Genetic Association Study of Acetylcholinesterase (ACHE) and Butyrylcholinesterase (BCHE) Variants in Sudden Infant Death Syndrome (SIDS)


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
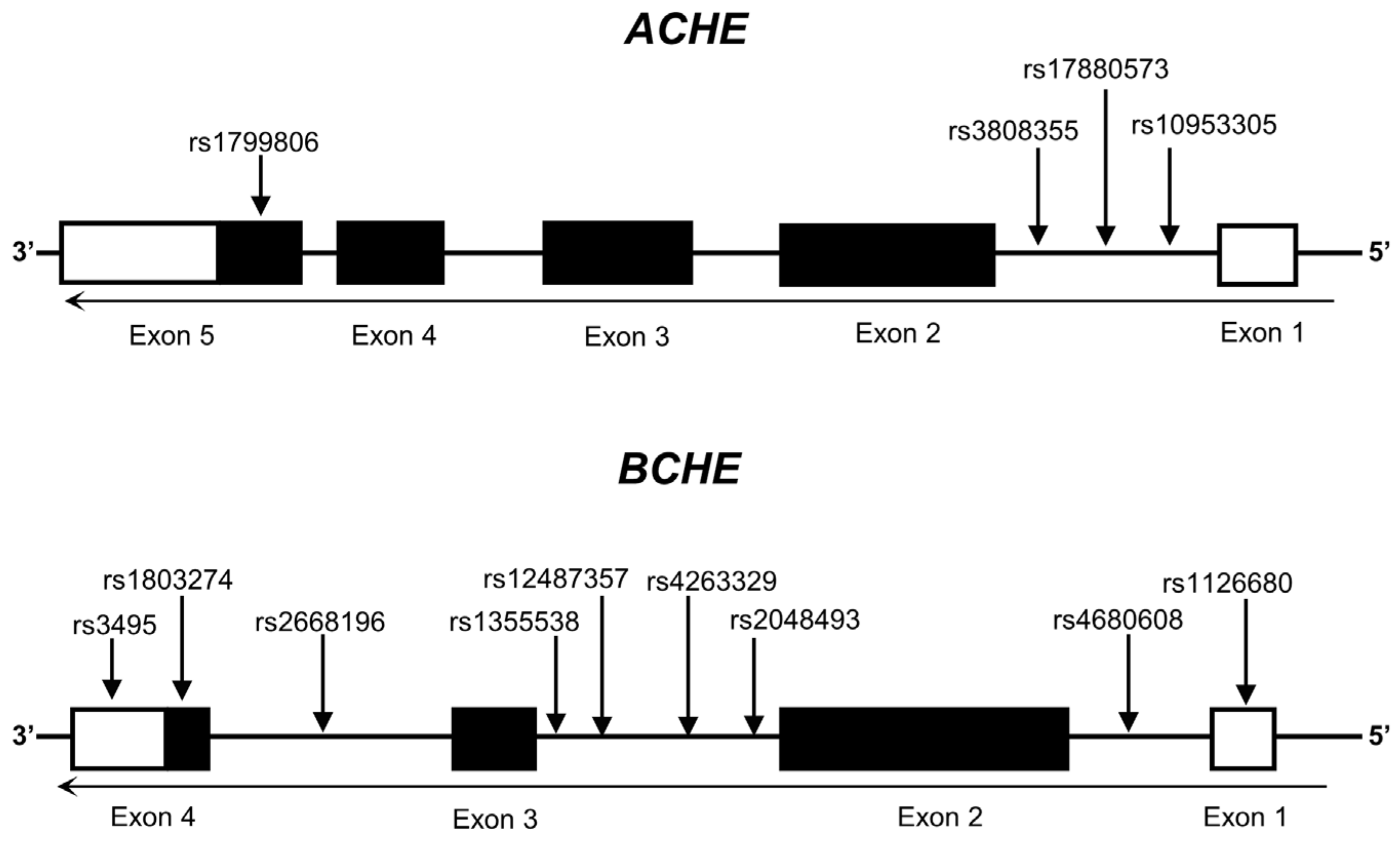
2.2. Selection of SNPs and Genotyping
2.3. Statistical Analyses
2.4. Expression Quantitative Trait Loci (eQTL) Analysis for Enzyme Expression
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krous, H.F.; Beckwith, J.B.; Byard, R.W.; Rognum, T.O.; Bajanowski, T.; Corey, T.; Cutz, E.; Hanzlick, R.; Keens, T.G.; Mitchell, E.A. Sudden infant death syndrome and unclassified sudden infant deaths: A definitional and diagnostic approach. Pediatrics 2004, 114, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Filiano, J.J.; Kinney, H.C. A perspective on neuropathologic findings in victims of the sudden infant death syndrome: The triple-risk model. Biol. Neonate 1994, 65, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Guntheroth, W.G.; Spiers, P.S. The triple risk hypotheses in sudden infant death syndrome. Pediatrics 2002, 110, e64. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Engelmann, T.A.; Preuss, V.; Hagemeier, L.; Radomsky, L.; Beushausen, K.; Keil, J.; Vennemann, B.; Falk, C.S.; Klintschar, M. Pulmonary immune profiling of SIDS: Impaired immune maturation and age-related cytokine imbalance. Pediatr. Res. 2023, 93, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Sarquella-Brugada, G.; Campuzano, O.; Cesar, S.; Iglesias, A.; Fernandez, A.; Brugada, J.; Brugada, R. Sudden infant death syndrome caused by cardiac arrhythmias: Only a matter of genes encoding ion channels? Int. J. Leg. Med. 2016, 130, 415–420. [Google Scholar] [CrossRef]
- Uzuntas, E.; Schürmann, P.; Rothämel, T.; Dörk, T.; Klintschar, M. Polymorphisms of the hypothalamic–pituitary–adrenal axis may lead to an inadequate response to stress and contribute to sudden infant death syndrome. Acta Paediatr. 2023, 112, 1478–1484. [Google Scholar] [CrossRef]
- Kinney, H.C.; Haynes, R.L. The Serotonin Brainstem Hypothesis for the Sudden Infant Death Syndrome. J. Neuropathol. Exp. Neurol. 2019, 78, 765–779. [Google Scholar] [CrossRef]
- Pfisterer, N.; Meyer-Bockenkamp, F.; Qu, D.; Preuss, V.; Rothämel, T.; Geisenberger, D.; Läer, K.; Vennemann, B.; Albers, A.; Engelmann, T.A.; et al. Sudden infant death syndrome revisited: Serotonin transporter gene, polymorphisms and promoter methylation. Pediatr. Res. 2022, 92, 694–699. [Google Scholar] [CrossRef]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef]
- Klinkenberg, I.; Sambeth, A.; Blokland, A. Acetylcholine and attention. Behav. Brain Res. 2011, 221, 430–442. [Google Scholar] [CrossRef]
- Teles-Grilo Ruivo, L.M.; Baker, K.L.; Conway, M.W.; Kinsley, P.J.; Gilmour, G.; Phillips, K.G.; Isaac, J.T.R.; Lowry, J.P.; Mellor, J.R. Coordinated Acetylcholine Release in Prefrontal Cortex and Hippocampus Is Associated with Arousal and Reward on Distinct Timescales. Cell Rep. 2017, 18, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.L.; Aitken, T.J.; Greenfield, V.Y.; Ostlund, S.B.; Wassum, K.M. Nucleus Accumbens Acetylcholine Receptors Modulate Dopamine and Motivation. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2016, 41, 2830–2838. [Google Scholar] [CrossRef] [PubMed]
- Rylett, R.J.; Schmidt, B.M. Regulation of the synthesis of acetylcholine. Prog. Brain Res. 1993, 98, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Sine, S.M. End-plate acetylcholine receptor: Structure, mechanism, pharmacology, and disease. Physiol. Rev. 2012, 92, 1189–1234. [Google Scholar] [CrossRef]
- Sarter, M.; Parikh, V. Choline transporters, cholinergic transmission and cognition. Nat. Rev. Neurosci. 2005, 6, 48–56. [Google Scholar] [CrossRef]
- Harrington, C.T.; Hafid, N.A.; Waters, K.A. Butyrylcholinesterase is a potential biomarker for Sudden Infant Death Syndrome. EBioMedicine 2022, 80, 104041. [Google Scholar] [CrossRef]
- Joda, M.; Waters, K.A.; Machaalani, R. Choline-acetyltransferase (ChAT) and acetylcholinesterase (AChE) in the human infant dorsal motor nucleus of the Vagus (DMNV), and alterations according to sudden infant death syndrome (SIDS) category. Neurobiol. Dis. 2023, 188, 106319. [Google Scholar] [CrossRef]
- Dick, A.; Ford, R. Cholinergic and oxidative stress mechanisms in sudden infant death syndrome. Acta Paediatr. 2009, 98, 1768–1775. [Google Scholar] [CrossRef]
- Kalaria, R.N.; Fiedler, C.; Hunsaker, J.C., 3rd; Sparks, D.L. Synaptic neurochemistry of human striatum during development: Changes in sudden infant death syndrome. J. Neurochem. 1993, 60, 2098–2105. [Google Scholar] [CrossRef]
- Sparks, D.L.; Hunsaker, J.C., 3rd. Sudden infant death syndrome: Altered aminergic-cholinergic synaptic markers in hypothalamus. J. Child Neurol. 1991, 6, 335–339. [Google Scholar] [CrossRef]
- Livolsi, A.; Niederhoffer, N.; Dali-Youcef, N.; Rambaud, C.; Olexa, C.; Mokni, W.; Gies, J.P.; Bousquet, P. Cardiac muscarinic receptor overexpression in sudden infant death syndrome. PLoS ONE 2010, 5, e9464. [Google Scholar] [CrossRef] [PubMed]
- Opdal, S.H.; Rognum, T.O. The sudden infant death syndrome gene: Does it exist? Pediatrics 2004, 114, e506–e512. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Chou, W.H.; Fang, C.P.; Liu, T.H.; Tsou, H.H.; Wang, Y.; Liu, Y.L. Serum Level and Activity of Butylcholinesterase: A Biomarker for Post-Stroke Dementia. J. Clin. Med. 2019, 8, 1778. [Google Scholar] [CrossRef]
- Lima, J.K.; Leite, N.; Turek, L.V.; Souza, R.L.; da Silva Timossi, L.; Osiecki, A.C.; Osiecki, R.; Furtado-Alle, L. 1914G variant of BCHE gene associated with enzyme activity, obesity and triglyceride levels. Gene 2013, 532, 24–26. [Google Scholar] [CrossRef]
- Jasiecki, J.; Żuk, M.; Krawczyńska, N.; Jońca, J.; Szczoczarz, A.; Lewandowski, K.; Waleron, K.; Wasąg, B. Haplotypes of butyrylcholinesterase K-variant and their influence on the enzyme activity. Chem.-Biol. Interact. 2019, 307, 154–157. [Google Scholar] [CrossRef]
- Qu, D.; Schürmann, P.; Rothämel, T.; Fleßner, J.; Rehberg, D.; Dörk, T.; Klintschar, M. Revisiting the association of sudden infant death syndrome (SIDS) with polymorphisms of NHE3 and IL13. Int. J. Leg. Med. 2023, 138, 743–749. [Google Scholar] [CrossRef]
- Qu, D.; Schürmann, P.; Rothämel, T.; Dörk, T.; Klintschar, M. Variants in genes encoding the SUR1-TRPM4 non-selective cation channel and sudden infant death syndrome (SIDS): Potentially increased risk for cerebral edema. Int. J. Leg. Med. 2022, 136, 1113–1120. [Google Scholar] [CrossRef]
- Sun, W.; Hu, Y. eQTL Mapping Using RNA-seq Data. Stat Biosci 2013, 5, 198–219. [Google Scholar] [CrossRef]
- Kahn, A.; Groswasser, J.; Franco, P.; Scaillet, S.; Sawaguchi, T.; Kelmanson, I.; Dan, B. Sudden infant deaths: Stress, arousal and SIDS. Early Hum. Dev. 2003, 75, S147–S166. [Google Scholar] [CrossRef]
- Rump, K.; Holtkamp, C.; Bergmann, L.; Nowak, H.; Unterberg, M.; Orlowski, J.; Thon, P.; Bazzi, Z.; Bazzi, M.; Adamzik, M.; et al. Midazolam impacts acetyl-And butyrylcholinesterase genes: An epigenetic explanation for postoperative delirium? PLoS ONE 2022, 17, e0271119. [Google Scholar] [CrossRef]
- Santarpia, L.; Grandone, I.; Contaldo, F.; Pasanisi, F. Butyrylcholinesterase as a prognostic marker: A review of the literature. J. Cachexia Sarcopenia Muscle 2013, 4, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Karami, A.; Darreh-Shori, T.; Schultzberg, M.; Eriksdotter, M. CSF and Plasma Cholinergic Markers in Patients with Cognitive Impairment. Front. Aging Neurosci. 2021, 13, 704583. [Google Scholar] [CrossRef] [PubMed]
- Ogunkeye, O.O.; Roluga, A.I. Serum cholinesterase activity helps to distinguish between liver disease and non-liver disease aberration in liver function tests. Pathophysiol. Off. J. Int. Soc. Pathophysiol. 2006, 13, 91–93. [Google Scholar] [CrossRef]
- Lampón, N.; Hermida-Cadahia, E.F.; Riveiro, A.; Tutor, J.C. Association between butyrylcholinesterase activity and low-grade systemic inflammation. Ann. Hepatol. 2012, 11, 356–363. [Google Scholar] [CrossRef]
- Zugno, A.I.; Fraga, D.B.; De Luca, R.D.; Ghedim, F.V.; Deroza, P.F.; Cipriano, A.L.; Oliveira, M.B.; Heylmann, A.S.; Budni, J.; Souza, R.P.; et al. Chronic exposure to cigarette smoke during gestation results in altered cholinesterase enzyme activity and behavioral deficits in adult rat offspring: Potential relevance to schizophrenia. J. Psychiatr. Res. 2013, 47, 740–746. [Google Scholar] [CrossRef]
- MacFarlane, P.M. Inflammation in sudden infant death syndrome. Pediatr. Res. 2024, 95, 885–886. [Google Scholar] [CrossRef]
- Qu, D.; Preuss, V.; Hagemeier, L.; Radomsky, L.; Beushausen, K.; Keil, J.; Nora, S.; Vennemann, B.; Falk, C.S.; Klintschar, M. Age-related cytokine imbalance in the thymus in sudden infant death syndrome (SIDS). Pediatr. Res. 2024, 95, 949–958. [Google Scholar] [CrossRef]
- Mychaleckyj, J.C.; Normeshie, C.; Keene, K.L.; Hauck, F.R. Organ weights and length anthropometry measures at autopsy for sudden infant death syndrome cases and other infant deaths in the Chicago infant mortality study. Am. J. Hum. Biol. 2024, 36, e24126. [Google Scholar] [CrossRef]
- Goldwater, P.N. Current SIDS research: Time to resolve conflicting research hypotheses and collaborate. Pediatr. Res. 2023, 94, 1273–1277. [Google Scholar] [CrossRef]
- Vivekanandarajah, A.; Nelson, M.E.; Kinney, H.C.; Elliott, A.J.; Folkerth, R.D.; Tran, H.; Cotton, J.; Jacobs, P.; Minter, M.; McMillan, K.; et al. Nicotinic Receptors in the Brainstem Ascending Arousal System in SIDS with Analysis of Pre-natal Exposures to Maternal Smoking and Alcohol in High-Risk Populations of the Safe Passage Study. Front. Neurol. 2021, 12, 636668. [Google Scholar] [CrossRef]
- Robles, A.; Michael, M.; McCallum, R. Pseudocholinesterase Deficiency: What the Proceduralist Needs to Know. Am. J. Med. Sci. 2019, 357, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Soliday, F.K.; Conley, Y.P.; Henker, R. Pseudocholinesterase deficiency: A comprehensive review of genetic, acquired, and drug influences. AANA J. 2010, 78, 313–320. [Google Scholar]


{kind=link}
{kind=link}
| SNP | Alleles | Genome Position (GRCh38) | Gene: Consequence | Reported Association with Other Diseases? | Altered Gene Expression in eQTL |
|---|---|---|---|---|---|
| rs3495 | T > C | chr3:165773193 | BCHE: Non-Coding Transcript Variant | Yes | BCHE |
| rs1126680 | C > T | chr3:165837337 | BCHE: Non-Coding Transcript Variant | Yes | BCHE |
| rs1803274 | C > T | chr3:165773492 | BCHE: Missense Variant | Yes | BCHE |
| rs1355538 | G > A | chr3:165787389 | BCHE: Intron Variant | Yes | BCHE |
| rs2048493 | C > G | chr3:165826514 | BCHE: Intron Variant | Yes | BCHE |
| rs2668196 | T > A | chr3:165784921 | BCHE: Intron Variant | Yes | BCHE |
| rs4263329 | A > G | chr3:165821822 | BCHE: Intron Variant | Yes | BCHE |
| rs4680608 | T > C | chr3:165835858 | BCHE: Intron Variant | Yes | BCHE |
| rs12487357 | G > C | chr3:165804028 | BCHE: Intron Variant | Yes | None |
| rs1799806 | G > C | chr7:100891037 | ACHE: Missense Variant | Yes | ACHE |
| rs3808355 | C > T | chr7:100894677 | ACHE: Intron Variant | No | ACHE |
| rs17880573 | C > T | chr7:100894791 | ACHE: Intron Variant | No | ACHE |
| rs10953305 | G > A | chr7:100894947 | ACHE: Intron Variant | No | ACHE |
| Gene | SNP | Alleles | Genotype Distribution (XX:XY:YY) | p Value | |||
|---|---|---|---|---|---|---|---|
| SIDS | Control | Additive | Dominant | Recessive | |||
| BCHE | rs3495 | T > C | 109:71:21 | 188:123:27 | 0.625 | 0.753 | 0.351 |
| rs1126680 | C > T | 149:45:07 | 239:91:05 | 0.938 | 0.793 | 0.900 | |
| rs1803274 | C > T | 133:60:08 | 231:92:15 | 0.795 | 0.635 | 0.799 | |
| rs1355538 | G > A | 73:83:44 | 114:162:62 | 0.321 | 0.514 | 0.314 | |
| rs2048493 | C > G | 88:84:29 | 153:142:43 | 0.844 | 0.737 | 0.602 | |
| rs2668196 | T > A | 135:61:5 | 218:107:13 | 0.637 | 0.529 | 0.466 | |
| rs4263329 | A > G | 168:29:04 | 284:51:02 | 0.324 | 0.832 | 0.203 | |
| rs4680608 | T > C | 136:51:14 | 233:93:12 | 0.204 | 0.774 | 0.095 | |
| rs12487357 | G > C | 176:23:02 | 284:53:01 | 0.235 | 0.314 | 0.559 | |
| ACHE | rs1799806 | G > C | Deviation from HWE | - | - | - | |
| rs3808355 | C > T | Low call rates | - | - | - | ||
| rs17880573 | C > T | 149:45:07 | 239:91:05 | 0.181 | 0.549 | 0.132 | |
| rs10953305 | G > A | 46:102:51 | 91:157:89 | 0.513 | 0.357 | 0.919 | |
| Stratum | Gene | SNP | Genotype Distribution | Additive (Y vs. X Alleles) | Dominant (YY/XY vs. XX Genotypes) | Recessive (YY vs. XX/XY Genotypes) | |||
|---|---|---|---|---|---|---|---|---|---|
| Case No. in SIDS (XX:XY:YY) | OR (95% CI) * | p Value * | OR (95% CI) | p value | OR (95% CI) | p Value | |||
| Female | BCHE | rs4680608 | 49:24:10 | 0.609 (0.416~0.893) | 0.010 (C) | 1.540 (0.939~2.524) | 0.086 | 3.717 (1.548~8.929) | 0.004 (CC) |
| Female | rs1355538 | 23:31:28 | 1.501 (1.075~2.096) | 0.016 (A) | 1.306 (0.767~2.222) | 0.325 | 2.309 (1.355~3.937) | 0.002 (AA) | |
| Female | rs3495 | 36:32:15 | 1.610 (1.140~2.272) | 0.006 (C) | 1.636 (1.008~2.656) | 0.045 (CC/TC) | 2.538 (1.282~5.025) | 0.006 (CC) | |
| Female | rs1803274 | 46:33:04 | 0.683 (0.460~1.015) | 0.058 | 1.736 (1.064~2.834) | 0.026 (TT/CT) | 1.091 (0.352~3.378) | 1.000 | |
| Female | rs2048493 | 28:35:20 | 1.562 (1.118~2.178) | 0.008 (G) | 1.625 (0.982~2.686) | 0.057 | 2.179 (1.2~3.953) | 0.009 (GG) | |
| Age 2–4 months | rs3495 | 39:24:12 | 0.774 (0.537~1.115) | 0.168 | 1.157 (0.701~1.910) | 0.569 | 2.193 (1.055~4.566) | 0.032 (CC) | |
| Age 2–4 months | rs4263329 | 62:10:03 | 0.757 (0.428~1.338) | 0.337 | 1.124 (0.577~2.187) | 0.732 | 6.993 (1.145~41.667) | 0.015 (GG) | |
| Autumn | rs2048493 | 15:17:11 | 1.630 (1.049~2.564) | 0.041 (G) | 1.544 (0.806~2.906) | 0.254 | 2.358 (1.065~4.950) | 0.034 (GG) | |
| Autumn | rs4680608 | 29:09:05 | 1.248 (0.737~2.105) | 0.465 | 1.071 (0.544~2.093) | 0.862 | 3.575 (1.328~10.820) | 0.032 (CC) | |
| Autumn | rs12487357 | 35:06:02 | 1.486 (0.730~3.048) | 0.303 | 1.202 (0.558~2.710) | 0.662 | 16.441 (1.852~238.6) | 0.035 (CC) | |
| Summer | ACHE | rs10953305 | 09:18:03 | 1.482 (0.854~2.500) | 0.178 | 1.159 (0.495~2.544) | 0.676 | 0.310 (0.097~0.984) | 0.049 (AA) |
| Haplotypes * | Frequencies in Controls # | Frequencies in Overall SIDS | p Value | Frequencies in Male SIDS | p Value | Frequencies in Female SIDS | p Value | |
|---|---|---|---|---|---|---|---|---|
| rs17880573 | rs10953305 | |||||||
| C | G | 49.9% (337:339) | 48.7% (196:206) | 0.707 | 47.8% (113:123) | 0.586 | 50% (83:83) | 0.987 |
| C | A | 34.9% (236:440) | 36.6% (147:255) | 0.581 | 37.8% (89:147) | 0.438 | 35% (58:108) | 0.994 |
| T | A | 14.8% (100:576) | 14.6% (59:343) | 0.935 | 14.3% (34:202) | 0.870 | 14.9% (25:141) | 0.953 |
| Haplotypes * | Frequencies in Controls # | Frequencies in SIDS | p Value | Frequencies in Male SIDS | p Value | Frequencies in Female SIDS | p Value | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| rs3495 | rs1803274 | rs1355538 | rs2048493 | rs1126680 | |||||||
| T | C | G | C | C | 57.3% (388:288) | 56.1% (226:176) | 0.707 | 63.1% (149:87) | 0.121 | 46.3% (77:89) | 0.010 |
| C | T | A | G | C | 10% (67:609) | 11.9% (48:354) | 0.316 | 9.7% (23:213) | 0.904 | 15.1% (25:141) | 0.058 |
| C | C | A | G | C | 8% (54:622) | 8.9% (36:366) | 0.598 | 8% (19:217) | 0.920 | 12.5% (21: 145) | 0.065 |
| T | C | A | C | C | 7.8% (56:620) | 8% (32:370) | 0.935 | 6.4% (15:221) | 0.338 | 7.9% (13:152) | 0.847 |
| T | C | A | G | C | 8.3% (53:623) | 7% (28:374) | 0.432 | 6.3% (15:221) | 0.411 | 7.9% (13:153) | 0.974 |
| C | T | A | G | T | 7.1% (48:628) | 6.7% (27:375) | 0.811 | 5.1% (12:224) | 0.282 | 9% (15:151) | 0.395 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, D.; Schürmann, P.; Rothämel, T.; Dörk, T.; Klintschar, M. Genetic Association Study of Acetylcholinesterase (ACHE) and Butyrylcholinesterase (BCHE) Variants in Sudden Infant Death Syndrome (SIDS). Genes 2024, 15, 1656. https://doi.org/10.3390/genes15121656
Qu D, Schürmann P, Rothämel T, Dörk T, Klintschar M. Genetic Association Study of Acetylcholinesterase (ACHE) and Butyrylcholinesterase (BCHE) Variants in Sudden Infant Death Syndrome (SIDS). Genes. 2024; 15(12):1656. https://doi.org/10.3390/genes15121656
Chicago/Turabian StyleQu, Dong, Peter Schürmann, Thomas Rothämel, Thilo Dörk, and Michael Klintschar. 2024. "Genetic Association Study of Acetylcholinesterase (ACHE) and Butyrylcholinesterase (BCHE) Variants in Sudden Infant Death Syndrome (SIDS)" Genes 15, no. 12: 1656. https://doi.org/10.3390/genes15121656
APA StyleQu, D., Schürmann, P., Rothämel, T., Dörk, T., & Klintschar, M. (2024). Genetic Association Study of Acetylcholinesterase (ACHE) and Butyrylcholinesterase (BCHE) Variants in Sudden Infant Death Syndrome (SIDS). Genes, 15(12), 1656. https://doi.org/10.3390/genes15121656