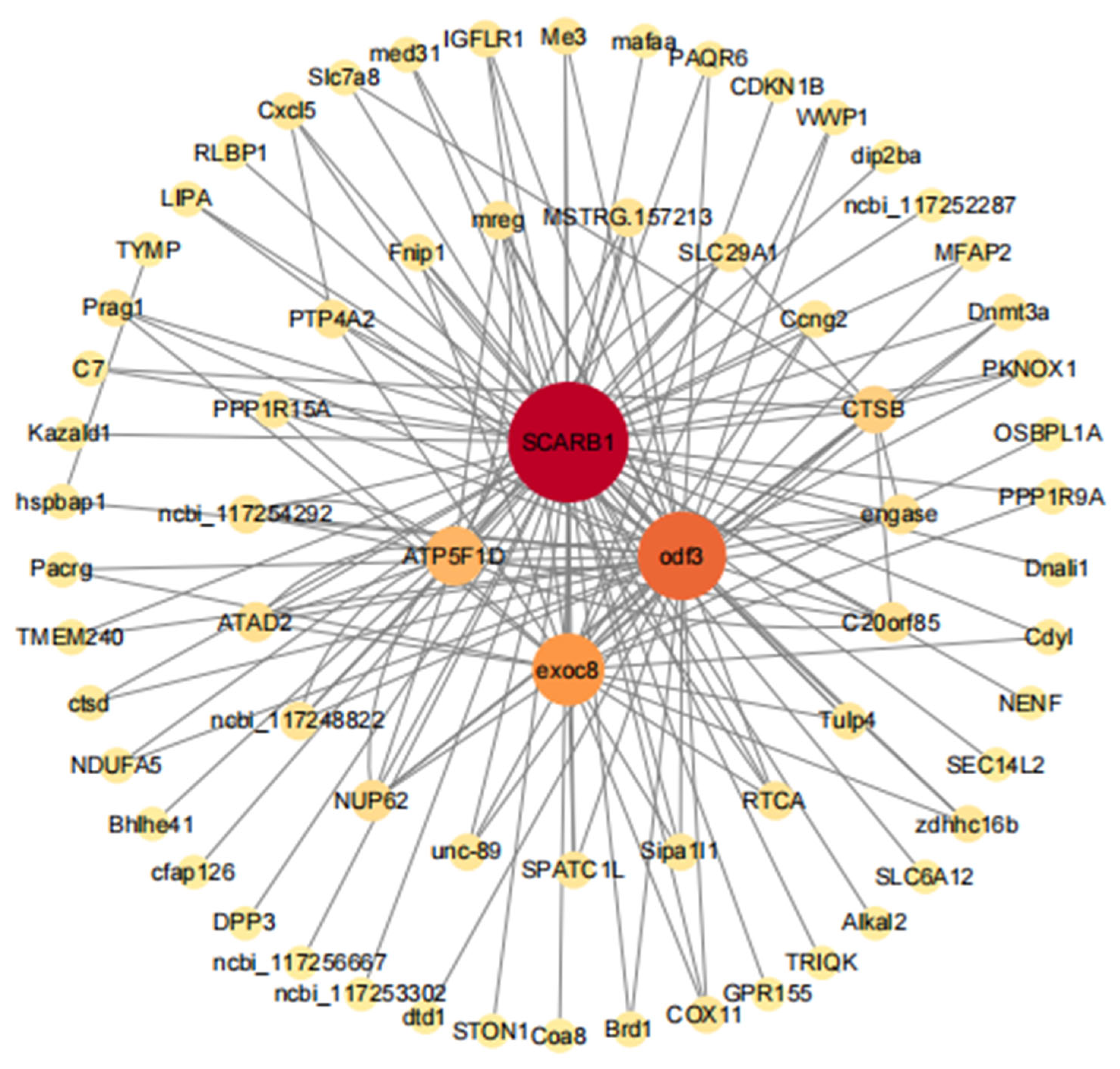
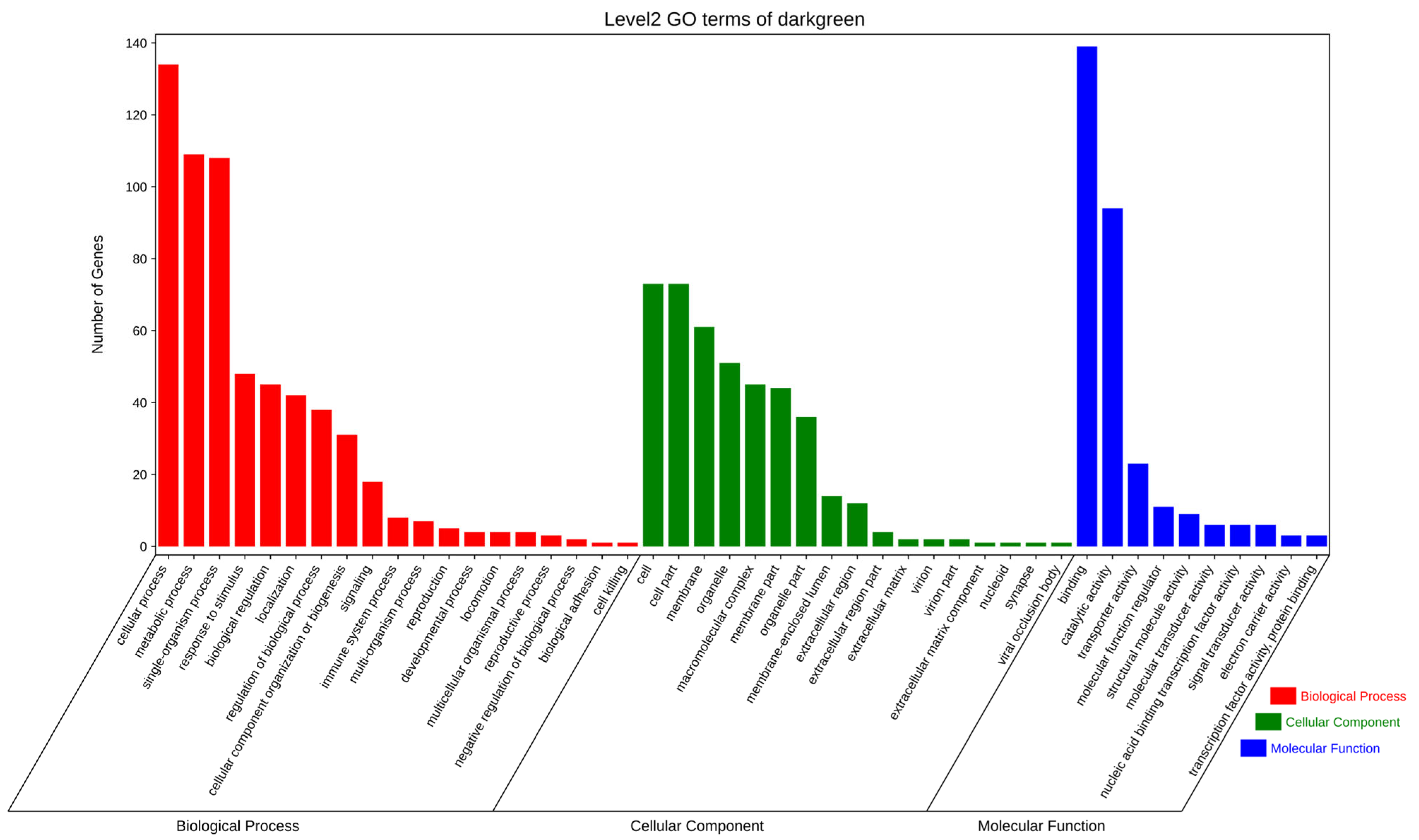
3.5.2. GO and KEGG Enrichment Analyses of Genes in the Dark-Green Module
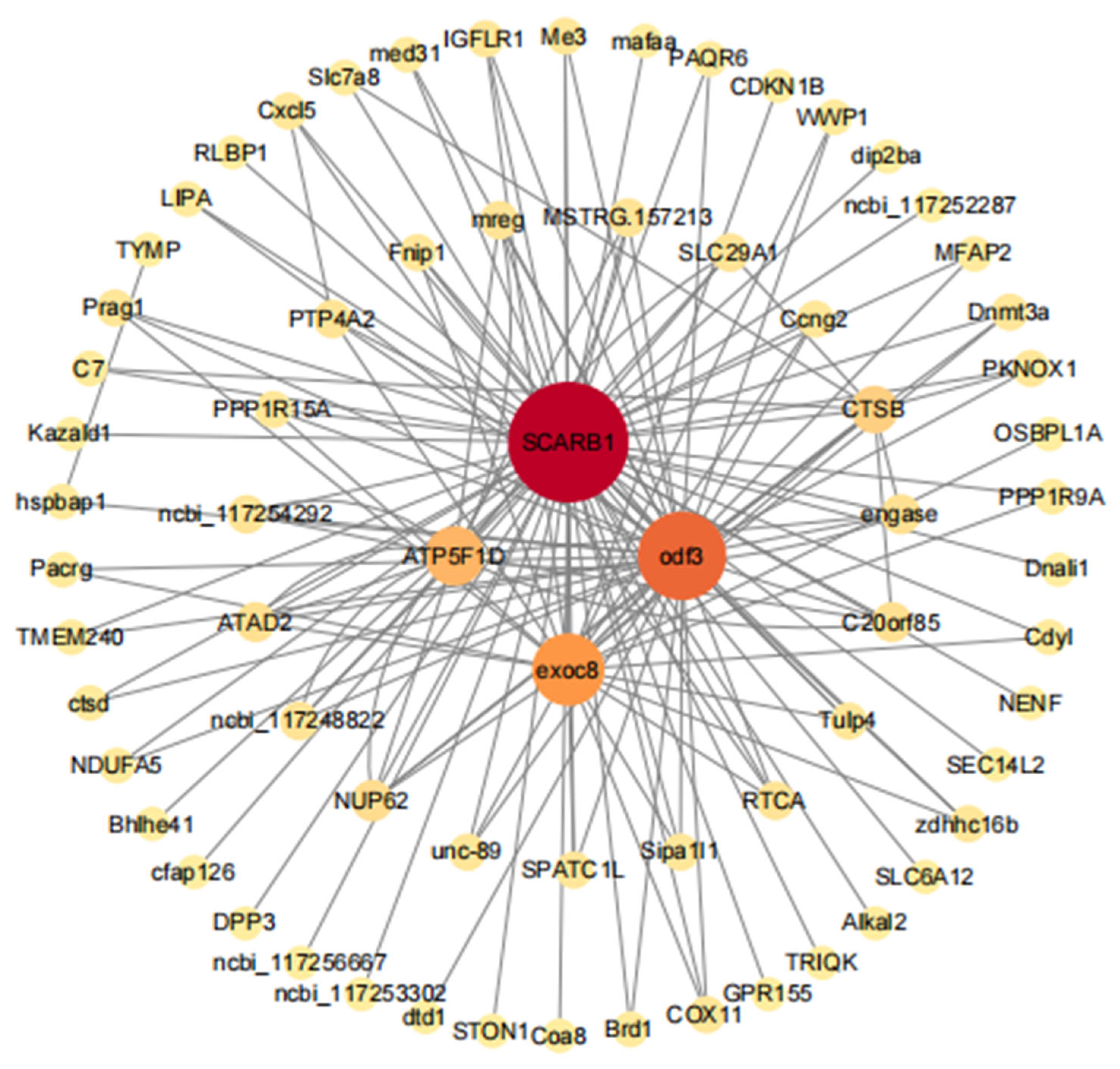
3.5.3. Gene Co-Expressed Network of the Dark-Green Module
3.6. Validation of Gene Expression via qRT-PCR
4. Discussion
4.1. Differential Expression between Fresh and Frozen Sperm of E. lanceolatus
4.2. Time-Series Expression Profiles and Co-Expression Network of Differentially Expressed Genes between Fresh Sperm and Frozen Sperm of E. lanceolatus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, Y.S.; Jiang, J.; Wang, N.; Qi, W.S.; Zhai, J.M.; Li, B.; Liang, Y.; Chen, Y.M.; Yang, C.J.; Chen, S.L. Sperm of the giant grouper: Cryopreservation, physiological and morphological analysis and application in hybridizations with red-spotted grouper. J. Reprod. Dev. 2015, 61, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.S.; Tang, J.; Ma, W.H.; Cheng, M.L.; Li, Z.T.; Wu, Y.P.; Zhang, J.J.; Wang, L.N.; Pang, Z.F.; Wang, X.M.; et al. Development and growth of hybrid offspring of brown grouper Epinephelus fuscoguttatus(♀) × blue speckled grouper Epinephelus tulcula(♂) using cryopreserved sperm. Adv. Fish. Sci. 2019, 40, 36–47. [Google Scholar] [CrossRef]
- Koh, I.C.C.; Yokoi, K.; Tsuji, M.; Tsuchihashi, Y.; Ohta, H. Cryopreservation of sperm from seven-band grouper, Epinephelus sptemfasciatus. Cryobiology 2010, 61, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.H.; Liao, I.C. Cryopreservation of finfish and shellfish gametes and embryos. In Reproductive Biotechnology in Finfish Aquaculture; Elsevier: Amsterdam, The Netherlands, 2001; pp. 161–189. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J.Y.; Gao, Z.H.; Wang, J.D.; Hao, H.R.; Song, J.T.; Wang, X.; Ma, Z.G.; Chen, Z.F.; Liu, S.J. Relationship between goose sperm migration rate and sperm motility parameters and their effects on reproductive efficiency of natural mated goose flocks. Chin. J. Anim. Sci. 2023, 59, 193–198. [Google Scholar] [CrossRef]
- Castro, L.S.; Hamilton, T.R.S.; Mendes, C.M.; Nichi, M.; Barnabe, V.H.; Visintin, J.A.; Assumpção, M.E.O.A. Sperm cryodamage occurs after rapid freezing phase:flow cytometry approach and antioxidant enzymes activity at different stages of cryopreservation. J. Anim. Sci. Biotechnol. 2016, 7, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.Y.; Li, Z.T.; Duan, P.F.; Qiu, Y.S.; Wang, X.Y.; Li, L.L.; Wang, L.N.; Liu, Y.; Li, W.S.; Wang, Q.B.; et al. Effect of long-term cryopreservation on ultrastructure and enzyme activity of Epinephelus lanceolatus sperm. J. Fish. China 2023, 47, 50–66. [Google Scholar] [CrossRef]
- Jung, Y.P.; Irfan, Z.; Yun, H.L.; Hyo, B.L.; Han, K.L. Effect of long-term storage on the quality of cryopreserved sperm of the giant grouper, Epinephelus lanceolatus. Aquaculture 2022, 555, 738154. [Google Scholar] [CrossRef]
- Yeste, M. Recent advances in boar sperm cryopreservation: State of the art and current perspectives. Reprod. Domest. Anim. 2015, 50, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.N.; Zhang, M.; Zhou, G.B.; Zhou, Y.M.; Wu, K.; Wang, C.D.; et al. Transcriptome sequencing reveals the differentially expressed lncRNAs and mRNAs involved in cryoinjuries in frozen-thawed giant panda (Ailuropoda melanoleuca) sperm. Int. J. Mol. Sci. 2018, 19, 3066. [Google Scholar] [CrossRef]
- Dai, D.H.; Qazi, I.H.; Ran, M.X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.B.; Angel, C.; Zeng, C.J. Exploration of miRNA and mRNA profiles in fresh and frozen-thawed boar sperm by transcriptome and small RNA sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gao, H.Y.; Zhang, Y.; Fan, G.Y.; Xu, H.; Zhai, J.M.; Xu, W.T.; Chen, Z.F.; Zhang, H.; Liu, S.S.; et al. A chromosome-level genome assembly of the giant grouper (Epinephelus lanceolatus) provides insights into its innate immunity and rapid growth. Mol. Ecol. Resour. 2019, 19, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.R.; Deng, Y.S.; Liang, C.G.; Geng, L.N.; Zhang, J.F.; Wu, X.Y.; Zhou, Z.Y.; Zhai, H.Y.; Cai, Q.X.; Zhang, L.; et al. Effects of feeding regimes and dietary energy on the production performance, plasma immune parameters and expression of genes involved in the TOR/S6K1 pathway of juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Aquaculture 2023, 571, 739457. [Google Scholar] [CrossRef]
- Contreras, P.; Dumorné, K.; Ulloa-Rodríguez, P.; Merino, O.; Figueroa, E.; Farías, J.G.; Valdebenito, I.; Risopatrón, J. Effects of short-term storage on sperm function in fish semen: A review. Rev. Aquac. 2020, 12, 1373–1389. [Google Scholar] [CrossRef]
- Cerolini, S.; Maldjian, A.; Pizzi, F.; Gliozzi, T.M. Changes in sperm quality and lipid composition during cryopreservation of boar semen. Reproduction 2001, 121, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Paasch, U.; Sharma, R.K.; Gupta, A.K.; Grunewald, S.; Mascha, E.J.; Thomas, A.J.; Glander, H.J.; Agarwal, A. Cryopreservation and thawing is associated with varying extent of activation of apoptotic machinery in subsets of ejaculated human spermatozoa. Biol. Reprod. 2004, 71, 1828–1837. [Google Scholar] [CrossRef]
- Boerke, A.; Dieleman, S.J.; Gadella, B.M. A possible role for sperm RNA in early embryo development. Theriogenology 2007, 68, S147–S155. [Google Scholar] [CrossRef]
- Braun, R.E. Packaging paternal chromosomes with protamine. Nat. Genet. 2001, 28, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Mulligan, B.P.; Kim, H.; Yang, B.; Lee, C. Quantitative analysis of sperm mRNA in the pig: Relationship with early embryo development and capacitation. Reprod. Fertil. Dev. 2012, 25, 807–817. [Google Scholar] [CrossRef]
- Liang, J.C.; Lü, C.R.; Hong, Q.H.; Wu, G.Q.; Quan, G.B. Analysis of sperm differential transcriptome expression before and after freezing in yunnan semi-fine wool sheep. China Anim. Husb. Vet. Med. 2022, 49, 973–981. [Google Scholar] [CrossRef]
- Papas, M.; Catalán, J.; Fernandez-Fuertes, B.; Arroyo, L.; Bassols, A.; Miró, J.; Yeste, M. Specific activity of superoxide dismutase in stallion seminal plasma is related to sperm cryotolerance. Antioxidants 2019, 8, 539. [Google Scholar] [CrossRef]
- Wang, Y.B.; Wang, Y.F.; Liu, X.; Zhou, J.Q.; Deng, H.B.; Zhang, G.L.; Xiao, Y.H.; Tang, W.B. WGCNA Analysis Identifies the Hub Genes Related to Heat Stress in Seedling of Rice (Oryza sativa L.). Genes 2022, 13, 1020. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef]
- Zheng, W.L. Study on the Mechanism of MAPK Signaling pathway in the Infection of Sarcocystis cruzi in Yak. Master’s Thesis, Qinghai University, Xining, China, 2022. [Google Scholar] [CrossRef]
- Zeng, L.; Wang, Y.H.; Song, W.; Xie, Z.L.; Zhang, H. Effects of Cu acclimation on oxidative damage in the large yellow croaker under cold stress based on transcriptome analysis. J. Fish. Sci. China 2022, 29, 1425–1436. [Google Scholar]
- Shang, Y.T.; Luo, X.B.; Zhang, H.; Chen, M.J.; Yin, W.; Cao, Z.J.; Deng, R.J.; Li, Y.; Li, F. Genome-Wide Identification and Analysis of the MAPK and MAPKK Gene Families in Potato (Solanum tuberosum L.). Agronomy 2023, 13, 93. [Google Scholar] [CrossRef]
- Miao, B.B.; Niu, S.F.; Wu, R.X.; Liang, Z.B.; Tang, B.G.; Zhai, Y.; Xu, X.Q. Gene Expression Profile and Co-Expression Network of Pearl Gentian Grouper under Cold Stress by Integrating Illumina and PacBio Sequences. Animals 2021, 11, 1745. [Google Scholar] [CrossRef]
- Yu, L.S.; Yang, X.J.; Ma, B.; Ying, H.J.; Shang, X.J.; He, B.F.; Zhang, Q. Abnormal arachidonic acid metabolic network may reduce sperm motility via P38 MAPK. Open Biol. 2019, 9, 180091. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.K.; Wang, G.; Wang, X.L. Research progress of heat stress on gene hsp70 and caspase-3 infish. Hebei Fish. 2022, 338, 28–32. [Google Scholar]
- He, X.Y.; Lin, Z.M.; Ning, J.Y.; Li, N.; Cui, X.L.; Zhao, B.X.; Hong, F.Z.; Miao, J.Y. Promoting TTC4 and HSP70 interaction and translocation of annexin A7 to lysosome inhibits apoptosis in vascular endothelial cells. FASEB J. 2020, 34, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, A.F.; Costantino, V.; Appiolaza, M.L.; Cacciamani, V.; Benardon, M.E.; Bocanegra, M.G.; Vallés, P. Heat shock protein 70 and CHIP promote Nox4 ubiquitination and degradation within the losartan antioxidative effect in proximal tubule cells. Cell. Physiol. Biochem. 2015, 36, 2183–2197. [Google Scholar] [CrossRef] [PubMed]
- Richter-Landberg, C.; Goldbaum, O. Stress proteins in neural cells:functional roles in health and disease. Cell. Mol. Life Sci. CMLS 2003, 60, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Bin, S.Y.; Zhong, D.D.; Du, X.S.; Zhang, Y.D.; Lin, Y.; Huang, Y.; Wen, L.T. Correlation analysis of polymorphism and its cold tolerance traits HSP70 gene of GIFT tilapia (Oreochromis niloticus). J. Guangxi Norm. Univ. (Nat. Sci. Ed.) 2019, 37, 211–217. [Google Scholar] [CrossRef]
- Helsen, C.; Kerkhofs, S.; Clinckemalie, L.; Spans, L.; Laurent, M.; Boonen, S.; Vanderschueren, D.; Claessens, F. Structural basis for nuclear hormone receptor DNA binding. Mol. Cell. Endocrinol. 2012, 348, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Häcker, H.; Tseng, P.H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef]
- Place, S.P.; Hofmann, G.E. Constitutive expression of a stress-inducible heat shock protein gene, hsp70, in phylogenetically distant Antarctic fish. Polar Biol. 2005, 28, 261–267. [Google Scholar] [CrossRef]
- Ren, K.X.; Li, S.Z.; Liang, S.Q.; Fan, F.; Lu, J.R.; Wei, T.X.; Cao, X.L.; Gong, L.; Li, H.Z.; Zhao, J.L.; et al. Notch signaling dependent monocyte conversion alleviates immune-mediated neuropathies by regulating RBP-J/NR4A1 axis. J. Autoimmun. 2022, 133, 102945. [Google Scholar] [CrossRef]
- Yang, E.J.; Yang, L.T.; Wang, W.Z.; Huang, J.S.; Zhang, J.D.; Wang, Z.L.; Chen, G. Identification and functional analysis of SNP from transcriptome of cobia (Rachycentron canadum) in response to hypoxia stress. Haiyang Xuebao 2022, 44, 113–124. [Google Scholar]
- Li, X.H. Role of Megalobrama Amblycephala bcl2l13 in Cell Apoptosis. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2022. [Google Scholar] [CrossRef]
- D’arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.F.; Sun, B.B.; Ke, W.J.; Yang, Y.; Yu, H.; Chen, Y.F. Advances in the study of pathogen-induced apoptosis pathways in aquatic animals. Fish. Sci. Technol. Inf. 2022, 49, 164–170. [Google Scholar] [CrossRef]
- Li, Y.; Huang, J.S.; Chen, Y.M.; Wen, Z.W.; Ou, G.H.; Huang, J.P.; Jiang, X.T.; Xie, R.T.; Ma, Q.; Chen, G. Effect of low temperature stress on antioxidant stress, apoptosis and histological structure of gills in cobia (Rachycentron canadum). S. China Fish. Sci. 2023, 19, 68–77. [Google Scholar] [CrossRef]
- Wang, H.L.; Montague, H.R.; Hess, N.H.; Zhang, Y.; Aguilar, G.L.; Dunham, R.A.; Butts, I.A.E.; Wang, X. Transcriptome analysis reveals key gene expression changes in Blue Catfish sperm in response to cryopreservation. Int. J. Mol. Sci. 2022, 23, 7618. [Google Scholar] [CrossRef]
- Yu, Z.X.; Ren, X.Y.; Shao, H.X.; Liu, P.; Li, J. Effect of low temperature stress on antioxidant system and apoptosis of Marsupenaeus japonicus. Prog. Fish. Sci. 2022, 43, 157–166. [Google Scholar]
- Peng, Y.; Akmentin, W.; Connelly, M.A.; Lund-Katz, S.; Phillips, M.C.; Williams, D.L. Scavenger receptor BI (SR-BI) clustered on microvillar extensions suggests that this plasma membrane domain is a way station for cholesterol trafficking between cells and high-density lipoprotein. Mol. Biol. Cell 2004, 15, 384–396. [Google Scholar] [CrossRef]
- Sèdes, L.; Thirouard, L.; Maqdasy, S.; Garcia, M.; Caira, F.; Lobaccaro, G.M.A.; Beaudoin, C.; Volle, D.H. Cholesterol: A gatekeeper of male fertility? Front. Endocrinol. 2018, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Sun, J.J.; Liu, Y.; Zheng, L.; Zhang, X.; Yang, H. Effect of miR-125a-5p targeting the Scarb1 gene on hypoxic/reoxygenation injury of cardiomyocyte and its mechanism. Chin. J. Med. Genet. 2020, 37, 980–986. [Google Scholar] [CrossRef]
- Zhao, W.L.; Li, Z.Z.; Ping, P.; Wang, G.S.; Yuan, X.B.; Sun, F. Outer dense fibers stabilize the axoneme to maintain sperm motility. J. Cell. Mol. Med. 2018, 22, 1755–1768. [Google Scholar] [CrossRef]
- Lü, Z.L.; Shan, X.D.; Du, J.; Zhu, S.M.; Zhuang, J.; Liang, X.; Zhao, Q.M. Impacts of ODF2 gene knockdown on the sperm motility and fertility of male mice. Natl. J. Androl. Zhonghua Nan Ke Xue Za Zhi 2022, 28, 867–872. [Google Scholar] [CrossRef]
- Coulter, M.E.; Musaev, D.; DeGennaro, E.M.; Zhang, X.C.; Henke, K.; James, K.N.; Smith, R.S.; Hill, R.S.; Partlow, J.N.; Saffar, M.A.; et al. Regulation of human cerebral cortical development by EXOC7 and EXOC8, components of the exocyst complex, and roles in neural progenitor cell proliferation and survival. Genet. Med. Off. J. Am. Coll. Med. Genet. 2020, 22, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Krishin, J.; Haider, N.; Aurangzeb, B.; Abdullah; Suleman, S.; Ahmad, W.; Hansen, T.; Basit, S. A novel nonsense variant in EXOC8 underlies a neurodevelopmental disorder. Neurogenetics 2022, 23, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J. Embryo Cryopreservation and Transcriptome Analysis of Surviving Larvae in Kelp Grouper (Epinephelus moara). Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2020. [Google Scholar] [CrossRef]
- Zhao, H.J.; Khan, Z.; Westlake, C.J. Ciliogenesis membrane dynamics and organization. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2023; Volume 133, pp. 20–31. [Google Scholar] [CrossRef]
- Torra-Massana, M.; Jodar, M.; Barragán, M.; Soler-Ventura, A.; Delgado-Dueñas, D.; Rodríguez, A.; Oliva, R.; Vassena, R. Altered mitochondrial function in spermatozoa from patients with repetitive fertilization failure after ICSI revealed by proteomics. Andrology 2021, 9, 1192–1204. [Google Scholar] [CrossRef]
- Zhou, Z. ATPAF1 Is Essential for the Mouse Heart to Maintain Structure and Function via Controlling ATP Synthase Assembly and Mitochondrial Respiration. Master’s Thesis, Huazhong University of Science and Technology, Huazhong, China, 2020. [Google Scholar] [CrossRef]
- Hsieh, R.H.; Au, H.K.; Yeh, T.S.; Chang, S.J.; Cheng, Y.F.; Tzeng, C.R. Decreased expression of mitochondrial genes in human unfertilized oocytes and arrested embryos. Fertil. Steril. 2004, 81, 912–918. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Forward Primer | Reverse Primer |
|---|---|---|
| ids | GGATGGTAAACTCCACGCCA | ACATCATTGTTGGCCCGACT |
| junb | TTCTACAACCGGGGCATCAC | GTAACTCCACCGGCTCCAAG |
| plekho2 | ACTCGATCAAGGCCCAAAGG | TCCAAGCGCAGGATACCATC |
| hsp70 | TCAATGACTCCCAGCGACAG | TTGTCCAGGCCGTATGCAAT |
| Ccdc103 | CGAGGTTGGAGCGTGATGTC | TTTCGCTTCGTTCTCTCGCT |
| ksr2 | TCCCTCAAAGATCACCAAGGA | CGTCTGACTGATGTGCAGGT |
| sema5b | TCGTGCTGATCATCTGTTCGT | TGGGAGAACTCCGACATCCA |
| jun | AGAAAGCGGATGAGGAACCG | GACGCGAGCTCCGAATTTTG |
| zfp36 | GCAGTAAGTGCCAGTTTGCC | CGTAGGGGCAGTAGCCAAAG |
| d10wsu102e | AGCCAAAGACTGGCAGATCC | GCTGCTGAAGCTTCTCGTTG |
| mcl1 | CGAAGGACTCTCACAACGGG | CGGAGGTTCTTGGTCGCATA |
| lamb1 | AACCCCAAGCACTCTTACGG | GTCGCATGCGTGACATTTGA |
| Zfand4 | TCGCCTCTTTCGTTCACTCC | GAAGGTGGGGAGGGGTCTAT |
| igll5 | GCAGTGGGATCTCTACCAGC | CTGGGAGCCCAAAGACACTT |
| β-actin | CTCTGGGCAACGGAACCTCT | GTGCGTGACATCAAGGAGAAGC |
| Sample | Raw Data (bp) | Clean Data (bp) | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|
| El-61-1 | 11,503,797,600 | 11,260,881,272 | 97.19% | 93.07% | 52.88% |
| El-61-2 | 12,572,943,600 | 12,403,629,774 | 97.01% | 92.46% | 53.00% |
| El-61-3 | 11,964,763,200 | 11,764,204,113 | 97.16% | 92.77% | 52.10% |
| El-49-1 | 11,607,607,500 | 11,310,292,815 | 97.22% | 92.95% | 52.59% |
| El-49-2 | 12,134,965,800 | 11,901,108,789 | 97.20% | 92.91% | 53.89% |
| El-49-3 | 11,793,498,600 | 11,588,109,505 | 97.12% | 92.74% | 54.43% |
| El-23-1 | 11,899,454,100 | 11,501,842,347 | 96.70% | 92.33% | 61.06% |
| El-23-2 | 11,563,097,700 | 11,268,692,649 | 96.94% | 92.31% | 55.27% |
| El-23-3 | 12,992,697,600 | 12,752,937,675 | 96.86% | 92.08% | 55.99% |
| El-0-1 | 12,138,655,500 | 11,919,323,093 | 97.47% | 93.30% | 52.50% |
| El-0-2 | 12,979,071,300 | 12,533,244,899 | 96.99% | 92.49% | 52.80% |
| El-0-3 | 11,868,128,400 | 11,614,133,308 | 96.89% | 92.13% | 53.61% |
| Sample | Total | Unique_Mapped (%) | Multiple_Mapped (%) | Total_Mapped (%) |
|---|---|---|---|---|
| El-61-1 | 60,355,436 | 36,288,656 (60.12%) | 5,790,104 (9.59%) | 42,078,760 (69.72%) |
| El-61-2 | 65,623,376 | 38,386,462 (58.50%) | 6,137,785 (9.35%) | 44,524,247 (67.85%) |
| El-61-3 | 63,992,594 | 38,520,927 (60.20%) | 6,298,505 (9.84%) | 44,819,432 (70.04%) |
| El-49-1 | 51,620,698 | 29,550,592 (57.25%) | 3,952,694 (7.66%) | 33,503,286 (64.90%) |
| El-49-2 | 52,834,236 | 29,249,147 (55.36%) | 4,232,201 (8.01%) | 33,481,348 (63.37%) |
| El-49-3 | 48,739,846 | 27,147,933 (55.70%) | 3,759,760 (7.71%) | 30,907,693 (63.41%) |
| El-23-1 | 47,965,958 | 20,982,137 (43.74%) | 3,701,637 (7.72%) | 24,683,774 (51.46%) |
| El-23-2 | 52,571,466 | 26,721,876 (50.83%) | 4,236,098 (8.06%) | 30,957,974 (58.89%) |
| El-23-3 | 59,635,270 | 31,264,050 (52.43%) | 4,801,141 (8.05%) | 36,065,191 (60.48%) |
| El-0-1 | 63,585,244 | 35,390,829 (55.66%) | 4,999,935 (7.86%) | 40,390,764 (63.52%) |
| El-0-2 | 63,890,574 | 35,368,631 (55.36%) | 4,969,086 (7.78%) | 40,337,717 (63.14%) |
| El-0-3 | 59,705,142 | 32,478,564 (54.40%) | 4,660,800 (7.81%) | 37,139,364 (62.20%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Tian, Y.; Qiu, Y.; Duan, P.; Wang, X.; Li, Z.; Li, L.; Liu, Y.; Wang, L. Effects of Long-Term Cryopreservation on the Transcriptomes of Giant Grouper Sperm. Genes 2024, 15, 523. https://doi.org/10.3390/genes15040523
Ding X, Tian Y, Qiu Y, Duan P, Wang X, Li Z, Li L, Liu Y, Wang L. Effects of Long-Term Cryopreservation on the Transcriptomes of Giant Grouper Sperm. Genes. 2024; 15(4):523. https://doi.org/10.3390/genes15040523
Chicago/Turabian StyleDing, Xiaoyu, Yongsheng Tian, Yishu Qiu, Pengfei Duan, Xinyi Wang, Zhentong Li, Linlin Li, Yang Liu, and Linna Wang. 2024. "Effects of Long-Term Cryopreservation on the Transcriptomes of Giant Grouper Sperm" Genes 15, no. 4: 523. https://doi.org/10.3390/genes15040523
APA StyleDing, X., Tian, Y., Qiu, Y., Duan, P., Wang, X., Li, Z., Li, L., Liu, Y., & Wang, L. (2024). Effects of Long-Term Cryopreservation on the Transcriptomes of Giant Grouper Sperm. Genes, 15(4), 523. https://doi.org/10.3390/genes15040523