
An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats
 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples Collection
2.2. Skeletal Muscle Satellite Cells (MuSCs) Isolation and Identification
2.3. Cell Culture and Transfection
2.4. Plasmid Construction and RNA Stability Assays
2.5. Total RNA Isolation and qPCR
2.6. Luciferase Reporter Assays
2.7. Extraction of Genomic DNA and Detection of DNA Quality
2.8. PCR Amplification and Sequencing
2.9. MassARRAY Genotyping
2.10. Growth Trait Determination
2.11. Bioinformatics Analysis and Data Analysis
3. Results
3.1. The Synonymous Mutation rs672215506 Affected the mRNA Stability of B4GALNT2 Gene
3.2. The Detection and Functional Verification of Non-Coding SNPs in B4GALNT2
3.3. Population Genetic Diversity Statistics of Six SNPs in Nanjiang Yellow Goat Population
3.4. Analysis of Linkage Disequilibrium and Construction of Haplotypes
3.5. Association of SNPs and Haplotype Combinations with Growth Traits
3.6. Association of SNPs and Haplotype Combinations with Lambing Number
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byrne, G.; Ahmad-Villiers, S.; Du, Z.; McGregor, C. B4GALNT2 and xenotransplantation: A newly appreciated xenogeneic antigen. Xenotransplantation 2018, 25, e12394. [Google Scholar] [CrossRef] [PubMed]
- Stwora-Wojczyk, M.M.; Kissinger, J.C.; Spitalnik, S.L.; Wojczyk, B.S. O-glycosylation in Toxoplasma gondii: Identification and analysis of a family of UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferases. Int. J. Parasitol. 2004, 34, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cooper, D.K.C.; Dai, Y.; Hara, H.; Cai, Z.; Mou, L. The Sda and Cad glycan antigens and their glycosyltransferase, β1,4GalNAcT-II, in xenotransplantation. Xenotransplantation 2018, 25, e12386. [Google Scholar] [CrossRef] [PubMed]
- Duca, M.; Malagolini, N.; Dall’Olio, F. The story of the Sd(a) antigen and of its cognate enzyme B4GALNT2: What is new? Glycoconj. J. 2023, 40, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Groux-Degroote, S.; Vicogne, D.; Cogez, V.; Schulz, C.; Harduin-Lepers, A. B4GALNT2 Controls Sd(a) and SLe(x) Antigen Biosynthesis in Healthy and Cancer Human Colon. Chembiochem Eur. J. Chem. Biol. 2021, 22, 3381–3390. [Google Scholar] [CrossRef] [PubMed]
- Galeev, A.; Suwandi, A.; Cepic, A.; Basu, M.; Baines, J.F.; Grassl, G.A. The role of the blood group-related glycosyltransferases FUT2 and B4GALNT2 in susceptibility to infectious disease. Int. J. Med. Microbiol. 2021, 311, 151487. [Google Scholar] [CrossRef] [PubMed]
- Groux-Degroote, S.; Wavelet, C.; Krzewinski-Recchi, M.A.; Portier, L.; Mortuaire, M.; Mihalache, A.; Trinchera, M.; Delannoy, P.; Malagolini, N.; Chiricolo, M.; et al. B4GALNT2 gene expression controls the biosynthesis of Sda and sialyl Lewis X antigens in healthy and cancer human gastrointestinal tract. Int. J. Biochem. Cell Biol. 2014, 53, 442–449. [Google Scholar] [CrossRef]
- Rausch, P.; Steck, N.; Suwandi, A.; Seidel, J.A.; Künzel, S.; Bhullar, K.; Basic, M.; Bleich, A.; Johnsen, J.M.; Vallance, B.A.; et al. Expression of the Blood-Group-Related Gene B4galnt2 Alters Susceptibility to Salmonella Infection. PLoS Pathog. 2015, 11, e1005008. [Google Scholar] [CrossRef]
- Pucci, M.; Gomes Ferreira, I.; Malagolini, N.; Ferracin, M.; Dall’Olio, F. The Sd(a) Synthase B4GALNT2 Reduces Malignancy and Stemness in Colon Cancer Cell Lines Independently of Sialyl Lewis X Inhibition. Int. J. Mol. Sci. 2020, 21, 6558. [Google Scholar] [CrossRef]
- Drouilhet, L.; Mansanet, C.; Sarry, J.; Tabet, K.; Bardou, P.; Woloszyn, F.; Lluch, J.; Harichaux, G.; Viguié, C.; Monniaux, D.; et al. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS Genet. 2013, 9, e1003809. [Google Scholar] [CrossRef]
- Drouilhet, L.; Lecerf, F.; Bodin, L.; Fabre, S.; Mulsant, P. Fine mapping of the FecL locus influencing prolificacy in Lacaune sheep. Anim. Genet. 2009, 40, 804–812. [Google Scholar] [CrossRef]
- Thomas, P.J.; Xu, R.; Martin, P.T. B4GALNT2 (GALGT2) Gene Therapy Reduces Skeletal Muscle Pathology in the FKRP P448L Mouse Model of Limb Girdle Muscular Dystrophy 2I. Am. J. Pathol. 2016, 186, 2429–2448. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Singhal, N.; Serinagaoglu, Y.; Chandrasekharan, K.; Joshi, M.; Bauer, J.A.; Janssen, P.M.; Martin, P.T. Deletion of Galgt2 (B4Galnt2) reduces muscle growth in response to acute injury and increases muscle inflammation and pathology in dystrophin-deficient mice. Am. J. Pathol. 2015, 185, 2668–2684. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.L.; Xu, R.; Martin, P.T. Soluble Heparin Binding Epidermal Growth Factor-Like Growth Factor Is a Regulator of GALGT2 Expression and GALGT2-Dependent Muscle and Neuromuscular Phenotypes. Mol. Cell. Biol. 2019, 39, e00140-19. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, J.; Chao, L.-M.; Cang, M.; Yu, H.; Wang, J.; Bao, S.-Q.; Liu, Y.-B.; Zhang, W.-G.; Ma, Q.; et al. Association analysis of B4GALNT2 gene polymorphisms with the number of lambs in Mongolian and Uzhumuqin sheep. J. Agric. Biotechnol. 2022, 30, 1510–1523. [Google Scholar]
- Rong, X.; Shao, S.; Liang, P.; Zhang, T.-W.; Zou, S.; Meng, K.; Qiang, H.; Feng, D.Z. Polymorphisms of B4GALNT2 and ESR1 genes and their association with the number of lambs in sheep. China Anim. Sci. Vet. 2021, 48, 3332–3342. [Google Scholar] [CrossRef]
- Guo, X.; Wang, X.; Liang, B.; Di, R.; Liu, Q.; Hu, W.; He, X.; Zhang, J.; Zhang, X.; Chu, M. Molecular Cloning of the B4GALNT2 Gene and Its Single Nucleotide Polymorphisms Association with Litter Size in Small Tail Han Sheep. Animals 2018, 8, 160. [Google Scholar] [CrossRef]
- Sa, C.; Wu, T.; Ma, Y.; He, Y.-M.; Ju, L.; He, T.; Wu, Y.; Liu, B. Polymorphisms of four candidate genes for multifetal traits and their association with the number of lambs in cashgoat. China Anim. Sci. Vet. 2023, 50, 1037–1047. [Google Scholar] [CrossRef]
- Yang, B.G.; Yuan, Y.; Zhou, D.K.; Ma, Y.H.; Mahrous, K.F.; Wang, S.Z.; He, Y.M.; Duan, X.H.; Zhang, W.Y.; E, G. Genome-wide selection signal analysis of Australian Boer goat reveals artificial selection imprinting on candidate genes related to muscle development. Anim. Genet. 2021, 52, 550–555. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, L.; Zhong, T.; Wang, L.; Guo, J.; Dong, Y.; Feng, J.; Song, T.; Li, L.; Zhang, H. The differential proliferation and differentiation ability of skeletal muscle satellite cell in Boer and Nanjiang brown goats. Small Rumin. Res. 2018, 169, 99–107. [Google Scholar] [CrossRef]
- Zhao, S.; Cao, J.; Sun, Y.; Zhou, H.; Zhu, Q.; Dai, D.; Zhan, S.; Guo, J.; Zhong, T.; Wang, L.; et al. METTL3 Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Regulating MEF2C mRNA Stability in a m6A-Dependent Manner. Int. J. Mol. Sci. 2023, 24, 14115. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.L.; Heath, D.A.; Reader, K.L.; Quirke, L.D.; Hudson, N.L.; Juengel, J.L.; McNatty, K.P. Oocytes in sheep homozygous for a mutation in bone morphogenetic protein receptor 1B express lower mRNA levels of bone morphogenetic protein 15 but not growth differentiation factor 9. Reproduction 2011, 142, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lu, J.-Y.; Ma, L.; Xia, W.; Hu, L.; Luo, B.; Yang, K. cDNA cloning and tissue expression of goat B4GALNT2 gene. J. Anim. Sci. Vet. Med. 2016, 1, 47–51. [Google Scholar]
- Guo, J.; Tao, H.; Li, P.; Li, L.; Zhong, T.; Wang, L.; Ma, J.; Chen, X.; Song, T.; Zhang, H. Whole-genome sequencing reveals selection signatures associated with important traits in six goat breeds. Sci. Rep. 2018, 8, 10405. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Guo, J.; Li, L.; Zhong, T.; Wang, L.; Zhan, S.; Lu, J.; Wang, D.; Dai, D.; Liu, G.E.; et al. Genetic Diversity and Selection Signatures in Jianchang Black Goats Revealed by Whole-Genome Sequencing Data. Animals 2022, 12, 2365. [Google Scholar] [CrossRef] [PubMed]
- Chaney, J.L.; Clark, P.L. Roles for Synonymous Codon Usage in Protein Biogenesis. Annu. Rev. Biophys. 2015, 44, 143–166. [Google Scholar] [CrossRef]
- Otsuka, H.; Sasai, H.; Nakama, M.; Aoyama, Y.; Abdelkreem, E.; Ohnishi, H.; Konstantopoulou, V.; Sass, J.O.; Fukao, T. Exon 10 skipping in ACAT1 caused by a novel c.949G>A mutation located at an exonic splice enhancer site. Mol. Med. Rep. 2016, 14, 4906–4910. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: The causes and consequences of codon bias. Nat. Rev. Genet. 2011, 12, 32–42. [Google Scholar] [CrossRef]
- Szewczuk, M.; Zych, S.; Wojcik, J.; Czerniawska-Piatkowska, E. Association of two SNPs in the coding region of the insulin-like growth factor 1 receptor (IGF1R) gene with growth-related traits in Angus cattle. J. Appl. Genet. 2013, 54, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Jiang, H.; Liang, Y.-J.; Xiao, C.; Liu, Y.; Jin, J.-G.; Cao, Y. HLF gene polymorphism and its association with muscle fatty acid and amino acid content in sheep. Heilongjiang Anim. Husb. Vet. Med. 2022, 11, 45–49. [Google Scholar]
- Cheng, Y.; Liu, S.; Wang, G.; Wei, W.; Huang, S.; Yang, R.; Geng, H.; Li, H.; Song, J.; Sun, L.; et al. Porcine IGF1 synonymous mutation alter gene expression and protein binding affinity with IGF1R. Int. J. Biol. Macromol. 2018, 116, 23–30. [Google Scholar] [CrossRef]
- Jolma, A.; Yan, J.; Whitington, T.; Toivonen, J.; Nitta, K.R.; Rastas, P.; Morgunova, E.; Enge, M.; Taipale, M.; Wei, G.; et al. DNA-binding specificities of human transcription factors. Cell 2013, 152, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Hoogendoorn, B.; Coleman, S.L.; Guy, C.A.; Smith, S.K.; O’Donovan, M.C.; Buckland, P.R. Functional analysis of polymorphisms in the promoter regions of genes on 22q11. Hum. Mutat. 2004, 24, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, C.; Hao, W.; Yin, W.; Ai, S.; Zhao, Y.; Duan, Z. Novel Single Nucleotide Polymorphisms and Haplotype of MYF5 Gene Are Associated with Body Measurements and Ultrasound Traits in Grassland Short-Tailed Sheep. Genes 2022, 13, 483. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, H.; Chen, X.; Liu, Z.; Zhang, W.; Xia, D. Functional and Activity Analysis of Cattle UCP3 Promoter with MRFs-Related Factors. Int. J. Mol. Sci. 2016, 17, 682. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. Analysis of Transcriptional Regulation in Non-Coding Region of FSHR Gene in Huyang Sheep. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2017. [Google Scholar]
- Kamarudin, N.J.; Wang, V.C.; Tan, X.T.; Ramesh, A.; Ling, M.H. A Simulation Study on the Effects of Founding Population Size and Number of Alleles Per Locus on the Observed Population Genetic Profile: Implications to Broodstock Management. EC Vet. Sci. 2020, 5, 176–180. [Google Scholar]
- Barrandeguy, M.E.; García, M. The Sensitiveness of Expected Heterozygosity and Allelic Richness Estimates for Analyzing Population Genetic Diversity. In Genetic Diversity; Working Title; IntechOpen: London, UK, 2021. [Google Scholar]
- Silió, L.; Rodríguez, M.C.; Fernández, A.; Barragán, C.; Benítez, R.; Óvilo, C.; Fernández, A.I. Measuring inbreeding and inbreeding depression on pig growth from pedigree or SNP-derived metrics. J. Anim. Breed. Genet. 2013, 130, 349–360. [Google Scholar] [CrossRef]
- Penedo, M.; Weisenberger, M.E.; Boyce, W.M.; Johnson, C.K. Wildlife translocation: The conservation implications of pathogen exposure and genetic heterozygosity. BMC Ecol. 2011, 11, 5. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar]
- Jiang, Y.; Wang, S.; Zhu, L.; Yang, H.; Hong, Q. Polymorphism analysis of myostatin gene in Yunshangmontenegrin sheep. Chin. Herbiv. Sci. 2020, 40, 5–7. [Google Scholar]
- Ma, X.; Du, L.; Zhang, L.; Xuan, J.; Wang, H.; Yuan, Z.; Wu, M.; Zhu, C.; Liu, R. Association between RIPK2 gene polymorphism and growth traits in Ujumuqin sheep. Chin. J. Agric. Sci. 2016, 49, 17. [Google Scholar]
- Peng, Y.; Liu, J.; Zhao, S.; Xu, Z.; Zuo, B. Detection of SNPs of RXRB gene in pigs and its association with growth, fattening and reproductive traits. J. Anim. Husb. Vet. Sci. 2021, 52, 596–609. [Google Scholar]
- Meng, K.; Zhang, T.; Liang, P.; Shao, S.; Zou, S.; Rong, X.; Qiang, H.; Feng, D. Analysis of MYF5 gene polymorphism and its association with growth traits in sheep. J. Agric. Biotechnol. 2022, 30, 496–505. [Google Scholar]
- Wu, S.-B.; Franks, T.K.; Hunt, P.; Wirthensohn, M.G.; Gibson, J.P.; Sedgley, M. Discrimination of SNP genotypes associated with complex haplotypes by high resolution melting analysis in almond: Implications for improved marker efficiencies. Mol. Breed. 2009, 25, 351–357. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.P.; Wu, D. Correlation analysis of LH-β gene polymorphism and reproductive performance in Nanjiang yellow sheep. Anim. Husb. Vet. Med. 2006, 38, 3–5. [Google Scholar]

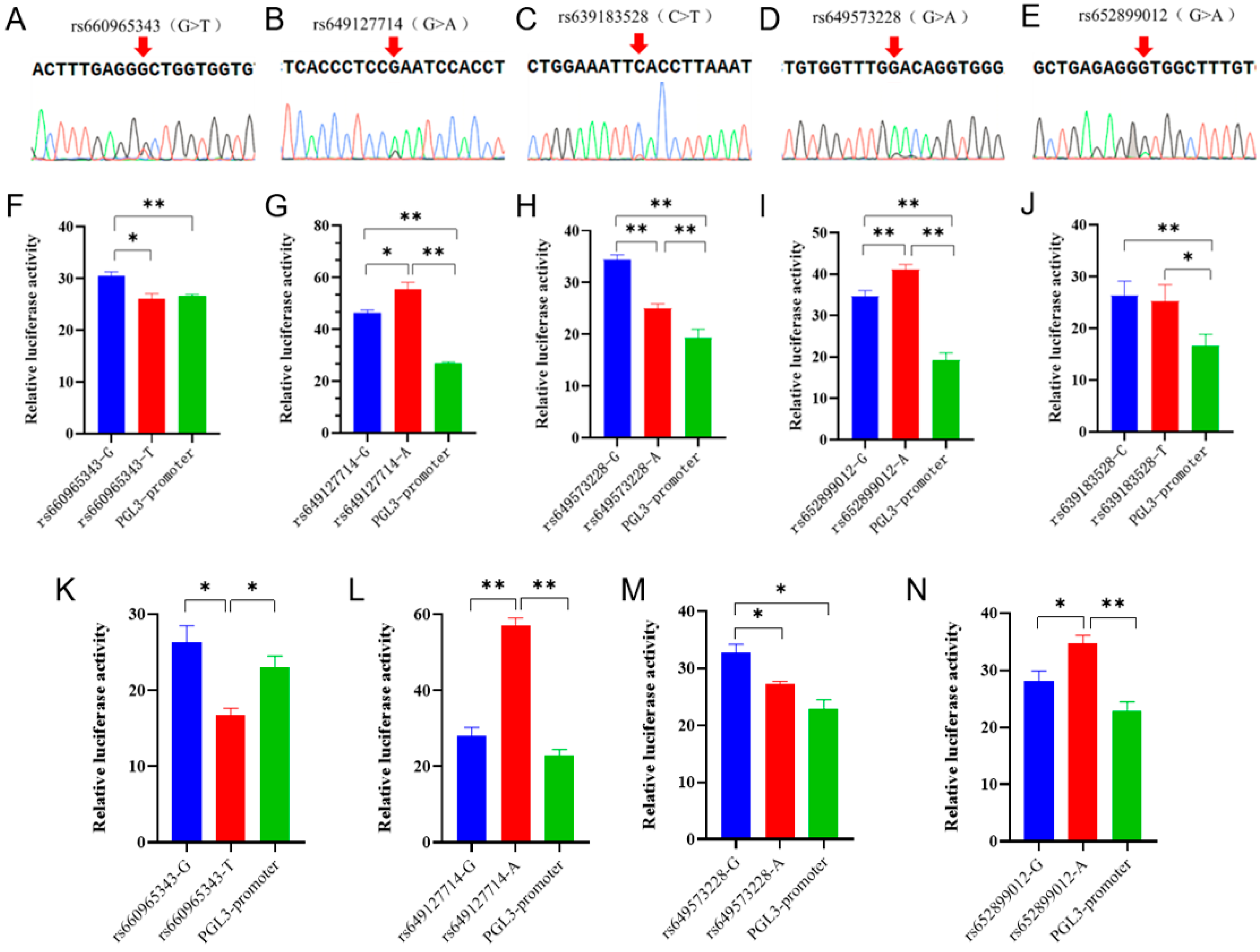


{kind=link}
{kind=link}
{kind=link}
| Locus | Genotype | Genotype Frequency | Allele Frequency | Ho | He | Ne | p | PIC |
|---|---|---|---|---|---|---|---|---|
| rs672215506 (G>A) | AA | 0.28 | 0.51 (A) | 0.5 | 0.5 | 2 | 0.16 | 0.37 |
| GA | 0.46 | 0.49 (G) | ||||||
| GG | 0.26 | |||||||
| rs660965343 (G>T) | GG | 0.4 | 0.64 (G) | 0.54 | 0.46 | 1.85 | 1 | 0.35 |
| GT | 0.46 | 0.36 (T) | ||||||
| TT | 0.14 | |||||||
| rs649127714 (G>A) | GG | 0.08 | 0.29 (G) | 0.59 | 0.41 | 1.7 | 0.58 | 0.33 |
| GA | 0.43 | 0.71 (A) | ||||||
| AA | 0.49 | |||||||
| rs639183528 (C>T) | CC | 0.62 | 0.77 (C) | 0.65 | 0.35 | 1.54 | 0.07 | 0.29 |
| CT | 0.31 | 0.23 (T) | ||||||
| TT | 0.07 | |||||||
| rs649573228 (G>A) | AA | 0.89 | 0.95 (A) | 0.9 | 0.1 | 1.11 | 0.75 | 0.1 |
| GA | 0.11 | 0.05 (G) | ||||||
| rs652899012 (G>A) | GG | 0.58 | 0.65 | 0.35 | 1.54 | 0.09 | 0.29 | |
| AG | 0.39 | 0.77 (G) | ||||||
| AA | 0.03 | 0.23 (A) |
| Haplotype and Haplotype Combination | Type | Genotype | Frequency |
|---|---|---|---|
| Haplotype | H1 | GG | 0.487 |
| H2 | AG | 0.141 | |
| H3 | AT | 0.372 | |
| H1H3 | GGAT | 0.379 | |
| Haplotype combination | H2H2 | GGGG | 0.241 |
| H3H3 | ATAT | 0.124 | |
| H2H3 | AGAT | 0.118 | |
| H1H2 | GGAG | 0.112 | |
| H1H1 | AGAG | 0.026 |
| Locus | Genotype | Number | Birth Weight |
|---|---|---|---|
| rs672215506 (G>A) | AA | 92 | 2.29 ± 0.28 a |
| GA | 149 | 2.26 ± 0.29 ab | |
| GG | 84 | 2.20 ± 0.31 b | |
| rs660965343 (G>T) | TT | 43 | 2.29 ± 0.26 a |
| GT | 151 | 2.27 ± 0.33 ab | |
| GG | 132 | 2.22 ± 0.27 b | |
| rs649127714 (G>A) | AA | 161 | 2.27 ± 0.34 |
| GA | 140 | 2.24 ± 0.24 | |
| GG | 25 | 2.22 ± 0.30 | |
| rs639183528 (C>T) | CC | 200 | 2.26 ± 0.32 |
| CT | 101 | 2.24 ± 0.26 | |
| TT | 23 | 2.23 ± 0.25 | |
| rs649573228 (G>A) | AA | 287 | 2.25 ± 0.31 |
| GA | 35 | 2.24 ± 0.24 | |
| AA | 11 | 2.26 ± 0.26 | |
| rs652899012 (G>A) | AG | 125 | 2.22 ± 0.27 |
| GG | 186 | 2.27 ± 0.32 |
| Locus | rs672215506 (G>A) | rs660965343 (G>T) | rs649127714 (G>A) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Genotype | AA | GA | GG | TT | GT | GG | AA | GA | GG |
| Number | 60 | 88 | 49 | 26 | 89 | 82 | 100 | 82 | 15 |
| BW-6 | 25.99 ± 3.87 | 25.83 ± 4.45 | 26.28 ± 4.58 | 25.42 ± 3.67 b | 25.99 ± 4.16 ab | 26.17 ± 4.64 a | 26.66 ± 4.42 A | 25.26 ± 3.91 B | 25.53 ± 5.03 B |
| BL-6 | 58.85 ± 5.21 | 59.06 ± 5.42 | 59.73 ± 5.53 | 58.00 ± 4.43 B | 59.16 ± 5.23 A | 59.54 ± 5.79 A | 59.79 ± 5.59 A | 58.37 ± 4.98 B | 59.33 ± 5.77 AB |
| BH-6 | 55.80 ± 4.44 B | 56.11 ± 4.96 A | 56.11 ± 4.96 A | 55.23 ± 3.92 B | 56.21 ± 4.79 A | 56.43 ± 4.80 A | 56.71 ± 4.99 A | 55.50 ± 4.24 B | 56.27 ± 4.67 AB |
| CC-6 | 65.19 ± 4.02 | 65.06 ± 4.56 | 65.71 ± 4.83 | 64.37 ± 3.65 B | 65.17 ± 4.23 AB | 65.65 ± 4.92 A | 65.89 ± 4.69 Aa | 64.54 ± 4.09 Bb | 65.07 ± 4.48 ABb |
| BW-12 | 34.08 ± 4.54 | 34.22 ± 5.04 | 34.49 ± 5.31 | 33.37 ± 4.18 Bb | 34.37 ± 4.80 ABa | 34.39 ± 5.33 Aa | 35.02 ± 5.23 A | 33.38 ± 4.33 B | 33.83 ± 5.55 B |
| BL-12 | 65.70 ± 4.76 Bb | 66.11 ± 5.14 ABb | 66.80 ± 5.65 Aa | 64.73 ± 4.19 Bc | 66.13 ± 5.01 Ab | 66.63 ± 5.54 Aa | 66.72 ± 5.46 A | 65.44 ± 4.68 B | 66.33 ± 5.39 AB |
| BH-12 | 62.75 ± 4.04 b | 63.13 ± 4.55 b | 63.73 ± 4.71 a | 62.15 ± 3.56 Bb | 63.08 ± 4.37 ABa | 63.57 ± 4.74 Aa | 63.61 ± 4.69 a | 62.55 ± 4.03 b | 63.53 ± 4.70 a |
| CC-12 | 74.86 ± 4.93 | 75.26 ± 5.28 | 75.87 ± 5.20 | 74.12 ± 4.73 Bb | 75.23 ± 4.92 ABa | 75.73 ± 5.49 Aa | 75.76 ± 5.51 | 74.76 ± 4.74 | 75.10 ± 4.73 |
| BW-18 | 47.23 ± 6.43 | 48.39 ± 7.62 | 48.20 ± 7.45 | 46.33 ± 6.08 B | 48.20 ± 7.37 A | 48.29 ± 7.39 A | 48.67 ± 7.79 a | 47.10 ± 6.08 b | 48.30 ± 8.80 ab |
| BL-18 | 72.78 ± 4.80 | 73.56 ± 5.42 | 73.53 ± 5.65 | 71.35 ± 4.23 B | 73.48 ± 5.11 A | 73.76 ± 5.68 A | 73.77 ± 5.58 a | 72.82 ± 4.84 b | 73.00 ± 5.66 b |
| BH-18 | 69.17 ± 3.95 | 69.30 ± 4.31 | 69.41 ± 4.38 | 68.38 ± 3.42 B | 69.25 ± 4.22 AB | 69.61 ± 4.40 A | 69.69 ± 4.43 A | 68.70 ± 3.69 AB | 69.80 ± 5.10 B |
| CC-18 | 85.52 ± 4.66 | 85.85 ± 5.10 | 85.71 ± 4.88 | 84.46 ± 4.34 B | 85.93 ± 4.62 A | 85.88 ± 5.33 A | 86.32 ± 5.13 a | 85.04 ± 4.37 b | 85.37 ± 5.80 b |
| Locus | rs639183528 (C > T) | rs649573228 (G>A) | rs652899012 (G>A) | ||||||
| Genotype | CC | CT | TT | AA | GA | AA | AG | GG | |
| Number | 119 | 63 | 15 | 174 | 21 | 7 | 71 | 117 | |
| BW-6 | 26.38 ± 4.45 | 25.47 ± 3.85 | 25.10 ± 4.69 | 26.14 ± 4.28 | 24.86 ± 4.56 | 25.71 ± 5.19 | 25.19 ± 4.00 | 26.46 ± 4.41 | |
| BL-6 | 59.45 ± 5.54 a | 58.89 ± 5.03 ab | 58.00 ± 5.57 b | 59.30 ± 5.41 A | 58.05 ± 5.31 B | 60.14 ± 6.15 A | 58.49 ± 5.09 B | 59.56 ± 5.52 AB | |
| BH-6 | 56.45 ± 4.97 | 55.76 ± 4.08 | 55.67 ± 4.82 | 56.32 ± 4.70 A | 55.14 ± 4.67 B | 57.29 ± 5.22 A | 55.48 ± 4.16 B | 56.53 ± 4.97 AB | |
| CC-6 | 65.55 ± 4.72 | 64.92 ± 4.07 | 64.40 ± 3.96 | 65.44 ± 4.47 A | 64.00 ± 4.42 B | 65.71 ± 4.03 a | 64.65 ± 4.23 b | 65.62 ± 4.64 ab | |
| BW-12 | 34.60 ± 5.17 | 33.79 ± 4.47 | 33.30 ± 5.02 | 34.43 ± 4.95 A | 32.67 ± 4.91 B | 34.07 ± 5.76 | 33.48 ± 4.48 | 34.71 ± 5.18 | |
| BL-12 | 66.39 ± 5.34 | 65.87 ± 4.76 | 65.47 ± 5.44 | 66.29 ± 5.22 A | 65.24 ± 4.82 B | 66.57 ± 6.16 a | 65.58 ± 4.72 b | 66.51 ± 5.39 ab | |
| BH-12 | 63.34 ± 4.66 | 62.86 ± 3.95 | 63.00 ± 4.77 | 63.29 ± 4.46 A | 62.24 ± 4.41 B | 63.86 ± 5.24 a | 62.65 ± 4.01 b | 63.43 ± 4.67 ab | |
| CC-12 | 75.45 ± 5.34 | 75.30 ± 5.02 | 73.97 ± 3.95 | 75.41 ± 5.18 | 74.52 ± 5.06 | 75.36 ± 5.02 | 74.80 ± 4.80 | 75.58 ± 5.40 | |
| BW-18 | 48.41 ± 7.58 | 47.40 ± 6.42 | 47.10 ± 7.66 | 48.14 ± 7.34 | 47.05 ± 6.45 | 47.79 ± 8.32 | 47.20 ± 6.63 | 48.53 ± 7.55 | |
| BL-18 | 73.61 ± 5.52 | 72.98 ± 4.91 | 72.40 ± 5.03 | 73.40 ± 5.32 a | 73.00 ± 5.2 b | 73.29 ± 5.88 | 72.77 ± 4.96 | 73.7 ± 5.48 | |
| BH-18 | 69.47 ± 4.34 | 68.94 ± 3.82 | 69.27 ± 4.76 | 69.4 ± 4.24 A | 68.57 ± 4.01 B | 68.71 ± 4.82 | 68.83 ± 3.81 | 69.55 ± 4.34 | |
| CC-18 | 86.02 ± 5.10 | 85.33 ± 4.53 | 84.87 ± 4.81 | 85.84 ± 4.92 A | 84.60 ± 4.91 B | 85.00 ± 4.93 | 85.08 ± 4.72 | 86.13 ± 5.03 | |
| Combined Haplotypes | H2H3 | H3H3 | H1H2 | H1H3 | H2H2 |
|---|---|---|---|---|---|
| Number | 27 | 26 | 26 | 75 | 49 |
| BW-6 | 26.30 ± 3.63 a | 25.42 ± 3.67 b | 25.77 ± 4.67 ab | 25.66 ± 4.42 ab | 26.28 ± 4.58 ab |
| BL-6 | 59.52 ± 5.4 Aa | 58 ± 4.43 Bb | 59.19 ± 6.06 Aa | 58.8 ± 5.29 ABab | 59.73 ± 5.53 Aa |
| BH-6 | 56.11 ± 4.46 Aa | 55.23 ± 3.92 Bb | 55.77 ± 5.04 ABa | 56.07 ± 5.02 ABa | 56.73 ± 4.51 Aa |
| CC-6 | 65.70 ± 3.71 Aa | 64.37 ± 3.65 Bc | 65.35 ± 4.91 Aab | 64.74 ± 4.40 ABbc | 65.71 ± 4.83 Aab |
| BW-12 | 34.33 ± 4.46 a | 33.37 ± 4.18 b | 33.83 ± 5.30 ab | 34.14 ± 4.95 ab | 34.49 ± 5.31 a |
| BL-12 | 66.07 ± 4.82 Aab | 64.73 ± 4.19 Bc | 66.00 ± 5.29 Aab | 66.02 ± 5.09 Ab | 66.80 ± 5.65 Aa |
| BH-12 | 62.85 ± 4.03 ABa | 62.15 ± 3.56 Bb | 63.00 ± 4.67 ABa | 63.01 ± 4.47 ABb | 63.73 ± 4.71 Aa |
| CC-12 | 75.35 ± 4.90 Aa | 74.12 ± 4.73 Bb | 75.46 ± 6.05 Aa | 74.88 ± 4.94 Bb | 75.87 ± 5.20 Aa |
| BW-18 | 47.76 ± 6.53 Aa | 46.33 ± 6.08 Bb | 48.38 ± 7.45 Aa | 47.93 ± 7.45 Ba | 48.20 ± 7.45 Aa |
| BL-18 | 73.56 ± 4.55 Aab | 71.35 ± 4.23 Bc | 73.81 ± 5.64 Aa | 73.2 ± 5.22 Ab | 73.53 ± 5.65 Aab |
| BH-18 | 69.52 ± 4.05 Aab | 68.38 ± 3.42 Bc | 69.69 ± 4.34 Aa | 69.03 ± 4.16 Ab | 69.41 ± 4.38 Aab |
| CC-18 | 86.33 ± 4.07 Aa | 84.46 ± 4.34 Bb | 86.08 ± 5.73 Aa | 85.61 ± 4.88 ABab | 85.71 ± 4.88 ABa |
| Combined Haplotypes | Number | Primiparity | Multiparity |
|---|---|---|---|
| H2H3 | 21 | 1.52 ± 0.51 | 1.82 ± 0.26 Cc |
| H3H3 | 23 | 1.74 ± 0.45 | 1.95 ± 0.15 BCb |
| H1H2 | 18 | 1.67 ± 0.49 | 1.94 ± 0.24 BCb |
| H1H3 | 47 | 1.57 ± 0.54 | 1.98 ± 0.21 Bb |
| H2H2 | 34 | 1.65 ± 0.54 | 2.13 ± 0.17 Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Chen, Z.; Chen, S.; Chen, Y.; Guo, J.; Zhong, T.; Wang, L.; Zhan, S.; Li, L.; Zhang, H.; et al. An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats. Genes 2024, 15, 330. https://doi.org/10.3390/genes15030330
Xu L, Chen Z, Chen S, Chen Y, Guo J, Zhong T, Wang L, Zhan S, Li L, Zhang H, et al. An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats. Genes. 2024; 15(3):330. https://doi.org/10.3390/genes15030330
Chicago/Turabian StyleXu, Liang, Zitong Chen, Shuheng Chen, Yu Chen, Jiazhong Guo, Tao Zhong, Linjie Wang, Siyuan Zhan, Li Li, Hongping Zhang, and et al. 2024. "An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats" Genes 15, no. 3: 330. https://doi.org/10.3390/genes15030330
APA StyleXu, L., Chen, Z., Chen, S., Chen, Y., Guo, J., Zhong, T., Wang, L., Zhan, S., Li, L., Zhang, H., & Cao, J. (2024). An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats. Genes, 15(3), 330. https://doi.org/10.3390/genes15030330