
Molecular Markers Help with Breeding for Agronomic Traits of Spring Wheat in Kazakhstan and Siberia

 , , , , , ,
, , , , , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
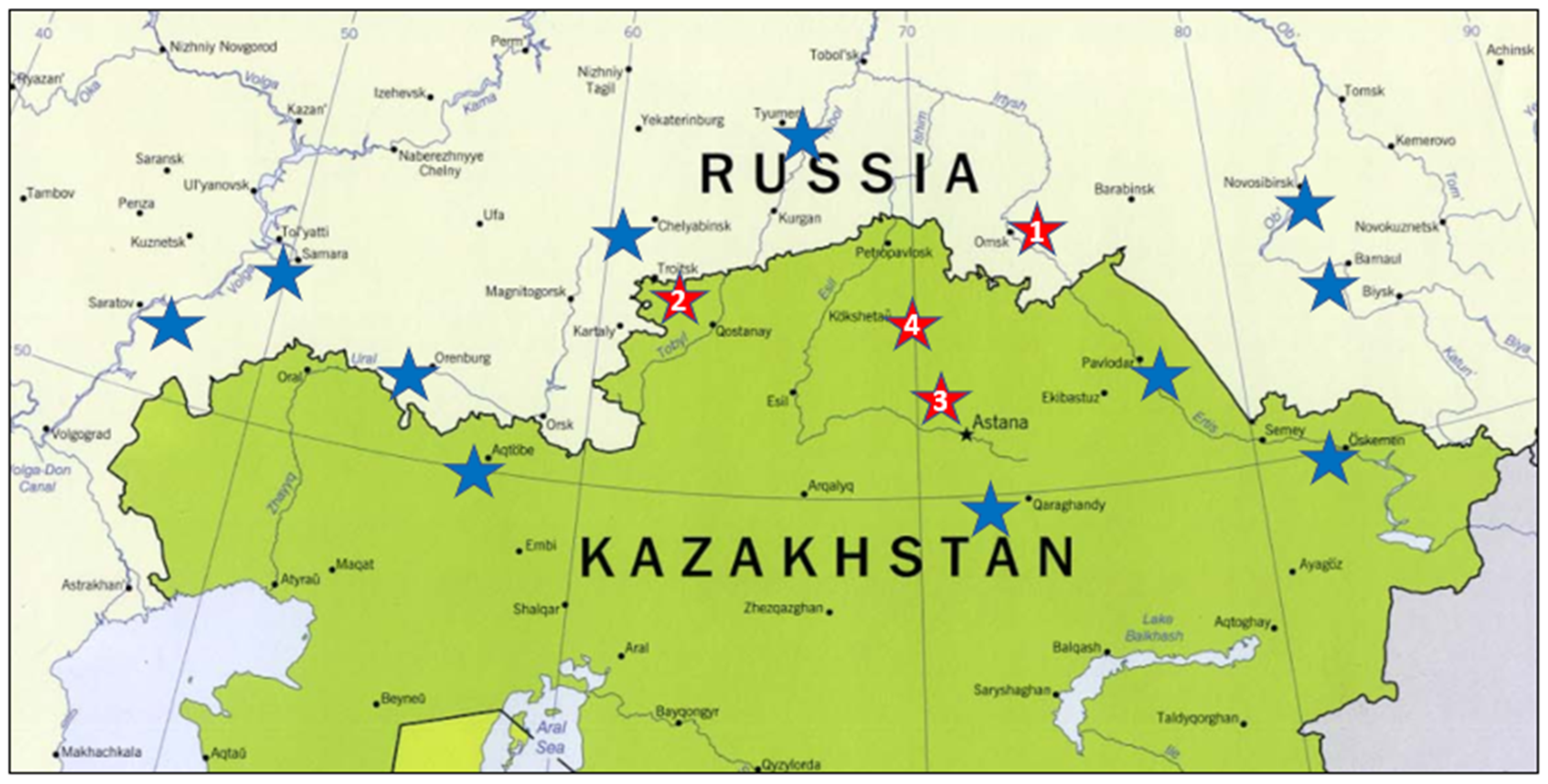
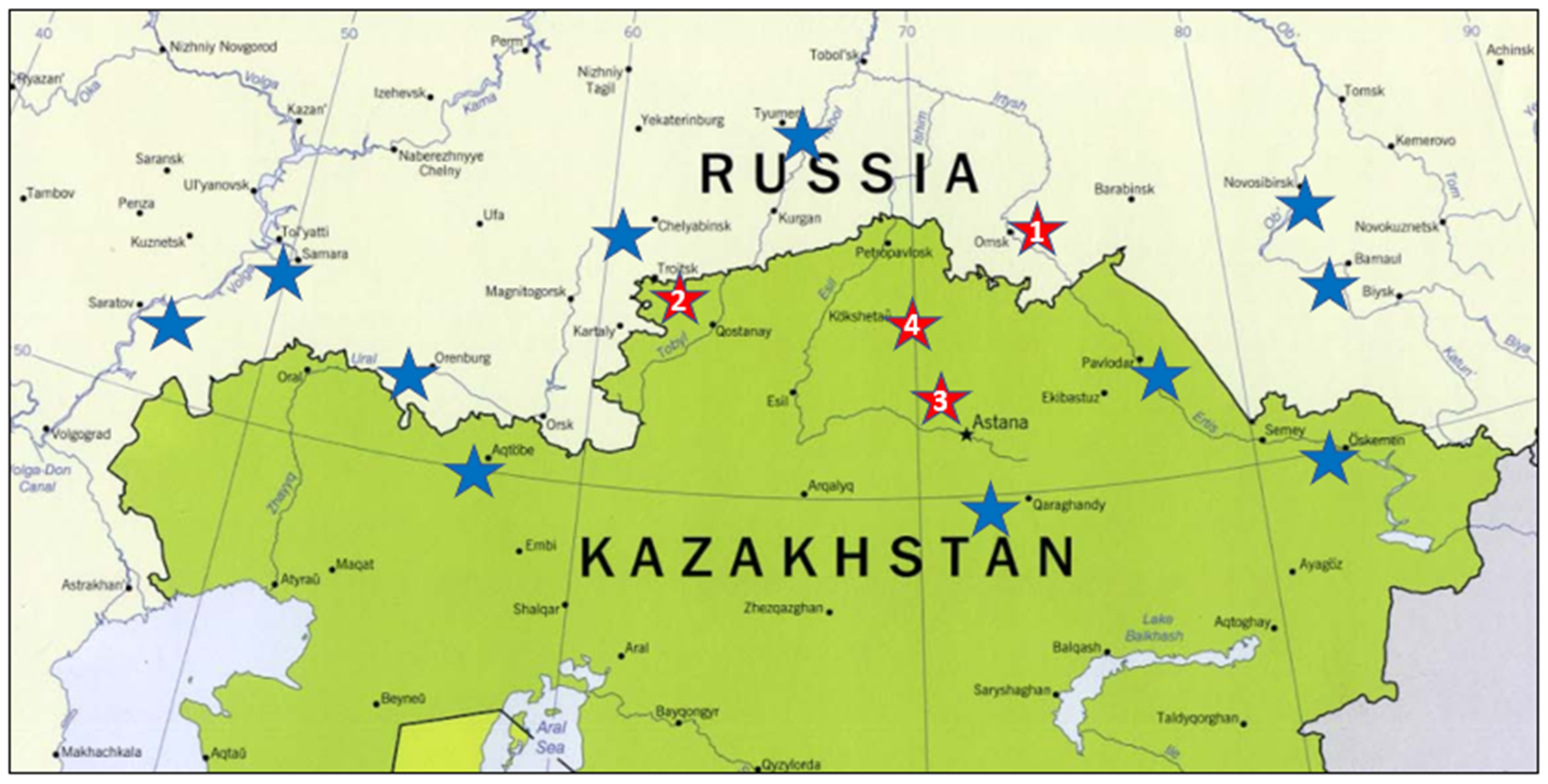
2.1. Material
2.2. Yield Trials
2.3. Genotyping
3. Results
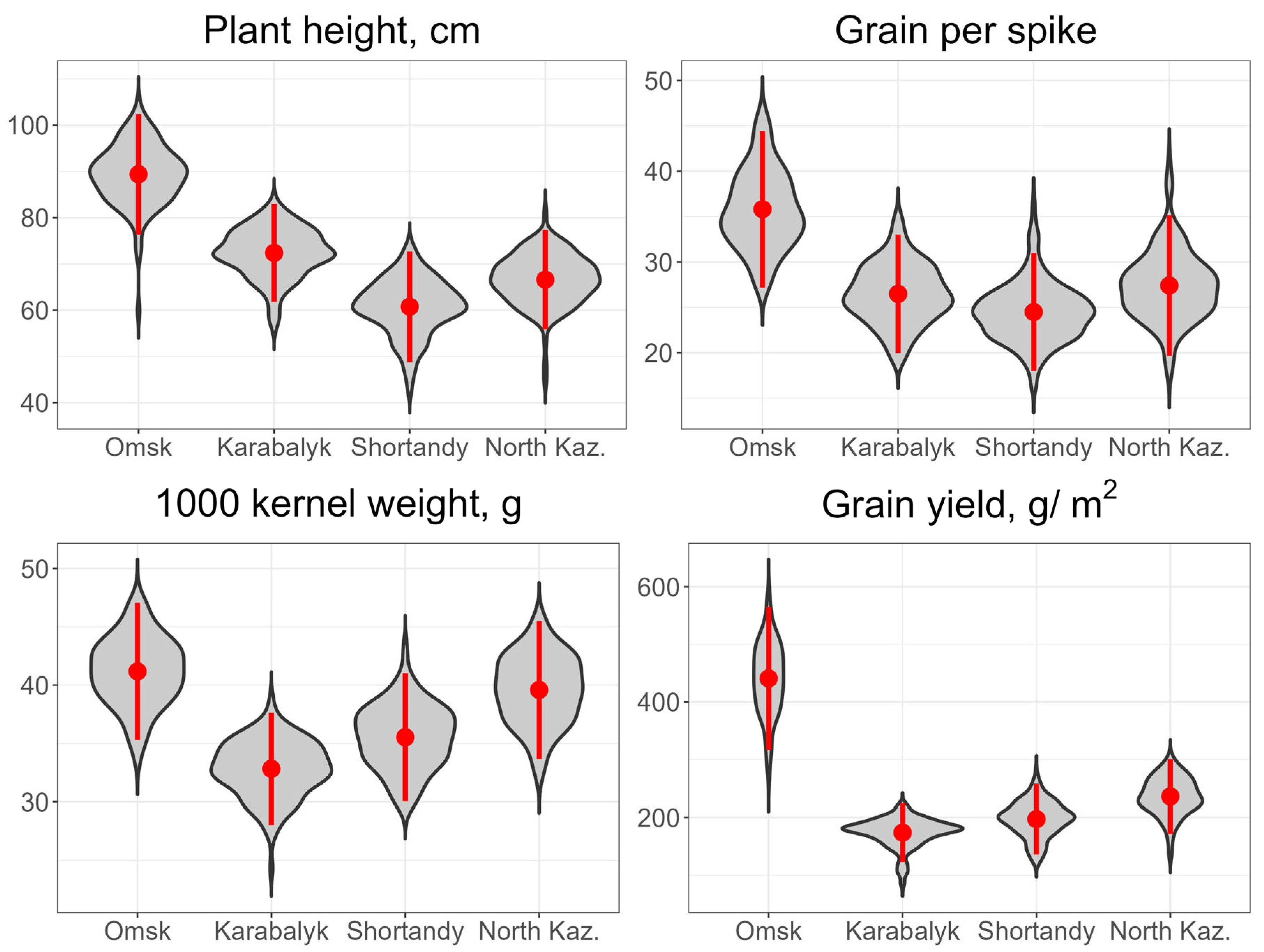
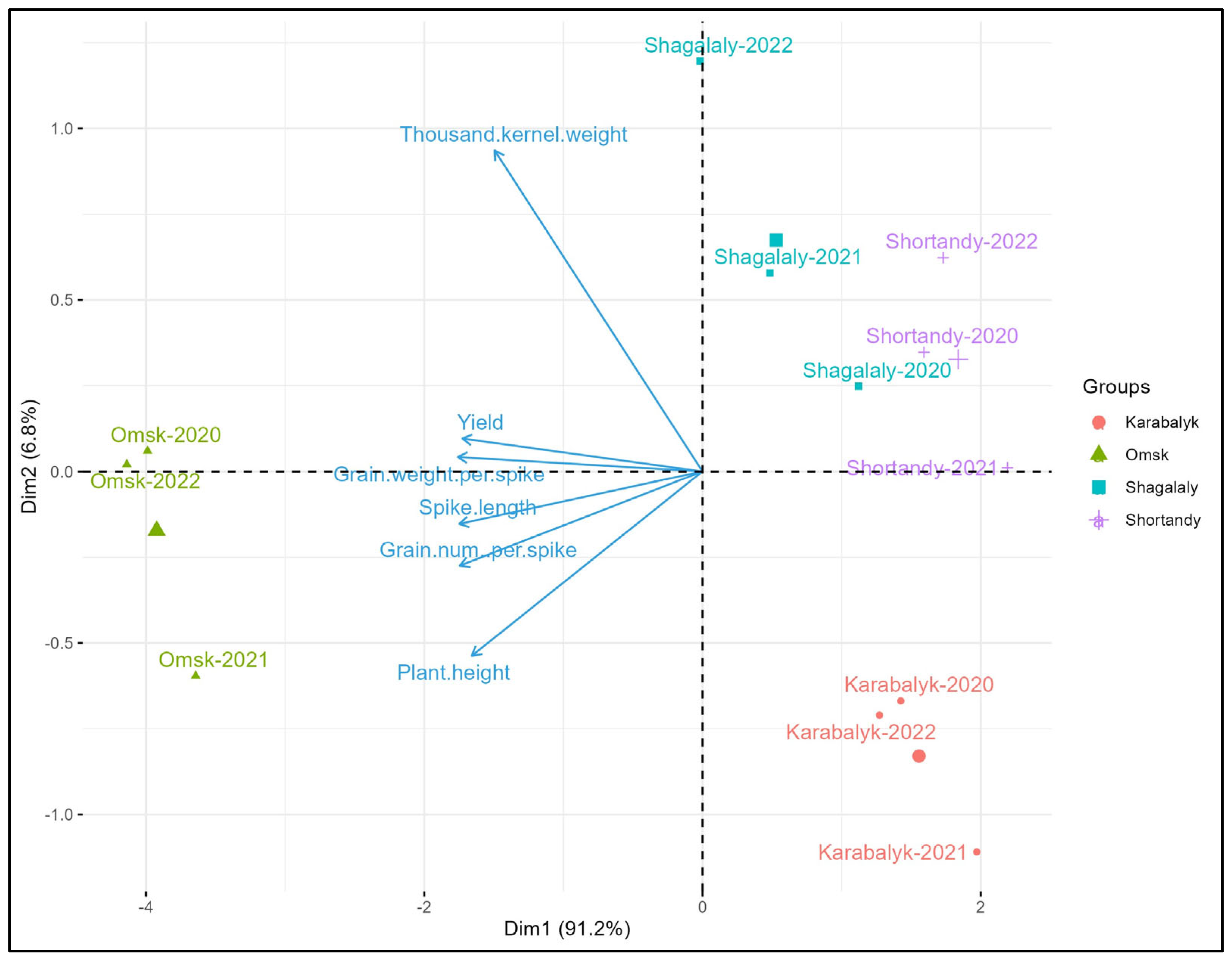
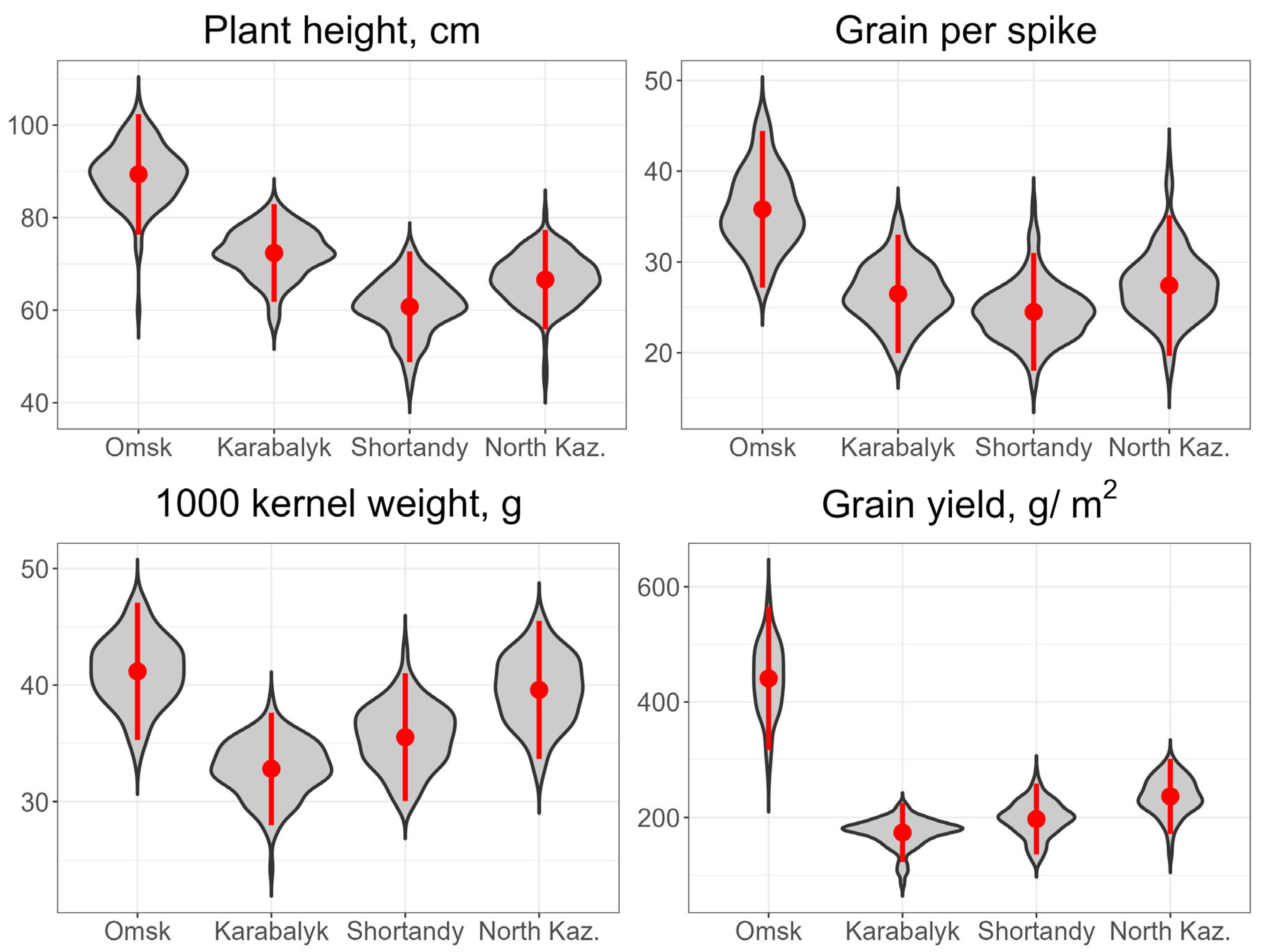
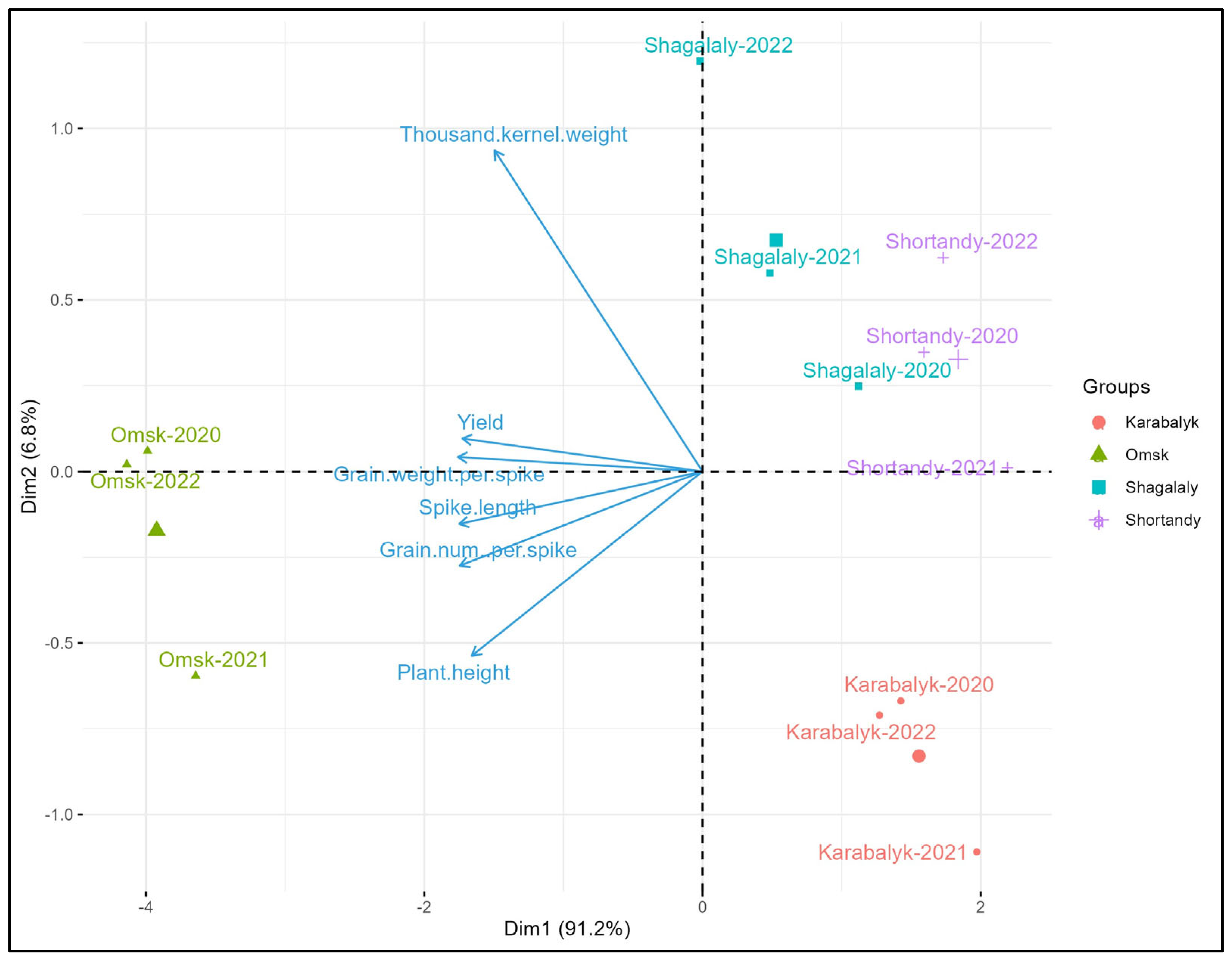
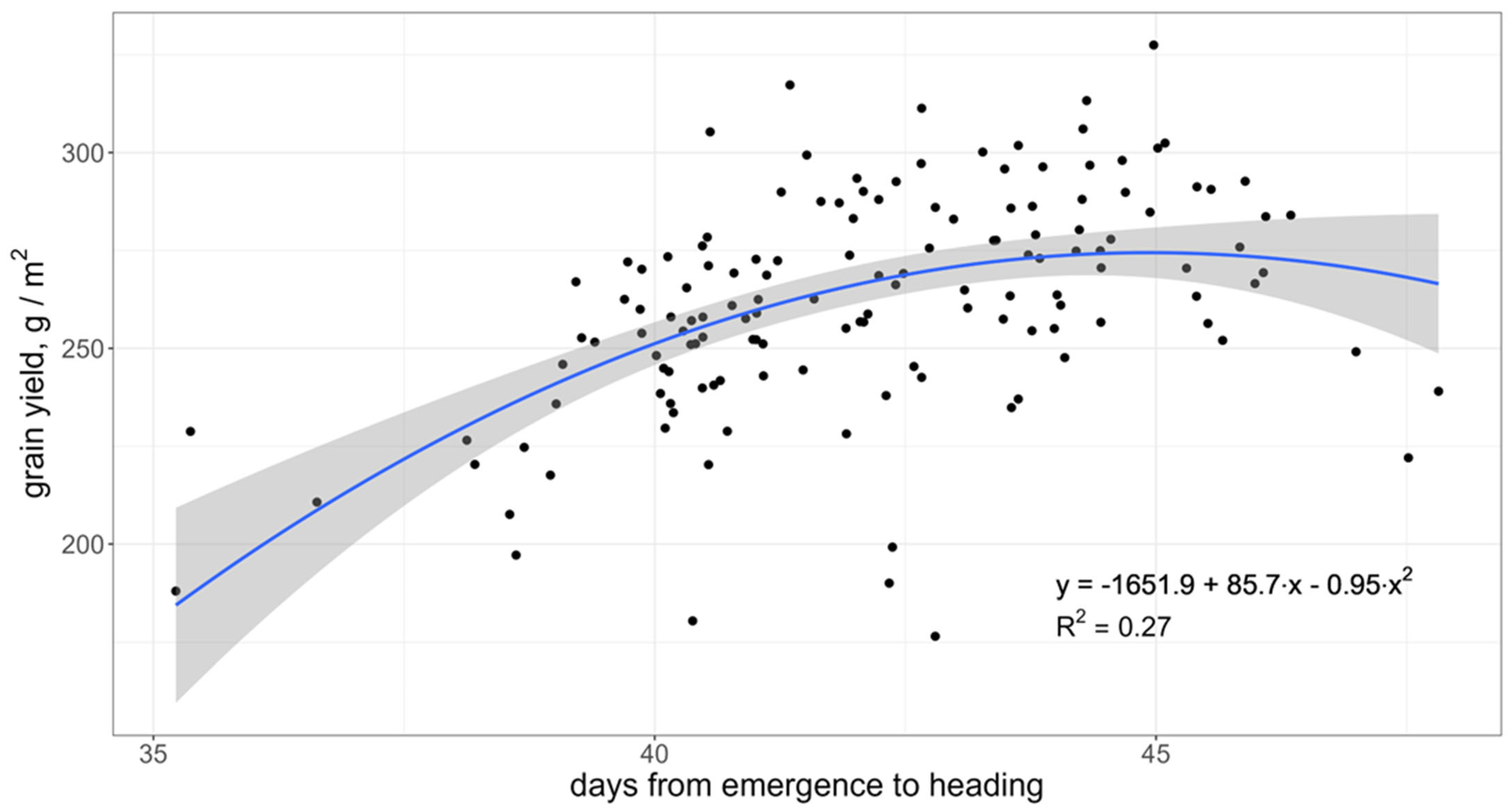
3.1. Grain Yield and Agronomic Traits across the Study Sites and Years
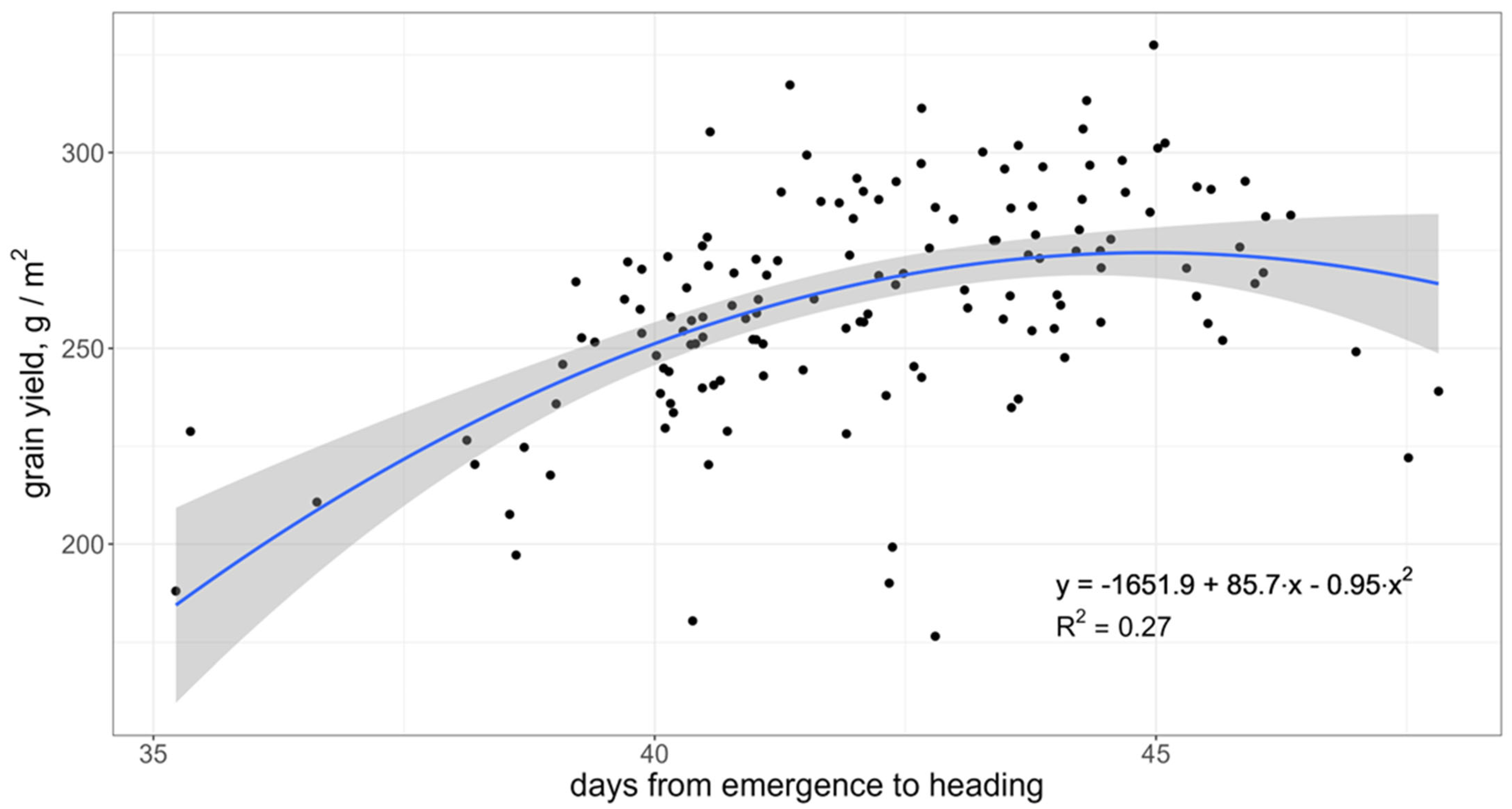
3.2. Association of Grain Yield with Adaptation Traits and Yield Components
3.3. Effects of Molecular Markers on Agronomic Traits
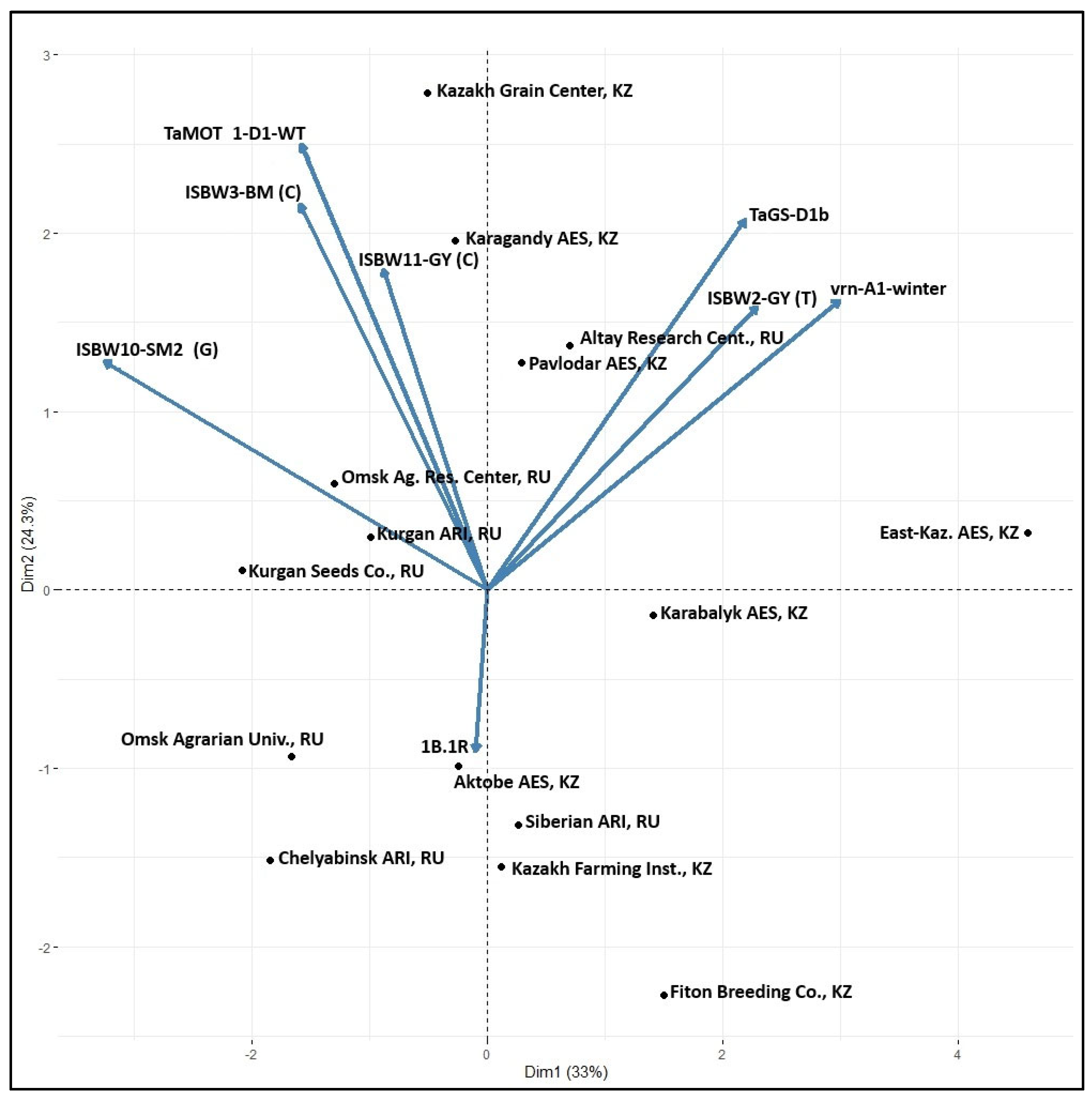
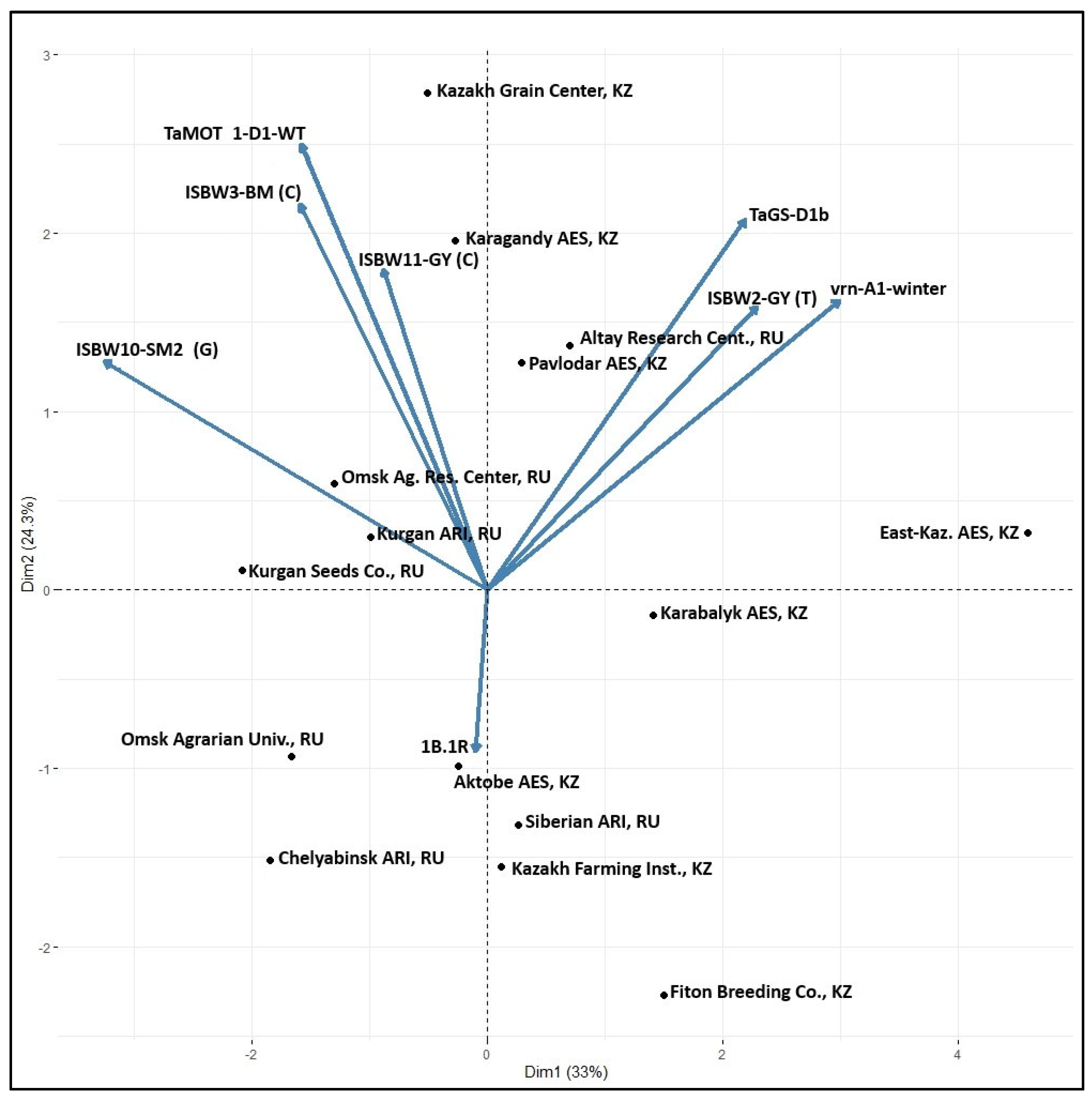
3.4. Markers Distribution across the Breeding Programs
3.5. Identification of Superior Germplasm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karatayev, M.; Clarke, M.; Salnikov, V.; Bekseitova, R.; Nizamova, M. Monitoring climate change, drought conditions and wheat production in Eurasia: The case study of Kazakhstan. Heliyon 2022, 8, e08660. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.P. Scientific support to plant breeding and seed production in Siberia in the XXI century. Vavilov J. Genet. Breed. 2021, 25, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Babar, H.; Akpınar, B.A.; Alaux, M.; Algharib, A.M.; Sehgal, D.; Ali, Z.; Aradottir, G.I.; Batley, J.; Bellec, A.; Bentley, A.R.; et al. Capturing wheat phenotypes at the genome level. Front. Plant. Sci. 2022, 851079. [Google Scholar]
- Song, L.; Wang, R.; Yang, X.; Zhang, A.; Liu, D. Molecular markers and their applications in marker-assisted selection (MAS) in bread wheat (Triticum aestivum L.). Agriculture 2023, 13, 642. [Google Scholar] [CrossRef]
- Sun, C.; Hu, H.; Cheng, Y.; Yang, X.; Qiao, Q.; Wang, C.; Zhang, L.; Chen, D.; Zhao, S.; Dong, Z.; et al. Genomics-assisted breeding: The next-generation wheat breeding era. Plant Breed. 2023, 142, 259–268. [Google Scholar] [CrossRef]
- Genievskaya, Y.; Abugalieva, S.; Rsaliyev, A.; Yskakova, G.; Turuspekov, Y. QTL mapping for seedling and adult plant resistance to leaf and stem rusts in Pamyati Azieva × Paragon mapping population of bread wheat. Agronomy 2020, 10, 1285. [Google Scholar] [CrossRef]
- Kelbin, V.N.; Skolotneva, E.S.; Shamanin, V.P.; Salina, E.A. Diversity of stem rust resistance in modern Siberian bread wheat (Triticum aestivum) germplasm. Plant Breed. 2022, 141, 194–203. [Google Scholar] [CrossRef]
- Bhatta, M.; Shamanin, V.P.; Shepelev, S.S.; Baenziger, P.S.; Pozherukova, V.; Pototskaya, I.; Morgounov, A.I. Marker-trait associations for enhancing agronomic performance, disease resistance, and grain quality in synthetic and bread wheat accessions in western Siberia. G3 Genes Genomes Genet. 2019, 9, 4209–4222. [Google Scholar] [CrossRef]
- Amalova, A.; Abugalieva, S.; Babkenov, A.; Babkenova, S.; Turuspekov, Y. Genome-wide association study of yield components in spring wheat collection harvested under two water regimes in Northern Kazakhstan. Peer J. 2021, 9, e11857. [Google Scholar] [CrossRef]
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012. [Google Scholar]
- Smith, A.B.; Cullis, B.R.; Thompson, R. The analysis of crop cultivar breeding and evaluation trials: An overview of current mixed model approaches. J. Agric. Sci. 2005, 143, 449–462. [Google Scholar] [CrossRef]
- Dreisigacker, S.; Sehgal, D.; Reyes Jaimez, A.E. CIMMYT Wheat Molecular Genetics: Laboratory Protocols and Applications to Wheat Breeding; CIMMYT: Mexico City, Mexico, 2016; pp. 1–154. [Google Scholar]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef]
- Chai, L.; Xin, M.; Dong, C.; Chen, Z.; Zhai, H.; Zhuang, J.; Cheng, X.; Wang, N.; Geng, J.; Wang, X.; et al. A natural variation in Ribonuclease H-like gene underlies Rht8 to confer “Green Revolution” trait in wheat. Mol. Plant 2022, 15, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Sherman, J.D.; Yan, L.; Talbert, L.; Dubcovsky, J. A PCR marker for growth habit in common wheat based on allelic variation at the VRN-A1 gene. Crop. Sci. 2004, 44, 1832–1838. [Google Scholar] [CrossRef]
- Alipour, H.; Abdi, H.; Rahimi, Y.; Bihamta, M.R. Genotype-by-year interaction for grain yield of Iranian wheat cultivars and its interpretation using Vrn and Ppd functional markers and environmental covariables. Cereal Res. Commun. 2021, 49, 681–690. [Google Scholar] [CrossRef]
- Efremova, T.T.; Chumanova, E.V.; Trubacheeva, N.V.; Arbuzova, V.S.; Belan, I.A.; Pershina, L.A. Prevalence of VRN1 locus alleles among spring common wheat cultivars cultivated in Western Siberia. Russ. J. Genet. 2016, 52, 146–153. [Google Scholar] [CrossRef]
- Zikhali, M.; Wingen, L.U.; Griffiths, S. Delimitation of the Earliness per se D1 (Eps-D1) flowering gene to a subtelomeric chromosomal deletion in bread wheat (Triticum aestivum). J. Exp. Botany 2016, 67, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Askhadullin, D.F.; Askhadullin, D.F.; Vasilova, N.Z.; Brykova, A.N. Earliness of spring bread wheat accessions under the conditions of the Middle Volga Region. Procd. Appl. Botany Genet. Breed. 2022, 183, 66–75. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.A.; Garkava-Gustavsson, L.; Åhman, I. A systematic review of rye (Secale cereale L.) as a source of resistance to pathogens and pests in wheat (Triticum aestivum L.). Hereditas 2017, 154, 14. [Google Scholar] [CrossRef]
- Rajaram, S. Prospects and promise of wheat breeding in the 21st century. Euphytica 2001, 119, 3–15. [Google Scholar] [CrossRef]
- Villareal, R.L.; Rajaram, S.; Mujeeb-Kazi, A.; Del Toro, E. The effect of chromosome 1B/1R translocation on the yield potential of certain spring wheats (Triticum aestivum L.). Plant Breed. 1991, 106, 77–81. [Google Scholar] [CrossRef]
- Moskal, K.; Kowalik, S.; Podyma, W.; Łapiński, B.; Boczkowska, M. The pros and cons of rye chromatin introgression into wheat genome. Agronomy 2021, 11, 456. [Google Scholar] [CrossRef]
- Korobkova, V.A.; Bespalova, L.A.; Yanovsky, A.S.; Chernook, A.G.; Kroupin, P.Y.; Arkhipov, A.V.; Yurkina, A.I.; Nazarova, L.A.; Mudrova, A.A.; Voropaeva, A.D.; et al. Permanent spreading of 1RS.1AL and 1RS.1BL translocations in modern wheat breeding. Plants 2023, 12, 1205. [Google Scholar] [CrossRef] [PubMed]
- Shamanin, V.; Salina, E.; Wanyera, R.; Zelenskiy, Y.; Olivera, P.; Morgounov, A. Genetic diversity of spring wheat from Kazakhstan and Russia for resistance to stem rust Ug99. Euphytica 2016, 212, 287–296. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, Y.-L.; Gao, L.-F.; Zhao, G.-Y.; Zhuo, R.-H.; Zhang, B.-S.; Jia, J.-Z. TaCK6-D1, the ortholog of rice OsCKX2, is associated with grain weight in hexaploid wheat. New Phytol. 2012, 195, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Zanke, C.; Ling, J.; Plieske, J.; Kollers, S.; Ebmeyer, E.; Korzun, V.; Argillier, O.; Stiewe, G.; Hinze, M.; Neumann, F.; et al. Analysis of main effect QTL for thousand grain weight in European winter wheat (Triticum aestivum L.) by genome-wide association mapping. Front. Plant Sci. 2015, 6, 644. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, D.; Mondal, S.; Guzman, C.G.; Franco, C.; Singh, R.; Dreisigacker, S. Validation of candidate gene-based markers and identification of novel loci for thousand-grain weight in spring bread wheat. Front. Plant Sci. 2019, 10, 01189. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Ali, M.; Shahid, M.; Arif, A.; Waheed, M.Q.; Xia, X.; Trethowan, R.; Tester, M.; Poland, J.; Ogbonnaya, F.C.; et al. Genetic networks underlying salinity tolerance in wheat uncovered with genome-wide analyses and selective sweeps. Theor. Appl. Genet. 2022, 135, 2925–2941. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, W.; Suleiman, S.; Tahir, I.; Sanchez-Garcia, M.; Jighly, A.; Hagras, A.; Thabet, S.; Baum, M. Heat-tolerant QTLs associated with grain yield and its components in spring bread wheat under heat-stressed environments of Sudan and Egypt. Crop. Sci. 2019, 59, 199–211. [Google Scholar] [CrossRef]
- Alemu, A.; Suliman, S.; Hagras, A.; Thabet, S.; Al-Abdallat, A.; Abdelmula, A.A.; Tadesse, W. Multi-model genome-wide association and genomic prediction analysis of 16 agronomic, physiological and quality related traits in ICARDA spring wheat. Euphytica 2021, 217, 205. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Testing Site | Rainfall May–August, mm | Average Temperature in July, °C | ||||||
|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2022 | Long-Term | 2020 | 2021 | 2022 | Long-Term | |
| Omsk, Omsk region | 150 | 90 | 217 | 207 | 22.1 | 21.1 | 20.5 | 19.2 |
| Karabalyk, Kostanay region | 118 | 89 | 90 | 179 | 25.2 | 22.1 | 23.4 | 20.8 |
| Shortandy, Akmola region | 124 | 88 | 100 | 136 | 17.7 | 19.5 | 19.5 | 18.5 |
| Shagalaly, North Kazakhstan | - | 131 | 179 | 193 | - | 20.8 | 21.2 | 20.0 |
| Site | Year | Coefficients of Correlation between the Grain Yield and: | ||||
|---|---|---|---|---|---|---|
| Plant Height | Spike Length | Grains per Spike | 1000 KW | Grain Weight per Spike | ||
| Karabalyk | 2020 | 0.29 *** | 0.01 | 0.40 *** | 0.15 | 0.28 *** |
| 2021 | - | 0.03 | 0.39 *** | 0.24 *** | 0.44 *** | |
| 2022 | 0.29 *** | −0.04 | 0.35 *** | 0.19 ** | 0.23 *** | |
| Omsk | 2020 | 0.27 *** | 0.19 ** | 0.40 *** | 0.24 *** | 0.41 *** |
| 2021 | 0.17 * | 0.14 | 0.40 *** | 0.11 | 0.35 *** | |
| 2022 | 0.19 ** | 0.20 ** | 0.45 *** | 0.25 *** | 0.48 *** | |
| Shagalaly | 2021 | 0.49 *** | 0.21 ** | 0.56 *** | 0.30 *** | 0.65 *** |
| 2022 | 0.44 *** | 0.20 ** | 0.44 *** | 0.32 *** | 0.35 *** | |
| Shortandy | 2020 | - | 0.29 *** | 0.38 *** | 0.33 *** | 0.38 *** |
| 2021 | 0.42 *** | 0.26 *** | 0.37 *** | 0.25 *** | 0.37 *** | |
| 2022 | 0.51 *** | 0.29 *** | 0.36 *** | 0.33 *** | 0.48 *** | |
| All | 2020 | 0.83 *** | 0.78 *** | 0.81 *** | 0.77 *** | 0.82 *** |
| 2021 | 0.84 *** | 0.75 *** | 0.77 *** | 0.63 *** | 0.79 *** | |
| 2022 | 0.77 *** | 0.76 *** | 0.79 *** | 0.64 *** | 0.79 *** | |
| Gene/Marker/Translocation | Trait | Number of Genotypes | Trait Average Value | Marker Effect 1, % | Number of Trials | |||
|---|---|---|---|---|---|---|---|---|
| −Marker | +Marker | −Marker | +Marker | Total | with Sign. Differences | |||
| Rht8 | Plant height, cm | 30 | 111 | 73.7 ± 0.9 | 74.3 ± 0.5 | −0.8 | 9 | 0 |
| TaPpdDD002 | Days to heading | 114 | 28 | 36.2 ± 0.2 | 36.7 ± 0.4 | +1.4 | 6 | 1 |
| Vrn-B1 | 98 | 44 | 36.0 ± 0.4 | 36.4 ± 0.2 | −1.1 | 6 | 0 | |
| PRR73-A1 | 53 | 87 | 36.1 ± 0.2 | 36.6 ± 0.3 | −1.4 | 6 | 0 | |
| Vrn-A1 | Days to heading | 28 | 113 | 35.9 ± 0.2 | 37.9 ± 0.4 | −5.6 1 | 6 | 6 |
| Yield, g/m2 | 225 ± 2 | 231 ± 3 | −2.7 1 | 11 | 6 | |||
| TaMOT1-D1 | Days to heading | 70 | 61 | 35.8 ± 0.2 | 36.5 ± 0.3 | −1.9 1 | 6 | 5 |
| Yield, g/m2 | 221 ± 3 | 231 ± 2 | −4.5 1 | 11 | 8 | |||
| GS5-2334-SNP | TKW, g | 103 | 37 | 32.7 ± 0.2 | 32.9 ± 0.4 | +0.6 | 11 | 0 |
| Yield, g/m2 | 223 ± 2 | 228 ± 3 | +0.2 | 11 | 0 | |||
| TaGS-D1 | TKW | 42 | 97 | 37.8 ± 0.3 | 38.6 ± 0.2 | +2.1 1 | 11 | 9 |
| Yield, g/m2 | 221 ± 4 | 228 ± 2 | +3.2 1 | 11 | 6 | |||
| ISBW10-SM2 1 | Total spikes/plant | 28 | 114 | 2.05 ± 0.04 1 | 2.21 ± 0.02 1 | +7.8 1 | 3 | 3 |
| Prod. spikes/plant | 1.63 ± 0.02 1 | 1.70 ± 0.01 1 | +4.3 1 | 5 | 3 | |||
| Yield, g/m2 | 222 ± 4 | 227 ± 2 | +2.2 | 11 | 2 | |||
| TaCwi-4A | Drought yield | 113 | 26 | 226 ± 2 | 227 ± 3 | −0.4 | 11 | 1 |
| Dreb1 | Drought yield | 64 | 78 | 224 ± 3 | 229 ± 2 | +2.2 | 11 | 1 |
| 1B.1R | Yield, g/m2 | 111 | 31 | 223 ± 2 | 237 ± 2 | +6.2 1 | 11 | 7 |
| Stem rust, % | 27.6 ± 1.3 1 | 6.6 ± 1.1 1 | −569.7 1 | 1 | 1 | |||
| ISBW3-BM | Yield, g/m2 | 97 | 44 | 220 ± 3 | 229 ± 3 | +4.1 1 | 11 | 9 |
| ISBW11-GY | Yield, g/m2 | 54 | 86 | 221 ± 3 | 231 ± 2 | +4.5 1 | 11 | 8 |
| ISBW1-GY | Yield, g/m2 | 31 | 111 | 222 ± 4 | 228 ± 2 | −2.7 | 11 | 0 |
| ISBW2-GY | Yield, g/m2 | 113 | 27 | 225 ± 2 | 233 ± 3 | +3.5 1 | 11 | 8 |
| IWAB8036 | Stem rust, % | 67 | 75 | 22.6 ± 1.6 1 | 23.4 ± 1.9 1 | −3.4 | 1 | 0 |
| Breeding Program | Number of Genotypes Evaluated | Favorable Allele Frequency (%) for Target Traits: | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Days to Heading | Yield, Stem Rust | TKW, Yield | Spike Number, Yield | Biomass, Yield | Yield | All | ||||
| TaMOT 1-D1 | vrn-A1 | 1B.1R | TaGS-D1b | ISBW10-SM2 (G) | ISBW3-BM (C) | ISBW11-GY (C) | ISBW2-GY (T) | Average | ||
| Kazakhstan | ||||||||||
| Aktobe AES | 22 | 20.0 | 0 | 4.5 | 86.4 | 77.3 | 81.8 | 54.5 | 68.2 | 49.1 |
| East Kaz. AES | 21 | 33.3 | 95.0 | 47.6 | 100.0 | 23.8 | 42.9 | 80.0 | 100.0 | 65.3 |
| Fiton Breeding Co. | 16 | 23.1 | 18.8 | 31.3 | 62.5 | 62.5 | 50.0 | 25.0 | 93.3 | 45.8 |
| Karabalyk AES | 25 | 45.8 | 40.0 | 4.2 | 84.0 | 62.5 | 56.0 | 56.0 | 84.0 | 54.1 |
| Karagandy AES | 36 | 65.7 | 41.7 | 0.0 | 88.6 | 100.0 | 97.2 | 52.8 | 86.1 | 66.5 |
| Kazakh Farm. Inst. | 28 | 33.3 | 10.7 | 3.6 | 53.6 | 67.9 | 64.3 | 46.4 | 82.1 | 45.2 |
| Kazakh Grain Cent. | 14 | 71.4 | 28.6 | 0.0 | 92.9 | 100.0 | 92.9 | 85.7 | 92.9 | 70.6 |
| Pavlodar AES | 25 | 70.8 | 45.8 | 0.0 | 72.0 | 84.0 | 80.0 | 50.0 | 92.0 | 61.8 |
| Total/average | 187 | 45.4 | 35.1 | 11.4 | 80.0 | 72.3 | 70.6 | 56.3 | 87.3 | 57.3 |
| Russia | ||||||||||
| Altay Res. Cent. | 27 | 69.2 | 34.6 | 3.7 | 86.4 | 85.2 | 51.9 | 74.1 | 96.3 | 62.7 |
| Chelyabinsk ARI | 14 | 53.8 | 0.0 | 21.4 | 41.7 | 100.0 | 35.7 | 78.6 | 69.2 | 50.1 |
| Kurgan ARI | 21 | 35.0 | 4.8 | 14.3 | 57.1 | 95.2 | 85.7 | 85.7 | 92.9 | 58.8 |
| Kurgan Seeds Co. | 15 | 71.4 | 7.1 | 57.1 | 71.4 | 93.3 | 73.3 | 86.7 | 60.0 | 65.0 |
| Siberian ARI | 15 | 47.4 | 0.0 | 21.1 | 78.9 | 68.4 | 52.6 | 42.1 | 77.8 | 48.5 |
| Omsk Agr. Univ. | 22 | 57.1 | 4.5 | 27.3 | 45.5 | 86.4 | 71.4 | 63.6 | 71.4 | 53.4 |
| Omsk Res. Cent. | 21 | 70.0 | 4.8 | 57.1 | 61.9 | 90.5 | 90.5 | 66.7 | 90.5 | 66.5 |
| Total/average | 135 | 57.7 | 8.0 | 28.9 | 63.3 | 88.4 | 65.9 | 71.1 | 79.7 | 57.9 |
| # | Genotype | Grain Yield 1, g/m2 | Days to Head. 2 | TKW 1, g | 1B.1R | TaMOT 1-D1-Ria | vrn-A1 | TaGS-D1b | ISBW 10-SM2-G | ISBW 2-GY-C | ISBW 3-BM-C | ISBW 11-GY-T |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Early heading group (35–39.9 days) | ||||||||||||
| 27 | Novosib.18 | 273 | 39.6 | 34.3 | − | − | − | + | + | − | − | − |
| 78 | Lut. 166-CΠ94 | 272 | 39.5 | 42.7 | − | + | − | + | + | + | + | + |
| 127 | Lut. KS 963 | 270 | 39.8 | 33.6 | − | − | − | + | + | − | + | + |
| 38 | Lut.1193 | 267 | 39.3 | 34.1 | − | − | − | + | + | + | − | + |
| 52 | Lut.2102 | 263 | 39.7 | 38.1 | − | + | − | + | + | − | + | − |
| Intermediate heading group (40–44 days) | ||||||||||||
| 41 | Saratov. 75 | 317 | 40.8 | 37.5 | − | − | − | + | + | − | + | − |
| 110 | Lut.37-17 | 313 | 43.7 | 40.3 | + | − | − | + | + | + | + | + |
| 36 | Ekada 113 | 311 | 42.5 | 39.5 | − | + | − | + | + | + | + | − |
| 126 | Line-241-00-4 | 306 | 43.3 | 38.1 | + | + | − | − | + | + | − | + |
| 130 | Erythrosp. 78 | 305 | 40.5 | 34.8 | − | + | − | − | + | − | + | + |
| 146 | Lut.186/04-61 | 302 | 43.5 | 39.0 | + | + | − | + | + | + | + | + |
| 49 | Lut.1764 | 300 | 43.0 | 38.4 | − | + | + | + | + | + | + | + |
| 132 | Lut.126-05 | 299 | 41.2 | 36.7 | − | + | − | − | + | − | + | + |
| 70 | Samgau | 297 | 42.2 | 40.6 | − | − | − | + | + | − | − | − |
| 28 | Lut. 307/97-23 | 297 | 43.6 | 36.8 | + | + | − | + | − | + | + | + |
| 148 | SPChS 69 | 296 | 43.0 | 39.0 | − | + | − | + | + | + | − | − |
| 50 | Lut.1082 | 296 | 43.4 | 40.0 | − | + | + | + | + | + | + | + |
| 116 | OK-1 | 293 | 41.8 | 39.9 | + | − | − | + | + | + | + | + |
| 112 | Lut.106-0/2003 | 293 | 42.2 | 36.3 | − | + | + | − | − | + | + | − |
| 68 | Lutescens 9-33 | 290 | 42.0 | 37.2 | − | + | + | − | − | + | + | − |
| 115 | A-125 | 290 | 41.0 | 35.2 | − | + | − | + | + | + | − | − |
| 18 | GVK 1857/9 | 288 | 43.7 | 39.2 | − | + | + | + | − | + | − | + |
| 107 | Chebarkul. | 288 | 42.1 | 38.7 | − | − | − | + | + | + | − | + |
| 1 | Altay. zhnitsa | 288 | 41.9 | 37.7 | − | + | − | + | + | + | − | + |
| Late heading group (44–48 days) | ||||||||||||
| 21 | Zauljbinka | 327 | 44.5 | 35.7 | + | − | + | + | − | + | − | + |
| 6 | Tobol. 1 | 302 | 44.8 | 38.4 | − | + | − | + | + | + | + | + |
| 7 | Lut.1012 | 301 | 44.5 | 37.9 | − | + | − | + | + | + | − | + |
| 69 | Pavlodar. 11 | 298 | 44.3 | 34.6 | − | − | − | − | + | + | + | − |
| 40 | LD-25 | 293 | 45.5 | 37.8 | − | + | + | + | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgounov, A.; Babkenov, A.; Ben, C.; Chudinov, V.; Dolinny, Y.; Dreisigacker, S.; Fedorenko, E.; Gentzbittel, L.; Rasheed, A.; Savin, T.; et al. Molecular Markers Help with Breeding for Agronomic Traits of Spring Wheat in Kazakhstan and Siberia. Genes 2024, 15, 86. https://doi.org/10.3390/genes15010086
Morgounov A, Babkenov A, Ben C, Chudinov V, Dolinny Y, Dreisigacker S, Fedorenko E, Gentzbittel L, Rasheed A, Savin T, et al. Molecular Markers Help with Breeding for Agronomic Traits of Spring Wheat in Kazakhstan and Siberia. Genes. 2024; 15(1):86. https://doi.org/10.3390/genes15010086
Chicago/Turabian StyleMorgounov, Alexey, Adylkhan Babkenov, Cécile Ben, Vladimir Chudinov, Yuriy Dolinny, Susanne Dreisigacker, Elena Fedorenko, Laurent Gentzbittel, Awais Rasheed, Timur Savin, and et al. 2024. "Molecular Markers Help with Breeding for Agronomic Traits of Spring Wheat in Kazakhstan and Siberia" Genes 15, no. 1: 86. https://doi.org/10.3390/genes15010086
APA StyleMorgounov, A., Babkenov, A., Ben, C., Chudinov, V., Dolinny, Y., Dreisigacker, S., Fedorenko, E., Gentzbittel, L., Rasheed, A., Savin, T., Shepelev, S., Zhapayev, R., & Shamanin, V. (2024). Molecular Markers Help with Breeding for Agronomic Traits of Spring Wheat in Kazakhstan and Siberia. Genes, 15(1), 86. https://doi.org/10.3390/genes15010086