
Comparison of Growth Performance, Carcass Properties, Fatty Acid Profile, and Genes Involved in Fat Metabolism in Nanyang and Landrace Pigs
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, Diets, and Management
2.2. Sample Collection
2.3. Assay of Lipid-Producing Enzyme Activities in Subcutaneous Fat Tissue
2.4. Assay of Fatty Acid Profile in Subcutaneous Fat Tissue
2.5. Assay of Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. Growth Performance and Carcass Traits
3.2. Assay of Lipid-Producing Enzyme Activities in Subcutaneous Fat Tissue
3.3. Assay of Fatty Acid Profile of Subcutaneous Fat Tissue
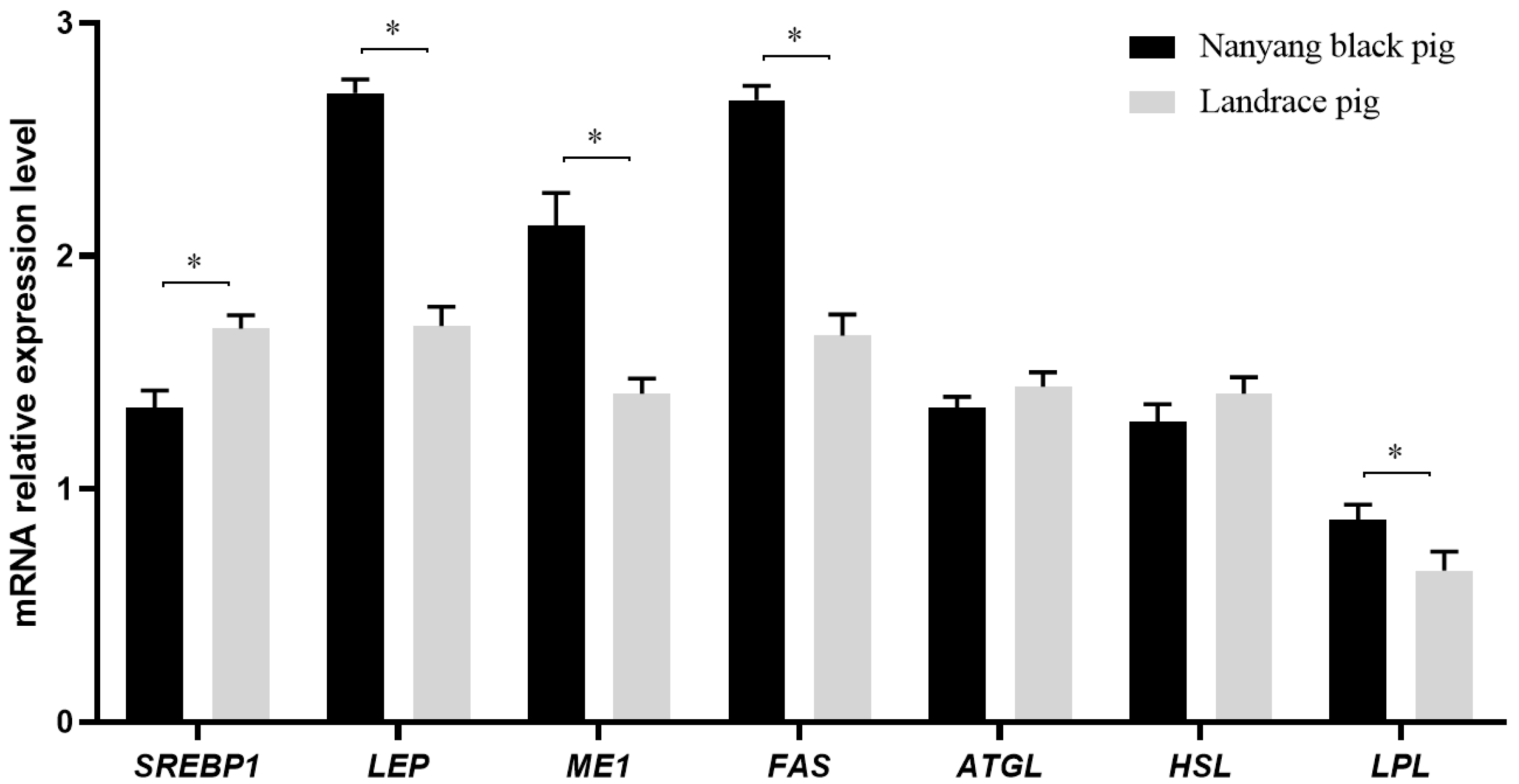
3.4. Assay of Expression of mRNAs Involved in Fat Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schumacher, M.; DelCurto-Wyffels, H.; Thomson, J.; Boles, J. Fat deposition and fat effects on meat quality: A review. Animals 2022, 12, 1550. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Yang, D.D.; Liu, Z.L.; Zeng, Y.Q.; Chen, W. Expression of lipid metabolism genes provides new insights into intramuscular fat deposition in Laiwu pigs. Asian-Australas. J. Anim. Sci. 2020, 33, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liao, Q.; Sun, Y.; Pan, T.; Liu, S.; Miao, W.; Li, Y.; Zhou, L.; Xu, G. Lipidomic and transcriptomic analysis of the longissimus muscle of Luchuan and Duroc pigs. Front. Nutr. 2021, 8, 667622. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Wei, P.; Khan, M.A.; Zhang, J.; Guo, L.; Liu, D.; Zhang, X.; Bai, Y.; Wang, S. Transcriptome analysis reveals differential gene expression in intramuscular adipose tissues of Jinhua and Landrace pigs. J. Vet. Med. Sci. 2018, 80, 953–959. [Google Scholar] [CrossRef]
- Poklukar, K.; Candek-Potokar, M.; Batorek Lukac, N.; Tomazin, U.; Skrlep, M. Lipid deposition and metabolism in local and modern pig breeds: A review. Animals 2020, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, K.; Yang, Q.; Du, M.; Liu, X.; Cao, G. Enhanced adipogenesis in Mashen pigs compared with Large White pigs. Ital. J. Anim. Sci. 2017, 16, 217–225. [Google Scholar] [CrossRef]
- Barea, R.; Isabel, B.; Nieto, R.; Lopez-Bote, C.; Aguilera, J.F. Evolution of the fatty acid profile of subcutaneous back-fat adipose tissue in growing Iberian and Landrace × Large White pigs. Animal 2013, 7, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.H.; Lu, J.X.; Chen, Y.; Zhao, Y.Q.; Guo, P.H.; Yang, J.T.; Zang, R.X. Comparison of the adipogenesis in intramuscular and subcutaneous adipocytes from Bamei and Landrace pigs. Biochem. Cell Biol. 2014, 92, 259–267. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.; Zhang, B.; Zhong, H.; Lu, Y.; Zhang, H. Candidate gene screening for lipid deposition using combined transcriptomic and proteomic data from Nanyang black pigs. BMC Genom. 2021, 22, 441. [Google Scholar] [CrossRef]
- Nong, Q.; Wang, L.; Zhou, Y.; Sun, Y.; Chen, W.; Xie, J.; Zhu, X.; Shan, T. Low dietary n-6/n-3 PUFA ratio regulates meat quality, reduces triglyceride content, and improves fatty acid composition of meat in Heigai pigs. Animals 2020, 10, 1543. [Google Scholar] [CrossRef]
- Song, B.; Di, S.; Cui, S.; Chen, N.; Wang, H.; Wang, X.; Gao, Q.; Tong, G.; Wang, H.; Huang, X.; et al. Distinct patterns of PPARgamma promoter usage, lipid degradation activity, and gene expression in subcutaneous adipose tissue of lean and obese swine. Int. J. Mol. Sci. 2018, 19, 3892. [Google Scholar] [CrossRef]
- Gerfault, V.; Louveau, I.; Mourot, J.; Le Dividich, J. Lipogenic enzyme activities in subcutaneous adipose tissue and skeletal muscle from neonatal pigs consuming maternal or formula milk. Reprod. Nutr. Dev. 2000, 40, 103–112. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Renaudeau, D.; Mourot, J. A comparison of carcass and meat quality characteristics of Creole and Large White pigs slaughtered at 90kg BW. Meat Sci. 2007, 76, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Serra, X.; Gil, F.; Pérez-Enciso, M.; Oliver, M.A.; Vázquez, J.M.; Gispert, M.; Díaz, I.; Moreno, F.; Latorre, R.; Noguera, J.L. A comparison of carcass, meat quality and histochemical characteristics of Iberian (Guadyerbas line) and Landrace pigs. Livest. Prod. Sci. 1998, 56, 215–223. [Google Scholar] [CrossRef]
- Kojima, M.; Nakajima, I.; Arakawa, A.; Mikawa, S.; Matsumoto, T.; Uenishi, H.; Nakamura, Y.; Taniguchi, M. Differences in gene expression profiles for subcutaneous adipose, liver, and skeletal muscle tissues between Meishan and Landrace pigs with different backfat thicknesses. PLoS ONE 2018, 13, e0204135. [Google Scholar] [CrossRef] [PubMed]
- Benitez, R.; Fernandez, A.; Isabel, B.; Nunez, Y.; De Mercado, E.; Gomez-Izquierdo, E.; Garcia-Casco, J.; Lopez-Bote, C.; Ovilo, C. Modulatory effects of breed, feeding status, and diet on adipogenic, lipogenic, and lipolytic gene expression in growing Iberian and Duroc pigs. Int. J. Mol. Sci. 2017, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.G.; Wang, L.J.; Xu, Z.R.; Huang, J.F.; Wang, Y.R. Developmental changes of carcass composition, meat quality and organs in the Jinhua pig and Landrace. Animal 2009, 3, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Gispert, M.; Font, I.F.M.; Gil, M.; Velarde, A.; Diestre, A.; Carrion, D.; Sosnicki, A.A.; Plastow, G.S. Relationships between carcass quality parameters and genetic types. Meat Sci. 2007, 77, 397–404. [Google Scholar] [CrossRef]
- Madeira, M.S.; Pires, V.M.; Alfaia, C.M.; Costa, A.S.; Luxton, R.; Doran, O.; Bessa, R.J.; Prates, J.A. Differential effects of reduced protein diets on fatty acid composition and gene expression in muscle and subcutaneous adipose tissue of Alentejana purebred and Large White × Landrace × Pietrain crossbred pigs. Br. J. Nutr. 2013, 110, 216–229. [Google Scholar] [CrossRef]
- Song, C.H.; Oh, S.M.; Lee, S.; Choi, Y.; Kim, J.D.; Jang, A.; Kim, J. The ratio of dietary n-3 polyunsaturated fatty acids influences the fat composition and lipogenic enzyme activity in adipose tissue of growing pigs. Food Sci. Anim. Resour. 2020, 40, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, J.F.; Hernández-Matamoros, A.; González, E. Free-range and low-protein concentrated diets in Iberian pigs: Effect on plasma insulin and leptin concentration, lipogenic enzyme activity, and fatty acid composition of adipose tissue. Animals 2020, 10, 1917. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, W.; Li, S.; Guo, D.; He, J.; Wang, Y. Acetyl-coA carboxylases and diseases. Front. Oncol. 2022, 12, 836058. [Google Scholar] [CrossRef]
- Jones, S.F.; Infante, J.R. Molecular pathways: Fatty acid synthase. Clin. Cancer Res. 2015, 21, 5434–5438. [Google Scholar] [CrossRef] [PubMed]
- Stanton, R.C. Glucose-6-phosphate dehydrogenase, NADPH, and cell survival. IUBMB Life 2012, 64, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Simmen, F.A.; Alhallak, I.; Simmen, R.C.M. Malic enzyme 1 (ME1) in the biology of cancer: It is not just intermediary metabolism. J. Mol. Endocrinol. 2020, 65, R77–R90. [Google Scholar] [CrossRef] [PubMed]
- Freire, J.P.; Mourot, J.; Cunha, L.F.; Almeida, J.A.; Aumaitre, A. Effect of the source of dietary fat on postweaning lipogenesis in lean and fat pigs. Ann. Nutr. Metab. 1998, 42, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Palma-Granados, P.; Seiquer, I.; Benitez, R.; Ovilo, C.; Nieto, R. Effects of lysine deficiency on carcass composition and activity and gene expression of lipogenic enzymes in muscles and backfat adipose tissue of fatty and lean piglets. Animal 2019, 13, 2406–2418. [Google Scholar] [CrossRef] [PubMed]
- Kasprzyk, A.; Tyra, M.; Babicz, M. Fatty acid profile of pork from a local and a commercial breed. Arch. Anim. Breed. 2015, 58, 379–385. [Google Scholar] [CrossRef]
- Nguyen, T.X.; Agazzi, A.; Comi, M.; Bontempo, V.; Guido, I.; Panseri, S.; Sauerwein, H.; Eckersall, P.D.; Burchmore, R.; Savoini, G. Effects of low ω6:ω3 ratio in sow diet and seaweed supplement in piglet diet on performance, colostrum and milk fatty acid profiles, and oxidative status. Animals 2020, 10, 2049. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Wang, K.; Ao, H.; Chen, S.; Tan, Z.; Wang, Y.; Xitong, Z.; Yang, T.; Zhang, F.; Liu, Y.; et al. Comparative adipose transcriptome analysis digs out genes related to fat deposition in two pig breeds. Sci. Rep. 2019, 9, 12925. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Items | Content | |
|---|---|---|
| 20–60 kg | 61–100 kg | |
| Ingredient (%) | ||
| Corn | 67.4 | 78.3 |
| Soybean meal | 19.5 | 15 |
| Wheat bran | 8.53 | 3.95 |
| Limestone | 0.89 | 0.55 |
| Soybean oil | 2.38 | 0.9 |
| NaCl | 0.3 | 0.3 |
| Premix 1 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 |
| Nutrient level 2 | ||
| Metabolic energy ME (MJ·kg−1) | 14.21 | 14.20 |
| Crude protein CP (%) | 16.16 | 14.17 |
| Calcium Ca (%) | 0.65 | 0.6 |
| Total phosphorus TP (%) | 0.45 | 0.35 |
| Available phosphorus AP (%) | 0.31 | 0.25 |
| Met + Cys (%) | 0.57 | 0.42 |
| Lysine Lys (%) | 1.17 | 1.03 |
| SID lysine (%) | 1.05 | 0.93 |
| Gene | ID | Sequence (5′-3′) | Product Length (bp) |
|---|---|---|---|
| β-actin | XM_021086047 | ACCTTCTACAACGAGCTGCGTG GTCTCCGGAGTCCATCACGATG | 207 |
| SREBP1 | NM_214157 | CTGGTGGTGGGCACGGAGGC TGCCACTGCCACCGCTGCTGC | 316 |
| LEP | U63540 | ACTGCCCCAGAAGCACATCC ATGGAGCCCAGGGATGAAG | 232 |
| ME1 | XM_001924333 | ACTGATGGAGAACGTATTC ACTGCCTCCATGAATTCATCC | 245 |
| FAS | EF589048 | TGTCTGGGTGGGTGTGAGCAG GAGTTAGGCTTCAGCAGGAC | 270 |
| ATGL | EU047807 | TTCGCGGGTTGCGGCTTCCTC GATGGTCTTCACCAGGTTGAA | 249 |
| HSL | AY686758 | CACAGCATGGACCTGCGCACA TCGAAGAGGTGTGCCACACTC | 191 |
| LPL | AY686761 | AGATGGAGAGCAAAGCCCTGC GGGACCCAACTTTCATACAT | 289 |
| Genetic Type | ||||
|---|---|---|---|---|
| Item | Nanyang Pigs | Landrace Pigs | SEM | p-Value |
| IBW (kg) | 22.16 | 21.37 | 0.677 | 0.172 |
| FBW (kg) | 74.85 b | 94.89 a | 2.621 | <0.001 |
| ADG (g/d) | 585.4 b | 816.9 a | 29.587 | <0.001 |
| ADFI (g/d) | 2254.1 | 2369.0 | 90.414 | 0.116 |
| F/G | 3.85 a | 2.90 b | 0.126 | 0.001 |
| Genetic Type | ||||
|---|---|---|---|---|
| Item | Nanyang Pigs | Landrace Pigs | SEM | p-Value |
| Carcass weight (kg) | 52.50 b | 68.27 a | 8.795 | 0.003 |
| Loin muscle area (cm2) | 27.70 b | 45.77 a | 4.273 | 0.001 |
| Lean percentage (%) | 50.39 b | 61.38 a | 6.393 | 0.005 |
| Average backfat thickness 1 (mm) | 22.11 a | 15.35 b | 3.883 | 0.003 |
| Genetic Type | ||||
|---|---|---|---|---|
| Item | Nanyang Pigs | Landrace Pigs | SEM | p-Value |
| ACC 1 | 23.21 a | 16.82 b | 1.817 | 0.010 |
| G6PDH 2 | 345.91 a | 213.63 b | 12.245 | 0.001 |
| ME 3 | 452.72 a | 321.84 b | 18.487 | 0.002 |
| FAS 4 | 51.27 a | 34.03 b | 2.313 | 0.001 |
| Genetic Type | ||||
|---|---|---|---|---|
| Item | Nanyang Pigs | Landrace Pigs | SEM | p-Value |
| C14:0 | 1.52 a | 1.32 b | 0.136 | 0.047 |
| C16:0 | 27.57 | 25.76 | 0.804 | 0.052 |
| C16:1 n-7 | 3.85 | 3.27 | 0.577 | 0.252 |
| C18:0 | 13.66 | 13.94 | 0.446 | 0.494 |
| C18:1 n-7 | 3.24 | 3.33 | 0.188 | 0.603 |
| C18:1 n-9 | 37.56 a | 35.22 b | 1.407 | 0.011 |
| C18:2 n-6 | 7.28 b | 12.42 a | 2.957 | 0.003 |
| C18:3 n-3 | 1.34 b | 1.42 a | 0.047 | 0.014 |
| C20:0 | 0.27 | 0.24 | 0.024 | 0.284 |
| C20:1 n-9 | 1.26 a | 0.96 b | 0.177 | 0.040 |
| C20:3 n-6 | 0.06 | 0.07 | 0.016 | 0.670 |
| C20:4 n-6 | 2.18 a | 1.83 b | 0.219 | 0.023 |
| C22:4 n-6 | 0.14 | 0.16 | 0.014 | 0.294 |
| C22:5 n-3 | 0.06 | 0.07 | 0.012 | 0.349 |
| Total SFA | 43.02 | 41.27 | 0.309 | 0.097 |
| Total MUFA | 45.91 a | 42.77 b | 1.787 | 0.002 |
| PUFA n3 | 1.40 | 1.49 | 0.113 | 0.351 |
| PUFA n6 | 9.66 b | 14.48 a | 0.516 | <0.001 |
| PUFA n3/n6 | 6.90 b | 9.72 a | 0.375 | <0.001 |
| Total PUFA | 11.07 b | 15.96 a | 1.440 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Meng, S.; Wang, H.; Zhang, C.; Sun, Z.; Huang, L.; Miao, Z. Comparison of Growth Performance, Carcass Properties, Fatty Acid Profile, and Genes Involved in Fat Metabolism in Nanyang and Landrace Pigs. Genes 2024, 15, 186. https://doi.org/10.3390/genes15020186
Zhang J, Meng S, Wang H, Zhang C, Sun Z, Huang L, Miao Z. Comparison of Growth Performance, Carcass Properties, Fatty Acid Profile, and Genes Involved in Fat Metabolism in Nanyang and Landrace Pigs. Genes. 2024; 15(2):186. https://doi.org/10.3390/genes15020186
Chicago/Turabian StyleZhang, Jinzhou, Shuaitao Meng, Heming Wang, Chuankuan Zhang, Zhe Sun, Luyao Huang, and Zhiguo Miao. 2024. "Comparison of Growth Performance, Carcass Properties, Fatty Acid Profile, and Genes Involved in Fat Metabolism in Nanyang and Landrace Pigs" Genes 15, no. 2: 186. https://doi.org/10.3390/genes15020186
APA StyleZhang, J., Meng, S., Wang, H., Zhang, C., Sun, Z., Huang, L., & Miao, Z. (2024). Comparison of Growth Performance, Carcass Properties, Fatty Acid Profile, and Genes Involved in Fat Metabolism in Nanyang and Landrace Pigs. Genes, 15(2), 186. https://doi.org/10.3390/genes15020186