

Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Experiments
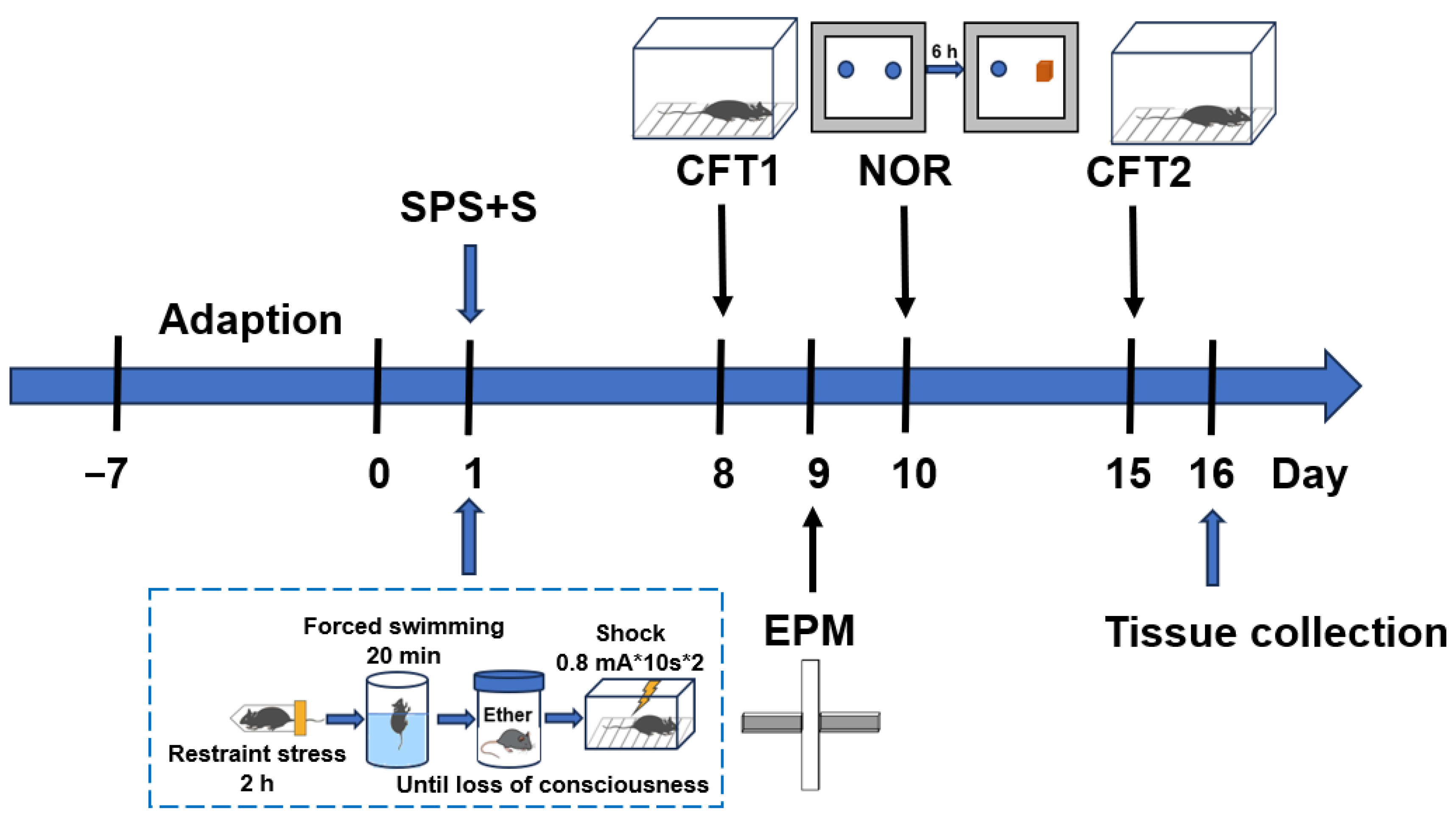
2.2.1. SPS+S Model
2.2.2. Contextual Freezing Test (CFT)
2.2.3. Elevated plus Maze (EPM) Test
2.2.4. Novel Object Recognition (NOR) Test
2.3. Drug Administration
2.4. SnRNA-seq and Analysis
2.4.1. Tissue Dissociation for 10× Genomics
2.4.2. Quality Control and Cell Type Identification
2.4.3. SnRNA-seq Data Analysis
3. Statistical Analysis
4. Results
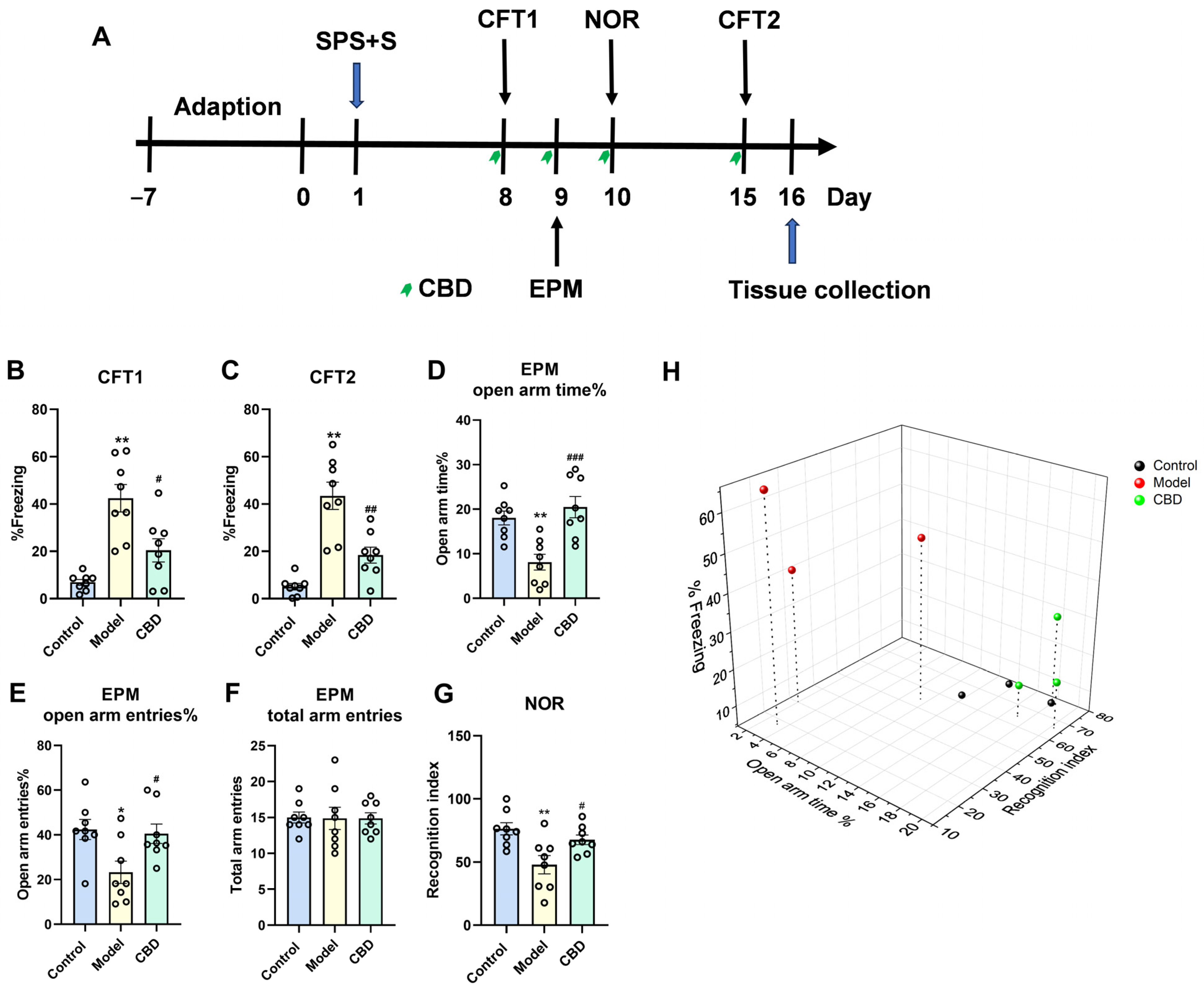
4.1. CBD Mitigated All PTSD-like Symptoms in the SPS+S Mice Model
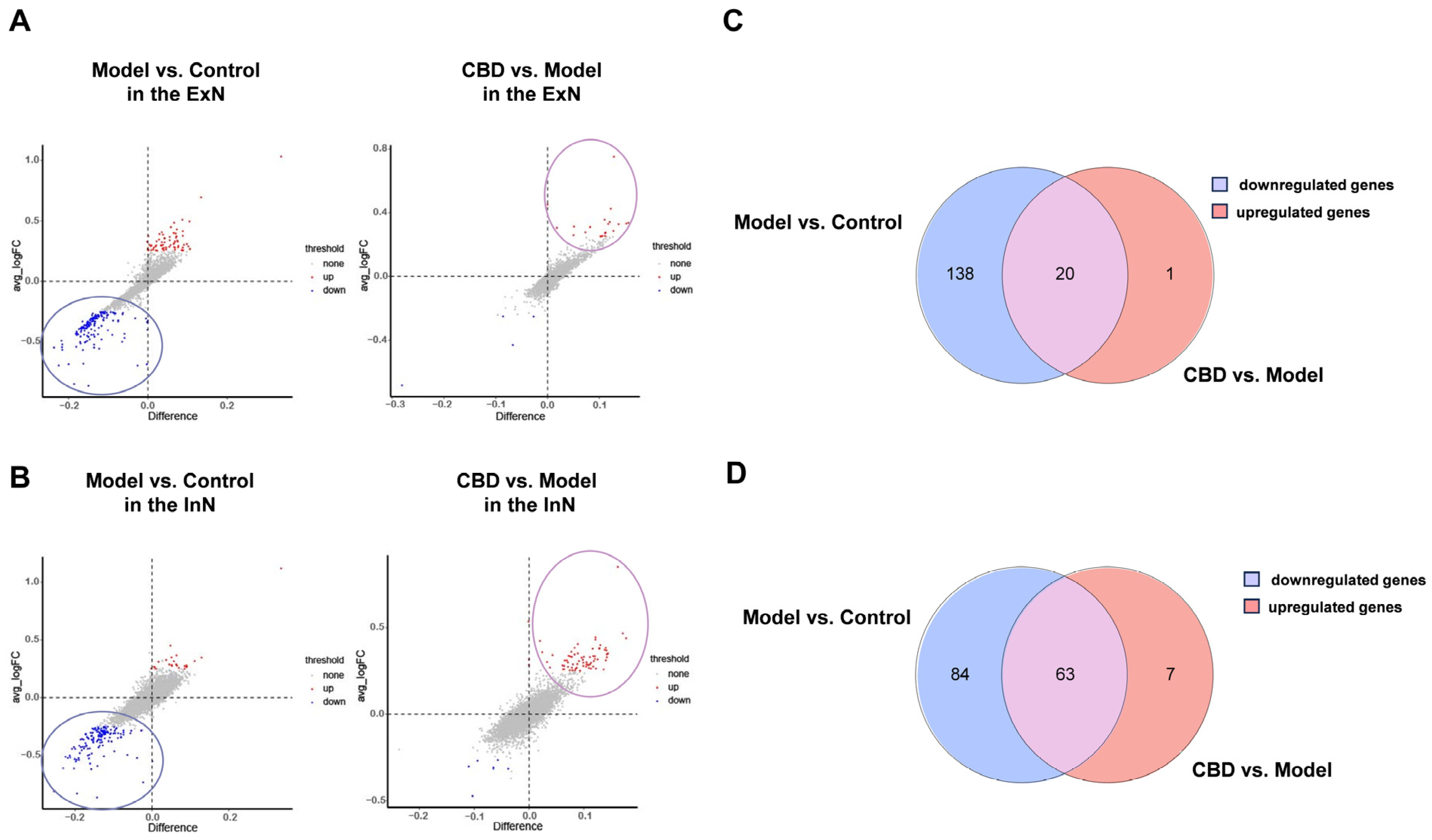
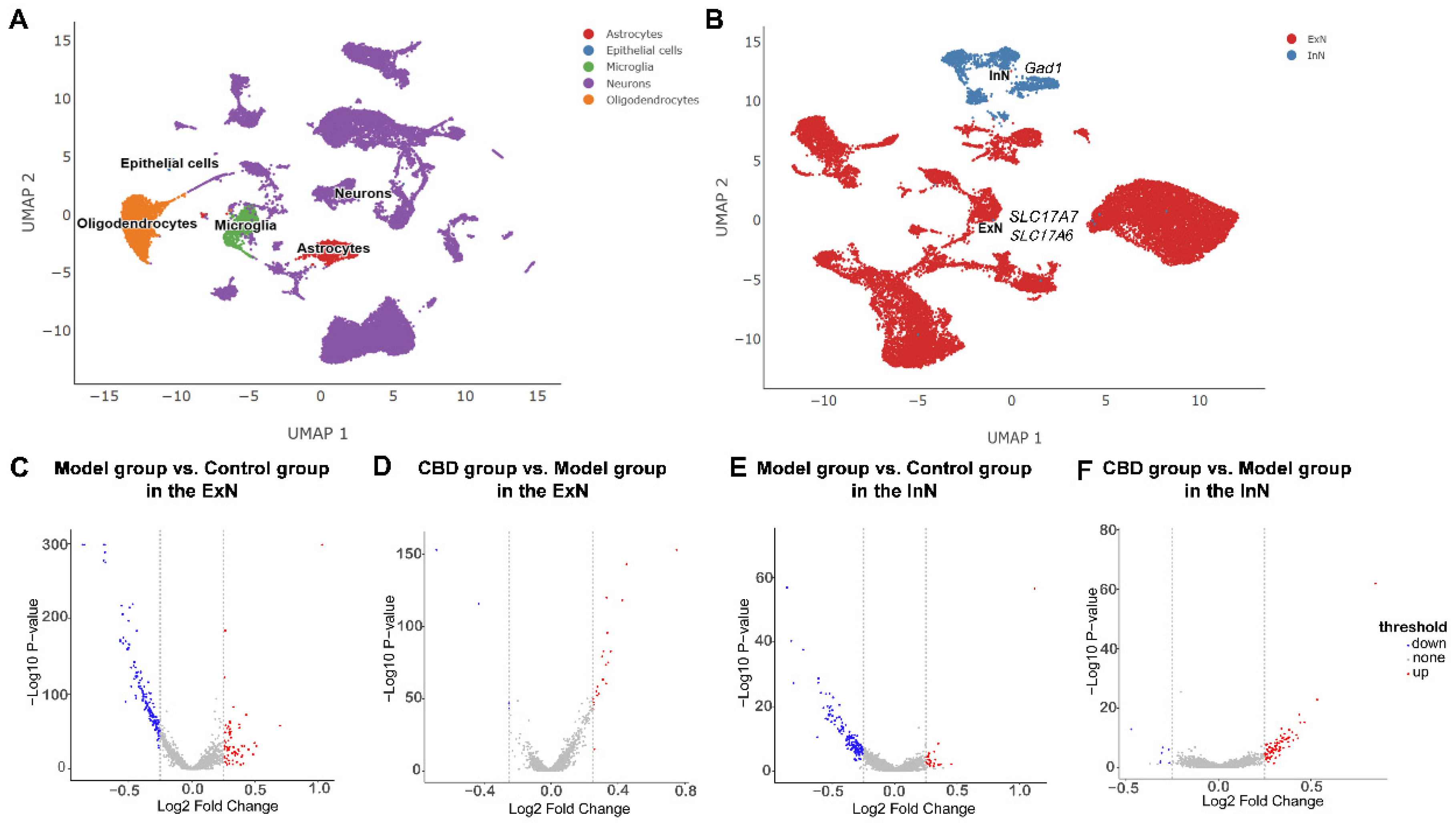
4.2. Analysis of Differentially Expressed Genes in the PTSD Mice
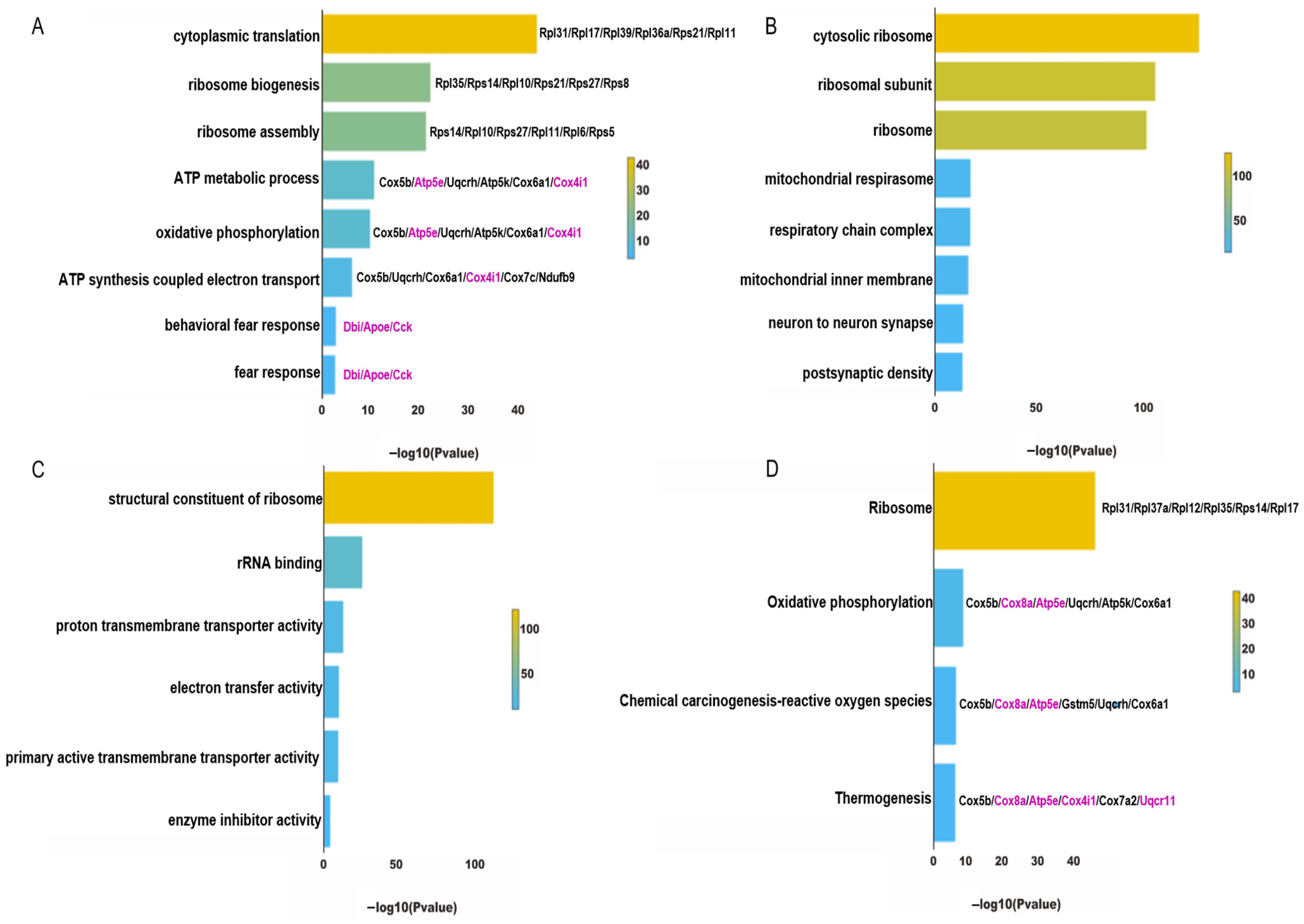
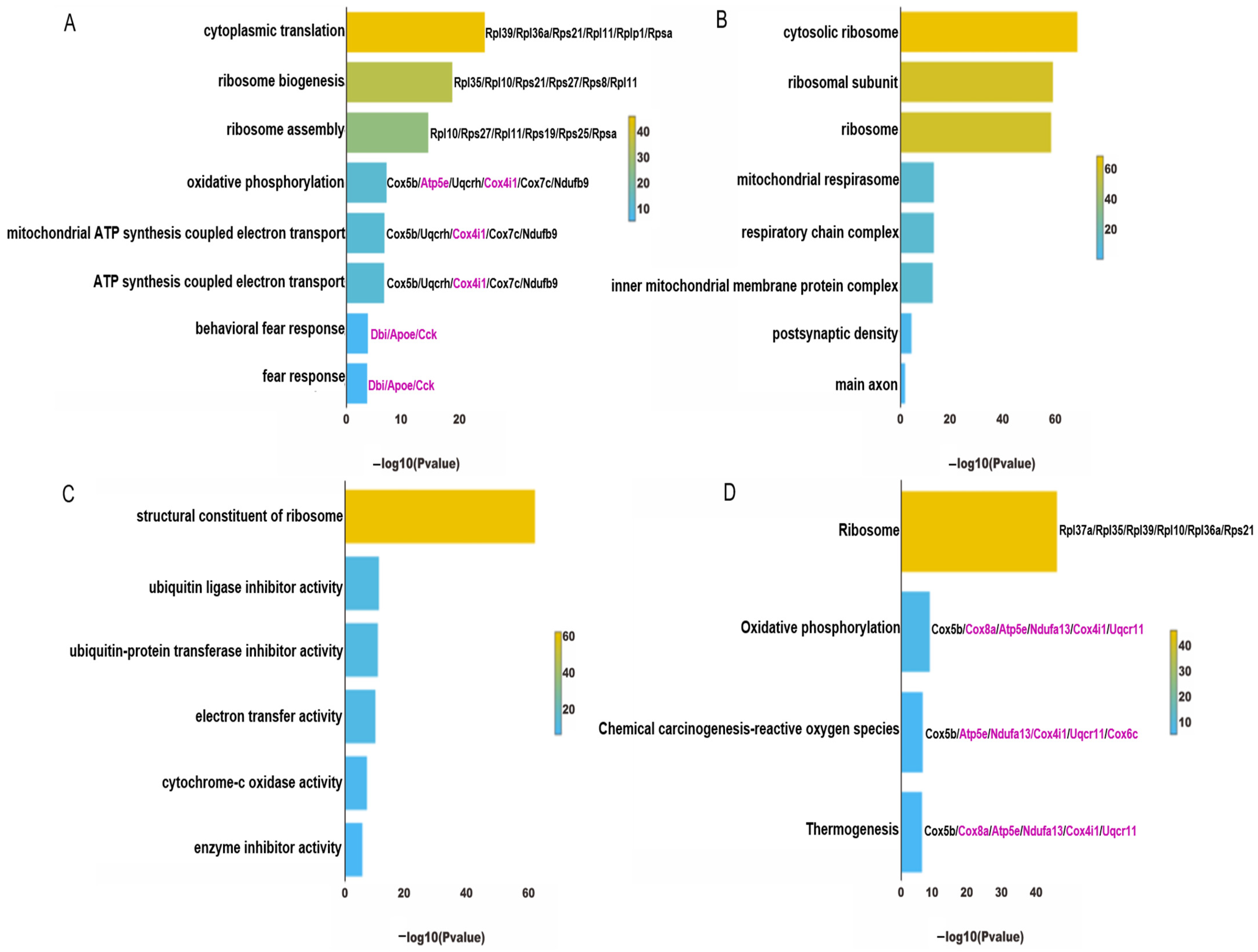
4.3. Functional Enrichment Analysis of ExNs in PTSD Mice
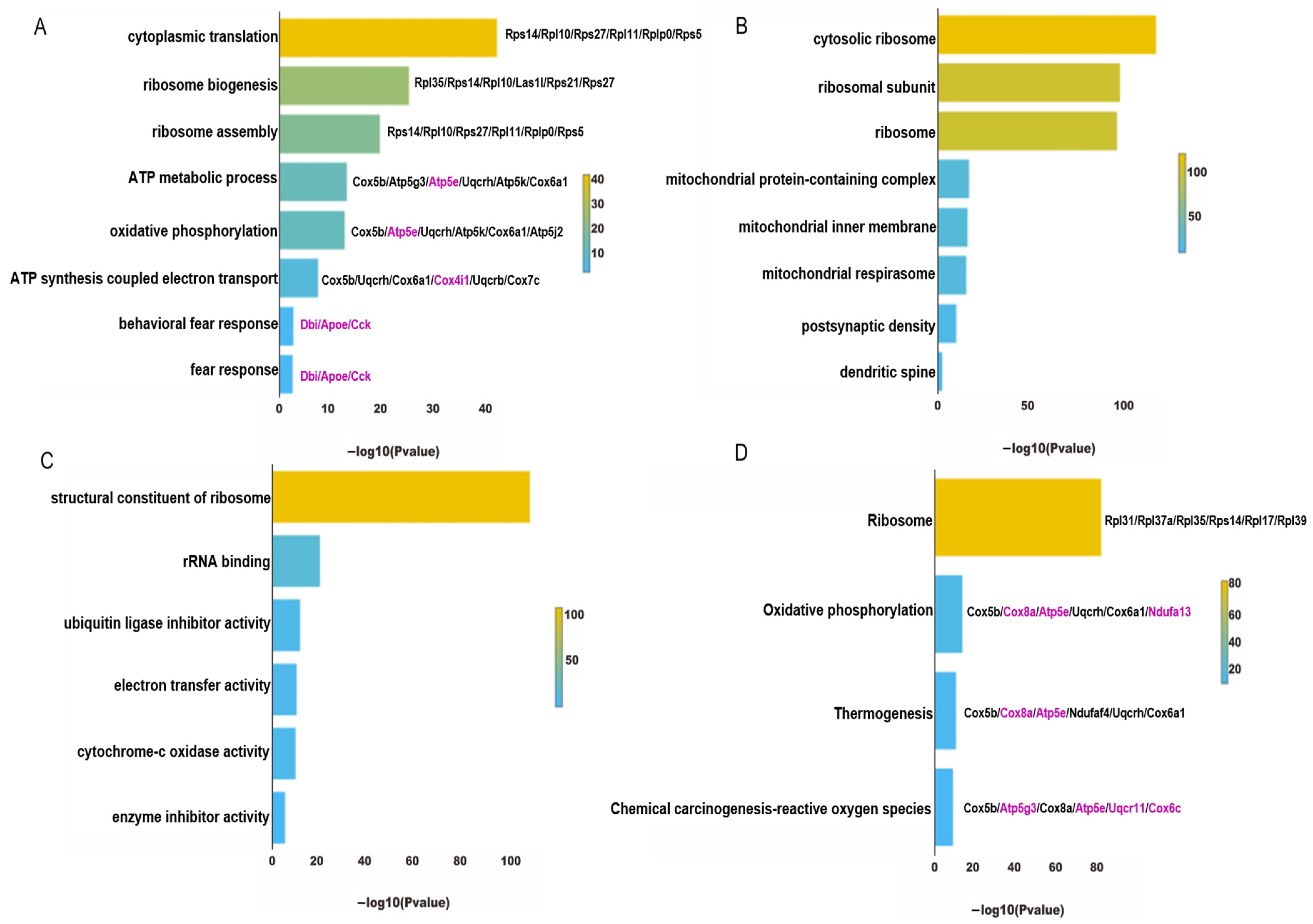
4.4. Functional Enrichment Analysis of InNs in PTSD Mice
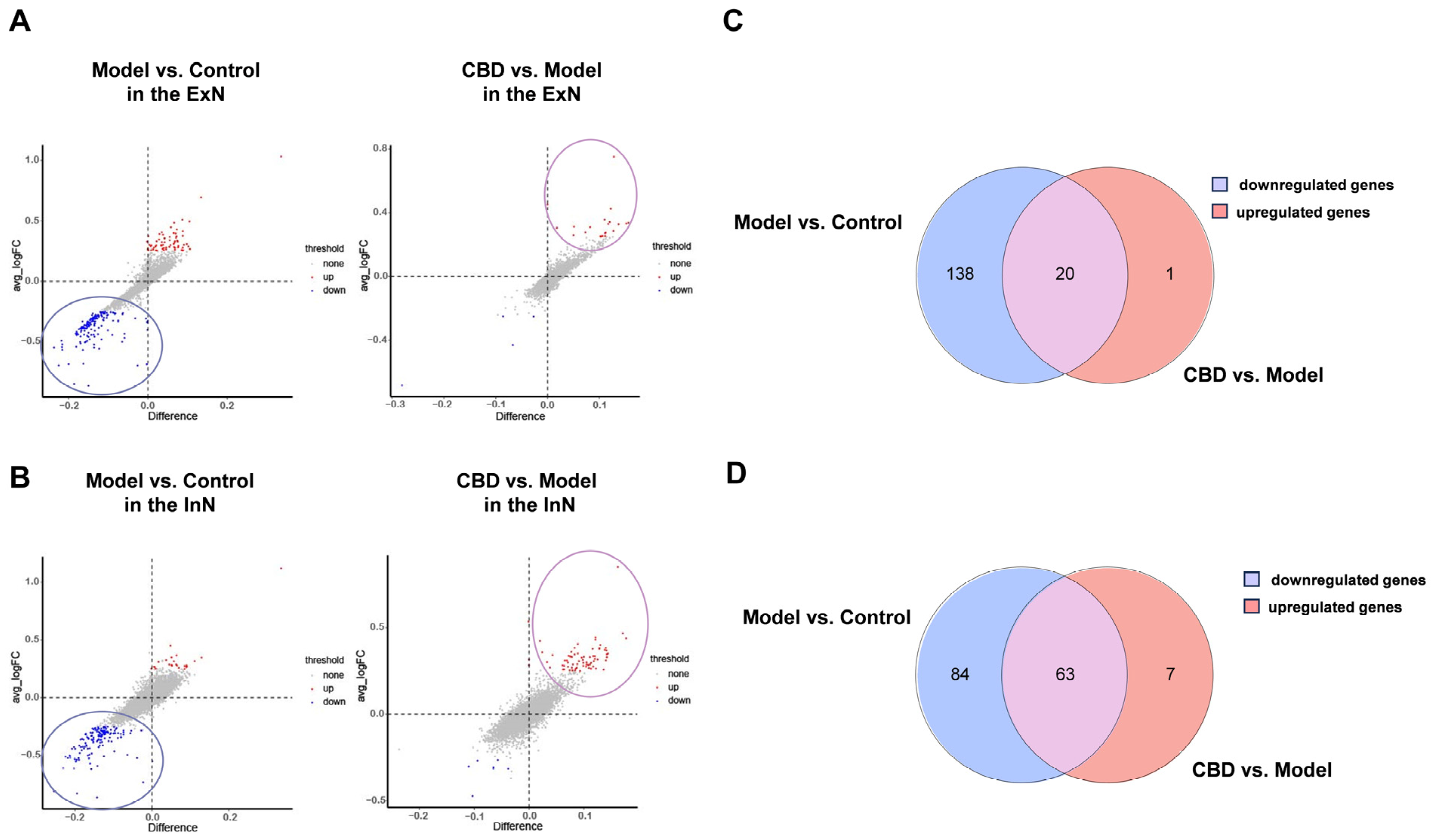
4.5. Identification and Functional Enrichment Analysis of Core Genes from ExNs and InNs Involved in CBD’s Anti-PTSD Effect
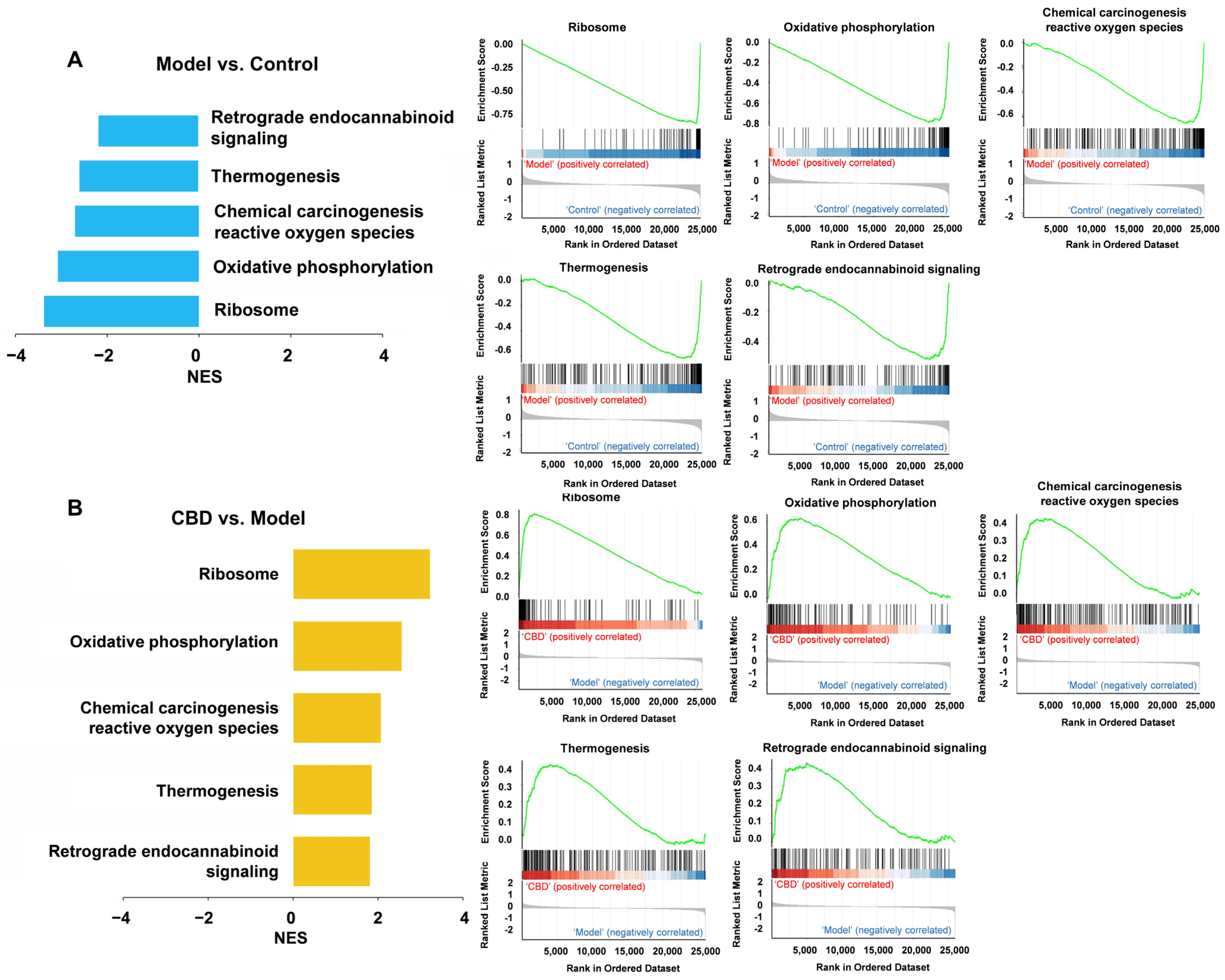
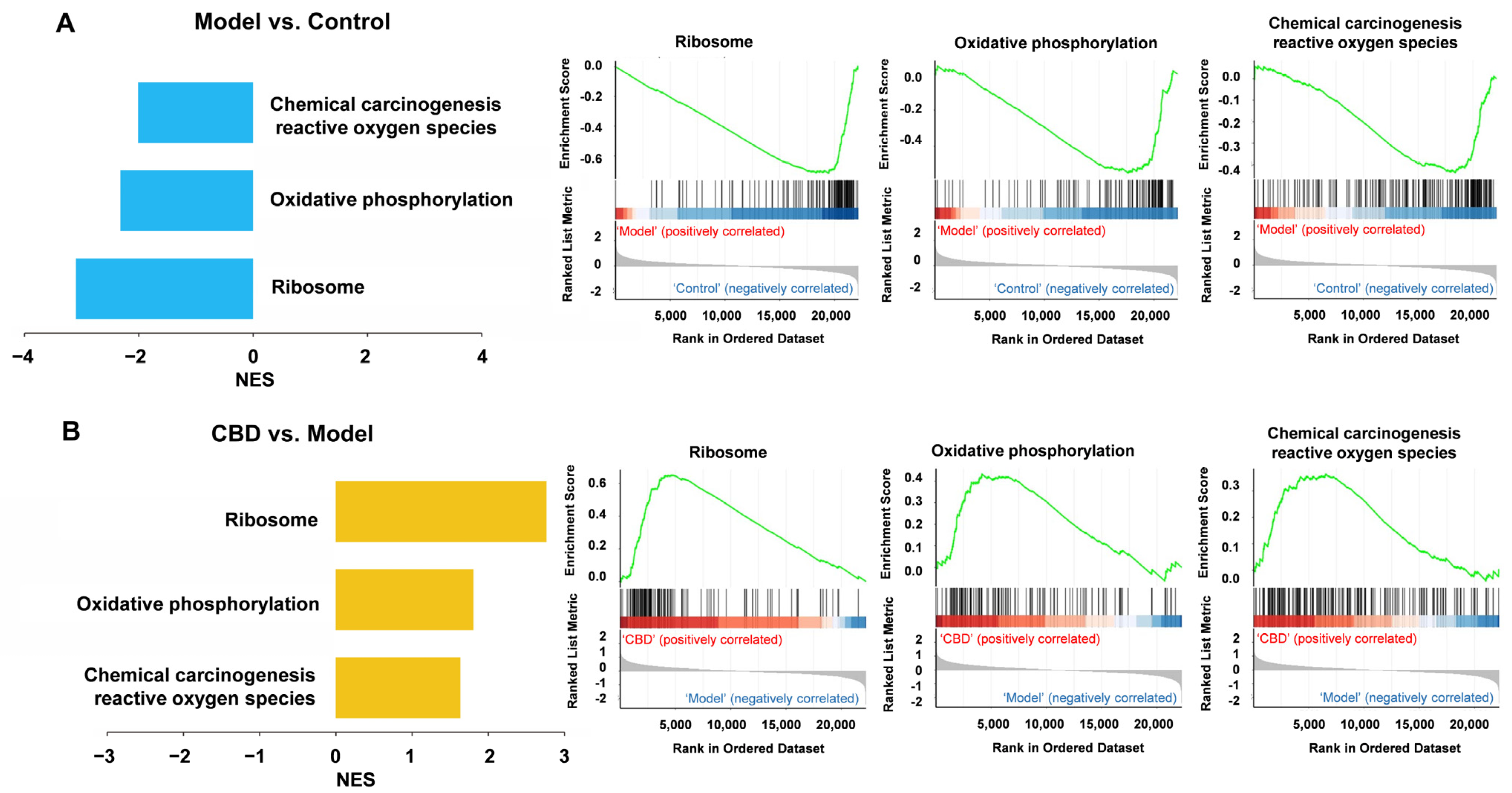
4.6. Gene Set Enrichment Analysis (GSEA) for CBD’s Anti-PTSD Effect in the ExNs and InNs
5. Discussion
5.1. Fear-Based Learning and Memory and the Regulation of CBD in PTSD
5.2. The Endocannabinoid System and the Regulation of CBD in PTSD
5.3. Oxidative Phosphorylation and the Regulation of CBD in PTSD
5.4. Oxidative Stress and the Regulation of CBD in PTSD
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ke, S.; Hartmann, J.; Ressler, K.J.; Liu, Y.; Koenen, K.C. The emerging role of the gut microbiome in posttraumatic stress disorder. Brain Behav. Immun. 2023, 114, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Keya, T.A.; Leela, A.; Habib, N.; Rashid, M.; Bakthavatchalam, P. Mental Health Disorders Due to Disaster Exposure: A Systematic Review and Meta-Analysis. Cureus 2023, 15, e37031. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, N.; Weston, D.; Hall, C.; Caulfield, T.; Williamson, V.; Fong, K. Mental health of staff working in intensive care during COVID-19. Occup. Med. 2021, 71, 62–67. [Google Scholar] [CrossRef]
- Schou, T.M.; Joca, S.; Wegener, G.; Bay-Richter, C. Psychiatric and neuropsychiatric sequelae of COVID-19—A systematic review. Brain Behav. Immun. 2021, 97, 328–348. [Google Scholar] [CrossRef] [PubMed]
- Koenen, K.C.; Sumner, J.A.; Gilsanz, P.; Glymour, M.M.; Ratanatharathorn, A.; Rimm, E.B.; Roberts, A.L.; Winning, A.; Kubzansky, L.D. Post-traumatic stress disorder and cardiometabolic disease: Improving causal inference to inform practice. Psychol. Med. 2017, 47, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.L.; Kubzansky, L.D.; Chibnik, L.B.; Rimm, E.B.; Koenen, K.C. Association of Posttraumatic Stress and Depressive Symptoms with Mortality in Women. JAMA Netw. Open 2020, 3, e2027935. [Google Scholar] [CrossRef]
- Roberts, A.L.; Liu, J.; Lawn, R.B.; Jha, S.C.; Sumner, J.A.; Kang, J.H.; Rimm, E.B.; Grodstein, F.; Kubzansky, L.D.; Chibnik, L.B.; et al. Association of Posttraumatic Stress Disorder with Accelerated Cognitive Decline in Middle-aged Women. JAMA Netw. Open 2022, 5, e2217698. [Google Scholar] [CrossRef] [PubMed]
- Ryder, A.L.; Azcarate, P.M.; Cohen, B.E. PTSD and Physical Health. Curr. Psychiatry Rep. 2018, 20, 116. [Google Scholar] [CrossRef]
- Lupien, S.J.; Juster, R.P.; Raymond, C.; Marin, M.F. The effects of chronic stress on the human brain: From neurotoxicity, to vulnerability, to opportunity. Front. Neuroendocrinol. 2018, 49, 91–105. [Google Scholar] [CrossRef]
- Pronier, E.; Morici, J.F.; Girardeau, G. The role of the hippocampus in the consolidation of emotional memories during sleep. Trends Neurosci. 2023, 46, 912–925. [Google Scholar] [CrossRef]
- Chaaya, N.; Battle, A.R.; Johnson, L.R. An update on contextual fear memory mechanisms: Transition between Amygdala and Hippocampus. Neurosci. Biobehav. Rev. 2018, 92, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, I.; Furini, C.R.; Myskiw, J.C. Fear Memory. Physiol. Rev. 2016, 96, 695–750. [Google Scholar] [CrossRef] [PubMed]
- Popik, B.; Crestani, A.P.; Silva, M.O.; Quillfeldt, J.A.; de Oliveira Alvares, L. Calpain modulates fear memory consolidation, retrieval and reconsolidation in the hippocampus. Neurobiol. Learn. Mem. 2018, 151, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Logue, M.W.; van Rooij, S.J.H.; Dennis, E.L.; Davis, S.L.; Hayes, J.P.; Stevens, J.S.; Densmore, M.; Haswell, C.C.; Ipser, J.; Koch, S.B.J.; et al. Smaller Hippocampal Volume in Posttraumatic Stress Disorder: A Multisite ENIGMA-PGC Study: Subcortical Volumetry Results From Posttraumatic Stress Disorder Consortia. Biol. Psychiatry 2018, 83, 244–253. [Google Scholar] [CrossRef]
- Villarreal, G.; Hamilton, D.A.; Petropoulos, H.; Driscoll, I.; Rowland, L.M.; Griego, J.A.; Kodituwakku, P.W.; Hart, B.L.; Escalona, R.; Brooks, W.M. Reduced hippocampal volume and total white matter volume in posttraumatic stress disorder. Biol. Psychiatry 2002, 52, 119–125. [Google Scholar] [CrossRef]
- Wignall, E.L.; Dickson, J.M.; Vaughan, P.; Farrow, T.F.; Wilkinson, I.D.; Hunter, M.D.; Woodruff, P.W. Smaller hippocampal volume in patients with recent-onset posttraumatic stress disorder. Biol. Psychiatry 2004, 56, 832–836. [Google Scholar] [CrossRef]
- Muhie, S.; Gautam, A.; Chakraborty, N.; Hoke, A.; Meyerhoff, J.; Hammamieh, R.; Jett, M. Molecular indicators of stress-induced neuroinflammation in a mouse model simulating features of post-traumatic stress disorder. Transl. Psychiatry 2017, 7, e1135. [Google Scholar] [CrossRef]
- Muhie, S.; Gautam, A.; Meyerhoff, J.; Chakraborty, N.; Hammamieh, R.; Jett, M. Brain transcriptome profiles in mouse model simulating features of post-traumatic stress disorder. Mol. Brain 2015, 8, 14. [Google Scholar] [CrossRef]
- Tanaka, M.; Li, H.; Zhang, X.; Singh, J.; Dalgard, C.L.; Wilkerson, M.; Zhang, Y. Region- and time-dependent gene regulation in the amygdala and anterior cingulate cortex of a PTSD-like mouse model. Mol. Brain 2019, 12, 25. [Google Scholar] [CrossRef]
- Feng, D.Y.; Guo, B.L.; Liu, G.H.; Xu, K.; Yang, J.; Tao, K.; Huang, J.; Wang, L.Y.; Wang, W.; Wu, S.X. Nerve growth factor against PTSD symptoms: Preventing the impaired hippocampal cytoarchitectures. Prog. Neurobiol. 2020, 184, 101721. [Google Scholar] [CrossRef]
- Ding, X.; Yang, M.; Wu, N.; Li, J.; Song, R. Blockade of dopamine D3 receptor in ventral tegmental area attenuating contextual fear memory. Biomed. Pharmacother. 2023, 158, 114179. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Song, X.; Song, D.; Xie, G.; Guo, H.; Wu, N.; Li, J. Comparison between cannabidiol and sertraline for the modulation of post-traumatic stress disorder-like behaviors and fear memory in mice. Psychopharmacology 2022, 239, 1605–1620. [Google Scholar] [CrossRef] [PubMed]
- Malikowska-Racia, N.; Salat, K.; Nowaczyk, A.; Fijalkowski, L.; Popik, P. Dopamine D2/D3 receptor agonists attenuate PTSD-like symptoms in mice exposed to single prolonged stress. Neuropharmacology 2019, 155, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.G.; Zhang, L.M.; Yao, J.Q.; Yin, Y.Y.; Zhang, X.Y.; Li, Y.F.; Cao, J.B. Anti-PTSD Effects of Hypidone Hydrochloride (YL-0919): A Novel Combined Selective 5-HT Reuptake Inhibitor/5-HT(1A) Receptor Partial Agonist/5-HT(6) Receptor Full Agonist. Front. Pharmacol. 2021, 12, 625547. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Zhu, S.; Ma, J.; Zhu, L.; Liu, Y.; Ou, G.; Li, R.; Wang, Y.; Liang, Y.; Jin, X.; et al. Intranasal temperature-sensitive hydrogels of cannabidiol inclusion complex for the treatment of post-traumatic stress disorder. Acta Pharm. Sin. B 2021, 11, 2031–2047. [Google Scholar] [CrossRef] [PubMed]
- Gazarini, L.; Stern, C.A.; Piornedo, R.R.; Takahashi, R.N.; Bertoglio, L.J. PTSD-like memory generated through enhanced noradrenergic activity is mitigated by a dual step pharmacological intervention targeting its reconsolidation. Int. J. Neuropsychopharmacol. 2014, 18, pyu026. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.A.J.; da Silva, T.R.; Raymundi, A.M.; de Souza, C.P.; Hiroaki-Sato, V.A.; Kato, L.; Guimaraes, F.S.; Andreatini, R.; Takahashi, R.N.; Bertoglio, L.J. Cannabidiol disrupts the consolidation of specific and generalized fear memories via dorsal hippocampus CB(1) and CB(2) receptors. Neuropharmacology 2017, 125, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.A.; Gazarini, L.; Takahashi, R.N.; Guimaraes, F.S.; Bertoglio, L.J. On disruption of fear memory by reconsolidation blockade: Evidence from cannabidiol treatment. Neuropsychopharmacology 2012, 37, 2132–2142. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Chen, R.; Wu, X.; Jiang, L.; Zhang, Y. Single-Cell RNA-Seq Reveals Hypothalamic Cell Diversity. Cell Rep. 2017, 18, 3227–3241. [Google Scholar] [CrossRef]
- Velmeshev, D.; Schirmer, L.; Jung, D.; Haeussler, M.; Perez, Y.; Mayer, S.; Bhaduri, A.; Goyal, N.; Rowitch, D.H.; Kriegstein, A.R. Single-cell genomics identifies cell type-specific molecular changes in autism. Science 2019, 364, 685–689. [Google Scholar] [CrossRef]
- Zeisel, A.; Munoz-Manchado, A.B.; Codeluppi, S.; Lonnerberg, P.; La Manno, G.; Jureus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Chatzinakos, C.; Pernia, C.D.; Morrison, F.G.; Iatrou, A.; McCullough, K.M.; Schuler, H.; Snijders, C.; Bajaj, T.; DiPietro, C.P.; Soliva Estruch, M.; et al. Single-Nucleus Transcriptome Profiling of Dorsolateral Prefrontal Cortex: Mechanistic Roles for Neuronal Gene Expression, Including the 17q21.31 Locus, in PTSD Stress Response. Am. J. Psychiatry 2023, 180, 739–754. [Google Scholar] [CrossRef]
- Chang, C.; Zuo, H.; Li, Y. Recent advances in deciphering hippocampus complexity using single-cell transcriptomics. Neurobiol. Dis. 2023, 179, 106062. [Google Scholar] [CrossRef] [PubMed]
- Acheson, D.T.; Gresack, J.E.; Risbrough, V.B. Hippocampal dysfunction effects on context memory: Possible etiology for posttraumatic stress disorder. Neuropharmacology 2012, 62, 674–685. [Google Scholar] [CrossRef] [PubMed]
- Shin, L.M.; Shin, P.S.; Heckers, S.; Krangel, T.S.; Macklin, M.L.; Orr, S.P.; Lasko, N.; Segal, E.; Makris, N.; Richert, K.; et al. Hippocampal function in posttraumatic stress disorder. Hippocampus 2004, 14, 292–300. [Google Scholar] [CrossRef]
- Desmedt, A.; Marighetto, A.; Piazza, P.V. Abnormal Fear Memory as a Model for Posttraumatic Stress Disorder. Biol. Psychiatry 2015, 78, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Golier, J.A.; Harvey, P.D.; Legge, J.; Yehuda, R. Memory performance in older trauma survivors: Implications for the longitudinal course of PTSD. Ann. N. Y. Acad. Sci. 2006, 1071, 54–66. [Google Scholar] [CrossRef]
- Klann, E.; Sweatt, J.D. Altered protein synthesis is a trigger for long-term memory formation. Neurobiol. Learn. Mem. 2008, 89, 247–259. [Google Scholar] [CrossRef]
- Liberzon, I.; Krstov, M.; Young, E.A. Stress-restress: Effects on ACTH and fast feedback. Psychoneuroendocrinology 1997, 22, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.I.; Yagi, T.; Nanba, T.; Asanuma, M. Application of Single Prolonged Stress Induces Post-traumatic Stress Disorder-like Characteristics in Mice. Acta Med. Okayama 2018, 72, 479–485. [Google Scholar] [PubMed]
- Wang, J.; Gao, F.; Cui, S.; Yang, S.; Gao, F.; Wang, X.; Zhu, G. Utility of 7,8-dihydroxyflavone in preventing astrocytic and synaptic deficits in the hippocampus elicited by PTSD. Pharmacol. Res. 2022, 176, 106079. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xiang, H.; Lu, J.; Chen, Z.; Huang, C.; Yuan, X. Lycopene ameliorates PTSD-like behaviors in mice and rebalances the neuroinflammatory response and oxidative stress in the brain. Physiol. Behav. 2020, 224, 113026. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Watt, H.; Kesavan, C.; Mohan, S. The negative impact of single prolonged stress (SPS) on bone development in mice. Stress 2013, 16, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Uhernik, A.L.; Montoya, Z.T.; Balkissoon, C.D.; Smith, J.P. Learning and memory is modulated by cannabidiol when administered during trace fear-conditioning. Neurobiol. Learn. Mem. 2018, 149, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Jurkus, R.; Day, H.L.; Guimaraes, F.S.; Lee, J.L.; Bertoglio, L.J.; Stevenson, C.W. Cannabidiol Regulation of Learned Fear: Implications for Treating Anxiety-Related Disorders. Front. Pharmacol. 2016, 7, 454. [Google Scholar] [CrossRef]
- Tudor, J.C.; Davis, E.J.; Peixoto, L.; Wimmer, M.E.; van Tilborg, E.; Park, A.J.; Poplawski, S.G.; Chung, C.W.; Havekes, R.; Huang, J.; et al. Sleep deprivation impairs memory by attenuating mTORC1-dependent protein synthesis. Sci. Signal. 2016, 9, ra41. [Google Scholar] [CrossRef] [PubMed]
- Roy-Byrne, P.P.; Cowley, D.S. Search for pathophysiology of panic disorder. Lancet 1998, 352, 1646–1647. [Google Scholar] [CrossRef]
- Sherrin, T.; Blank, T.; Saravana, R.; Rayner, M.; Spiess, J.; Todorovic, C. Region specific gene expression profile in mouse brain after chronic corticotropin releasing factor receptor 1 activation: The novel role for diazepam binding inhibitor in contextual fear conditioning. Neuroscience 2009, 162, 14–22. [Google Scholar] [CrossRef]
- Siiskonen, H.; Oikari, S.; Korhonen, V.P.; Pitkanen, A.; Voikar, V.; Kettunen, M.; Hakumaki, J.; Wahlfors, T.; Pussinen, R.; Penttonen, M.; et al. Diazepam binding inhibitor overexpression in mice causes hydrocephalus, decreases plasticity in excitatory synapses and impairs hippocampus-dependent learning. Mol. Cell. Neurosci. 2007, 34, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Guo, Y.; Yao, J.Q.; Fang, X.X.; Sun, L.J.; Jiang, X.Y.; Ding, Z.C.; Ran, Y.H.; Wang, H.L.; Zhang, L.M.; et al. Rapid anti-PTSD-like activity of the TSPO agonist YL-IPA08: Emphasis on brain GABA, neurosteroids and HPA axis function. Behav. Brain Res. 2020, 379, 112320. [Google Scholar] [CrossRef] [PubMed]
- Asaro, A.; Carlo-Spiewok, A.S.; Malik, A.R.; Rothe, M.; Schipke, C.G.; Peters, O.; Heeren, J.; Willnow, T.E. Apolipoprotein E4 disrupts the neuroprotective action of sortilin in neuronal lipid metabolism and endocannabinoid signaling. Alzheimer’s Dement. 2020, 16, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The Endocannabinoid System: A Potential Target for the Treatment of Various Diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Parker, L.A. The endocannabinoid system and the brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Pinilla, P.; Lopez-Gil, J.; Crespo-Facorro, B. Immune system: A possible nexus between cannabinoids and psychosis. Brain Behav. Immun. 2014, 40, 269–282. [Google Scholar] [CrossRef]
- Patel, S.; Hill, M.N.; Cheer, J.F.; Wotjak, C.T.; Holmes, A. The endocannabinoid system as a target for novel anxiolytic drugs. Neurosci. Biobehav. Rev. 2017, 76, 56–66. [Google Scholar] [CrossRef]
- Fidelman, S.; Mizrachi Zer-Aviv, T.; Lange, R.; Hillard, C.J.; Akirav, I. Chronic treatment with URB597 ameliorates post-stress symptoms in a rat model of PTSD. Eur. Neuropsychopharmacol. 2018, 28, 630–642. [Google Scholar] [CrossRef]
- Xue, F.; Xue, S.S.; Liu, L.; Sang, H.F.; Ma, Q.R.; Tan, Q.R.; Wang, H.N.; Zhou, C.H.; Peng, Z.W. Early intervention with electroacupuncture prevents PTSD-like behaviors in rats through enhancing hippocampal endocannabinoid signaling. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 93, 171–181. [Google Scholar] [CrossRef]
- Reich, C.G.; Mihalik, G.R.; Iskander, A.N.; Seckler, J.C.; Weiss, M.S. Adolescent chronic mild stress alters hippocampal CB1 receptor-mediated excitatory neurotransmission and plasticity. Neuroscience 2013, 253, 444–454. [Google Scholar] [CrossRef]
- Su, Y.A.; Wu, J.; Zhang, L.; Zhang, Q.; Su, D.M.; He, P.; Wang, B.D.; Li, H.; Webster, M.J.; Traumatic Stress Brain Study, G.; et al. Dysregulated mitochondrial genes and networks with drug targets in postmortem brain of patients with posttraumatic stress disorder (PTSD) revealed by human mitochondria-focused cDNA microarrays. Int. J. Biol. Sci. 2008, 4, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Prajapati, S.K.; Krishnamurthy, S. Supplementation of taurine improves ionic homeostasis and mitochondrial function in the rats exhibiting post-traumatic stress disorder-like symptoms. Eur. J. Pharmacol. 2021, 908, 174361. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yu, Z.; Zhang, Y.; Huang, X.; Hou, J.; Zhao, Y.; Luo, W.; Chen, L.; Ou, L.; Li, H.; et al. Iron-induced neuronal damage in a rat model of post-traumatic stress disorder. Neuroscience 2016, 330, 90–99. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Li, Y.; Tang, Y.; Kong, X.; Feng, Z.; Anthony, T.G.; Watford, M.; Hou, Y.; Wu, G.; et al. The role of leucine and its metabolites in protein and energy metabolism. Amino Acids. 2016, 48, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, L.M.; Tandoc, K.; Topisirovic, I.; Furic, L. Cross-talk between protein synthesis, energy metabolism and autophagy in cancer. Curr. Opin. Genet. Dev. 2018, 48, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Henningsen, K.; Palmfeldt, J.; Christiansen, S.; Baiges, I.; Bak, S.; Jensen, O.N.; Gregersen, N.; Wiborg, O. Candidate hippocampal biomarkers of susceptibility and resilience to stress in a rat model of depression. Mol. Cell. Proteom. 2012, 11, M111.016428. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Hu, F.; Wu, J.; Zhang, S. Cannabidiol attenuates OGD/R-induced damage by enhancing mitochondrial bioenergetics and modulating glucose metabolism via pentose-phosphate pathway in hippocampal neurons. Redox Biol. 2017, 11, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Ibork, H.; Idrissi, S.E.; Zulu, S.S.; Miller, R.; Hajji, L.; Morgan, A.M.; Taghzouti, K.; Abboussi, O. Effect of Cannabidiol in LPS-Induced Toxicity in Astrocytes: Possible Role for Cannabinoid Type-1 Receptors. Neurotox Res. 2023, 41, 615–626. [Google Scholar] [CrossRef]
- Zieker, J.; Zieker, D.; Jatzko, A.; Dietzsch, J.; Nieselt, K.; Schmitt, A.; Bertsch, T.; Fassbender, K.; Spanagel, R.; Northoff, H.; et al. Differential gene expression in peripheral blood of patients suffering from post-traumatic stress disorder. Mol. Psychiatry 2007, 12, 116–118. [Google Scholar] [CrossRef]
- Borovac Stefanovic, L.; Kalinic, D.; Mimica, N.; Beer Ljubic, B.; Aladrovic, J.; Mandelsamen Perica, M.; Curic, M.; Grosic, P.F.; Delas, I. Oxidative status and the severity of clinical symptoms in patients with post-traumatic stress disorder. Ann. Clin. Biochem. 2015, 52, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Chen, L.; Wang, J.; Wang, X.; Yang, S.; Zhu, G. Polysaccharides from Polygonatum cyrtonema Hua prevent post-traumatic stress disorder behaviors in mice: Mechanisms from the perspective of synaptic injury, oxidative stress, and neuroinflammation. J. Ethnopharmacol. 2024, 319, 117165. [Google Scholar] [CrossRef] [PubMed]
- Ebenezer, P.J.; Wilson, C.B.; Wilson, L.D.; Nair, A.R.; Francis, J. The Anti-Inflammatory Effects of Blueberries in an Animal Model of Post-Traumatic Stress Disorder (PTSD). PLoS ONE 2016, 11, e0160923. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Piao, F.; Zhao, Y.; Li, S.; Wang, Y.; Liu, P. Subchronic exposure to arsenic decreased Sdha expression in the brain of mice. Neurotoxicology 2009, 30, 538–543. [Google Scholar] [CrossRef]
- Shibanuma, M.; Inoue, A.; Ushida, K.; Uchida, T.; Ishikawa, F.; Mori, K.; Nose, K. Importance of mitochondrial dysfunction in oxidative stress response: A comparative study of gene expression profiles. Free Radic. Res. 2011, 45, 672–680. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, G.; Qin, Y.; Wu, N.; Han, X.; Li, J. Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts. Genes 2024, 15, 519. https://doi.org/10.3390/genes15040519
Xie G, Qin Y, Wu N, Han X, Li J. Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts. Genes. 2024; 15(4):519. https://doi.org/10.3390/genes15040519
Chicago/Turabian StyleXie, Guanbo, Yihan Qin, Ning Wu, Xiao Han, and Jin Li. 2024. "Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts" Genes 15, no. 4: 519. https://doi.org/10.3390/genes15040519
APA StyleXie, G., Qin, Y., Wu, N., Han, X., & Li, J. (2024). Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts. Genes, 15(4), 519. https://doi.org/10.3390/genes15040519