
Association of Variants in Innate Immune Genes TLR4 and TLR5 with Reproductive and Milk Production Traits in Czech Simmental Cattle

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. DNA Samples
2.3. Sequencing with PacBio Technology
2.4. Low-Coverage Whole-Genome Sequencing
2.5. Processing of Sequencing Data
2.6. Mutation Effect Prediction
2.7. Genotyping
2.8. Phenotypic Traits
2.9. Statistical Tests
2.10. Quantitative Trait Loci
3. Results
3.1. Diversity Detected
3.2. Variant Effect Prediction
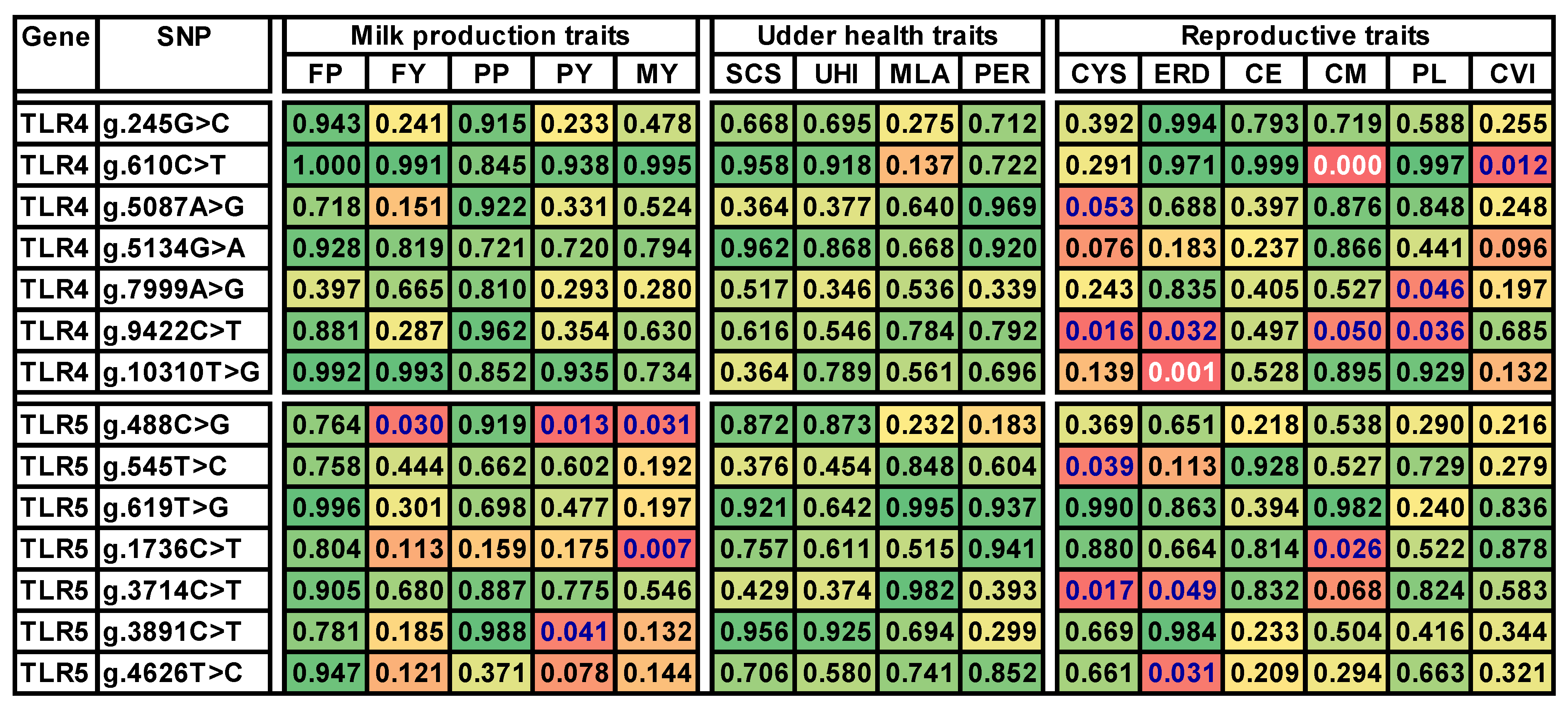
3.3. Association between Genotypes and Trait Values
3.4. Additional Statistical Tests
4. Discussion
4.1. Aberrant Hardy–Weinberg Equilibrium
4.2. Associations with Phenotypic Traits
4.3. Association Patterns Related to the TLR4 and -5 Mechanisms of Action
4.4. Context of the Known QTLs
4.5. Impact on Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Capitan, A.; Michot, P.; Baur, A.; Saintilan, R.; Hoze, C.; Valour, D.; Guillaume, F.; Boichon, D.; Barbat, A.; Boichard, D.; et al. Genetic tools to improve reproduction traits in dairy cattle. Reprod. Fer. Dev. 2014, 27, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Boichard, D.; Ducrocq, V.; Fritz, S. Sustainable dairy cattle selection in the genomic era. J. Anim. Breed. Genet. 2015, 132, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Jungi, T.W.; Farhat, K.; Burgener, I.A.; Werling, D. Toll-like receptors in domestic animals. Cell Tissue Res. 2011, 343, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Jann, O.C.; Werling, D.; Chang, J.S.; Haig, D.; Glass, E.J. Molecular evolution of bovine Toll-like receptor 2 suggests substitutions of functional relevance. BMC Evol. Biol. 2008, 8, 288. [Google Scholar] [CrossRef]
- Seabury, C.M.; Seabury, P.M.; Decker, J.E.; Schnabel, R.D.; Taylor, J.F.; Womack, J.E. Diversity and evolution of 11 innate immune genes in Bos taurus taurus and Bos taurus indicus cattle. Proc. Natl. Acad. Sci. USA 2010, 107, 151–156. [Google Scholar] [CrossRef]
- Fisher, C.A.; Bhattarai, E.K.; Osterstock, J.B.; Dowd, S.E.; Seabury, P.M.; Vikram, M.; Whitlock, R.H.; Schukken, Y.H.; Schnabel, R.D.; Taylor, J.F.; et al. Evolution of the bovine TLR gene family and member associations with Mycobacterium avium subspecies paratuberculosis infection. PLoS ONE 2011, 6, 11. [Google Scholar] [CrossRef]
- Koets, A.; Santema, W.; Mertens, H.; Oostenrijk, D.; Keestra, M.; Overdijk, M.; Labouriau, R.; Franken, P.; Frijters, A.; Nielen, M.; et al. Susceptibility to paratuberculosis infection in cattle is associated with single nucleotide polymorphisms in Toll-like receptor 2 which modulate immune responses against Mycobacterium avium subspecies paratuberculosis. Prev. Vet. Med. 2010, 93, 305–315. [Google Scholar] [CrossRef]
- Ruiz-Larrañaga, O.; Manzano, C.; Iriondo, M.; Garrido, J.M.; Molina, E.; Vazquez, P.; Juste, R.A.; Estonba, A. Genetic variation of toll-like receptor genes and infection by Mycobacterium avium ssp. paratuberculosis in Holstein-Friesian cattle. J. Dairy Sci. 2011, 94, 3635–3641. [Google Scholar] [CrossRef]
- Kannaki, T.R.; Shanmugam, M.; Verma, P.C. Toll-like receptors and their role in animal reproduction. Anim. Reprod. Sci. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Sharma, B.S.; Leyva, I.; Schenkel, F.; Karrow, N.A. Association of toll-like receptor 4 polymorphisms with somatic cell score and lactation persistency in Holstein bulls. J. Dairy Sci. 2006, 89, 3626–3635. [Google Scholar] [CrossRef] [PubMed]
- Beecher, C.; Daly, M.; Childs, S.; Berry, D.P.; Magee, D.A.; McCarthy, T.V.; Giblin, L. Polymorphisms in bovine immune genes and their associations with somatic cell count and milk production in dairy cattle. BMC Genet. 2010, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Bjelka, M.; Novák, K. Association of TLR gene variants in a Czech Red Pied cattle population with reproductive traits. Vet. Immunol. Immunopathol. 2020, 220, 109997. [Google Scholar] [CrossRef] [PubMed]
- White, S.N.; Taylor, K.H.; Abbey, C.A. Haplotype variation in bovine Toll-like receptor 4 and computational prediction of a positively selected ligand-binding domain. Proc. Natl. Acad. Sci. USA 2003, 100, 10364–10369. [Google Scholar] [CrossRef] [PubMed]
- Seabury, C.M.; Cargill, E.J.; Womack, J.E. Sequence variability and protein domain architectures for bovine Toll-like receptors 1, 5, and 10. Genomics 2007, 90, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Novák, K.; Bjelka, M.; Samake, K.; Valčíková, T. Potential of TLR-gene diversity in Czech indigenous cattle for resistance breeding as revealed by hybrid sequencing. Arch. Anim. Breed. 2019, 62, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Schatz, M.C.; Walenz, B.P.; Martin, J.; Howard, J.T.; Ganapathy, G.; Wang, Z.; Rasko, D.A.; McCombie, W.R.; Jarvis, E.D.; et al. Hybrid error correction and de novo assembly of single-molecule sequencing reads. Nat. Biotech. 2012, 30, 692. [Google Scholar] [CrossRef]
- Sim, N.L.; Kumar, P.; Hu, J.; Henikoff, S.; Schneider, G.; Ng, P.C. SIFT web server: Predicting effects of amino acid substitutions on proteins. Nucl. Acids Res. 2012, 40, W452–W457. [Google Scholar] [CrossRef]
- Richardson, I.W.; Berry, D.P.; Wiencko, H.L.; Higgins, I.M.; More, S.J.; McClure, J.; Lynn, D.J.; Bradley, D.G. A genome-wide association study for genetic susceptibility to Mycobacterium bovis infection in dairy cattle identifies a susceptibility QTL on chromosome 23. Genet. Sel. Evol. 2016, 48, 19. [Google Scholar] [CrossRef]
- Samaké, K.; Novák, K. Haplotype disequilibrium in the TLR genes of Czech Red Pied cattle. Diversity 2023, 15, 811. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Meyer, K. WOMBAT—A tool for mixed model analyses in quantitative genetics by restricted maximum likelihood (REML). J. Zhejiang Univ. Sci. B 2007, 8, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.S.; Mount, J.; Karrow, N.A. Functional characterization of a single nucleotide polymorphism in the 5′-UTR region of the bovine toll-like receptor 4 gene. Dev. Biol. 2008, 132, 331–336. [Google Scholar]
- Wang, M.Q.; Song, H.L.; Zhu, X.R.; Xing, S.Y.; Zhang, M.R.; Zhang, H.M.; Wang, X.L.; Yang, Z.P.; Ding, X.D.; Karrow, N.A.; et al. Toll-like receptor 4 gene polymorphisms influence milk production traits in Chinese Holstein cows. J. Dairy Res. 2018, 85, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.X.; Zhao, H.; Zhou, H.H. False discovery rate control with groups. J. Am. Stat. Assoc. 2010, 105, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Balding, D.J. A tutorial on statistical methods for population association studies. Nature Rev. Genet. 2006, 7, 781–791. [Google Scholar] [CrossRef]
- Opsal, M.A.; Lien, S.; Brenna-Hansen, S.; Olsen, H.G.; Våge, D.I. Association analysis of the constructed linkage maps covering TLR2 and TLR4 with clinical mastitis in Norwegian Red cattle. J. Anim. Breed. Genet. 2008, 125, 110–118. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.; Bortfeldt, R.H.; Regens, J.; Brockmann, G.A. Single nucleotide polymorphism and haplotype effects associated with somatic cell score in German Holstein cattle. Genet. Sel. Evol. 2014, 46, 35. [Google Scholar] [CrossRef]
- Novák, K.; Pikousová, J.; Czerneková, V.; Mátlová, V. Diversity of the TLR4 immunity receptor in Czech native cattle breeds revealed using the Pacific Biosciences sequencing platform. Anim. Biotech. 2017, 28, 228–236. [Google Scholar] [CrossRef]
- Mullen, M.P.; McClure, M.C.; Kearney, J.F. Relationships between a TLR4 allele associated with IBK and production traits in dairy cattle. In Book of Abstracts of the 69th Annual Meeting of the European Federation of Animal Science, Dubrovnik, Croatia, 27–31 August 2018; Wageningen Academic Publishers: Wageningen, The Netherlands, 2018; p. 611. [Google Scholar]
- El-Domany, W.B.; Radwan, H.A.; Ateya, A.I.; Ramadan, H.H.; Marghani, B.H.; Nasr, S.M. Genetic polymorphisms in LTF/EcoRI and TLR4/AluI loci as candidates for milk and reproductive performance assessment in Holstein cattle. Repr. Dom. Anim. 2019, 54, 678–686. [Google Scholar] [CrossRef]
- Sheldon, I.M. Genes and environmental factors that influence disease resistance to microbes in the female reproductive tract of dairy cattle. Reprod. Fert. Dev. 2015, 27, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Bicalho, M.L.S.; Santin, T.; Rodrigues, M.X.; Marques, C.E.; Lima, S.F.; Bicalho, R.C. Dynamics of the microbiota found in the vaginas of dairy cows during the transition period: Associations with uterine diseases and reproductive outcome. J. Dairy Sci. 2017, 100, 3043–3058. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.; Barker, G.; Lappas, M. TLR2, TLR3 and TLR5 regulation of pro-inflammatory and pro-labour mediators in human primary myometrial cells. J. Reprod. Immunol. 2017, 122, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wahid, H.H.; Dorian, C.L.; Chin, P.Y.; Hutchinson, M.R.; Rice, K.C.; Olson, D.M.; Moldenhauer, L.M.; Robertson, S.A. Toll-like receptor 4 is an essential upstream regulator of on-time parturition and perinatal viability in mice. Endocrinology 2015, 156, 3828–3841. [Google Scholar] [CrossRef] [PubMed]
- Elovitz, M.A.; Wang, Z.; Chien, E.K.; Rychlik, D.F.; Phillippe, M. A new model for inflammation-induced preterm birth: The role of platelet-activating factor and Toll-like receptor-4. Am. J. Pathol. 2003, 163, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Kadam, L.; Gomez-Lopez, N.; Mial, T.N.; Kohan-Ghadr, H.R.; Drewlo, S. Rosiglitazone regulates TLR4 and rescues HO-1 and NRF2 expression in myometrial and decidual macrophages in inflammation-induced preterm birth. Reprod. Sci. 2017, 24, 1590–1599. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Glynn, D.J.; Sharkey, D.J.; Robertson, S.A. Female tract response to seminal fluid in mice. Biol. Reprod. 2015, 93, 3. [Google Scholar] [CrossRef]
- Cole, J.B.; Wiggans, G.R.; Ma, L.; Sonstegard, T.S.; Lawlor, T.J., Jr.; Crooker, B.A.; Van Tassell, C.P.; Yang, J.; Wang, S.; Matukumalli, L.K.; et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary U.S. Holstein cows. BMC Genom. 2011, 12, 408. [Google Scholar] [CrossRef]
- Purfield, D.C.; Bradley, D.G.; Evans, R.D.; Kearney, F.J.; Berry, D.P. Genome-wide association study for calving performance using high-density genotypes in dairy and beef cattle. Genet. Sel. Evol. 2015, 47, 47. [Google Scholar] [CrossRef]

{kind=link}
| SNP Identifier | Chromosome/ /Position | Substitution a | PCR Fragment | Extension Primer | Multiplex Part | SNP Type + Effect Prediction | |
|---|---|---|---|---|---|---|---|
| Primer Name | Sequence 5′→3′ | ||||||
| TLR4 | |||||||
| rs29017188 | 8_108829143 | g.245G>C | 4_1 | T245F | CTTCTTCTTCCTCTAACTTCCCCTC | A1 | 5′-UTR, expression |
| rs43578094 | 8_108829508 | g.610C>T | 4_1 | T610R | (T5)GGGCCCAGCACAGGGAAACTGAGCA | A2 | intron, modifier |
| rs8193046 | 8_108833985 | g.5087A>G | 4_2 | T087F | (T10)GCTAAGGTGCATGCAGGAAGACACC | A3 | intron, modifier |
| rs8193047 | 8_108834032 | g.5134G>A | 4_2 | T134R | (T13)GATTTTGTAGAGATTCAGCTCCATGCA | A4 | synonymous, low |
| rs43578100 | 8_108836897 | g.7999A>G | 4_3 | T999F | (T21)GGTTTCCTATTCAGCAGAAATATT | A5 | intron, modifier |
| rs8193060 | 8_108838320 | g.9422C>T | 4_4 | T422F | ACTCGCTCCGGATCCTAGACTGCAG | B1 | synonymous, low |
| rs8193072 | 8_108839208 | g.10310T>G | 4_5 | T010F | (T25)CCACCTGAGGAGGAGAATCCCCTGA | C4 | 3′-UTR, low |
| TLR5 | |||||||
| 16_27307783 | g.488C>G | 5_1 | T488F | (T6)CCAGGGATGAAACCCRTGTCTCCTG | B2 | upstream, modifier | |
| ss73689429 | 16_27307726 | g.545T>C | 5_1 | T545F | (T10)CCAGGGAAGTCTTGCTGGCCTACTG | B3 | upstream, modifier |
| ss73689429 | 16_27307652 | g.619T>G | 5_2 | T619R | CCACAGCACCTTTGAGGCTGTGAC | C1 | upstream, modifier |
| ss73689443 | 16_27306535 | g.1736C>T | 5_3 | T736F | (T2)GTACTTACAAYCATGCTTGCTATTTTT | C2 | upstream, modifier |
| rs55617187 | 16_27304557 | g.3714C>T | 5_4 | T714R | (T15)GATTGAGCCAATGGATAAAAGCACT | B4 | synonymous, low |
| rs55617251 | 16_27304380 | g.3891C>T | 5_4 | T891R | (T18)CACGAGGAACAGAGTCAAGGTGACAGT | B5 | synonymous, low |
| rs55617288 | 16_27303645 | g.4626T>C | 5_5 | T626R | (T9)GGGGTCGCAAAGAGTAGGACATGACC | C3 | downstream, modifier |
| Gene | SNP a | Number of Bulls | Genotype Frequencies Observed | Allelic Frequencies | χ2 | WT Allele Frequency in NGS | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Homozygotes 1 | Heterozygotes | Homozygotes 2 | p | q | WT Read Frequency c | % of p | ||||
| TLR4 | g.245G>C | 81 | 0.086 | 0.630 | 0.284 | 0.401 | 0.599 | 7.804 b | 0.457 | 113.9 |
| TLR4 | g.610C>T | 72 | 0.861 | 0.139 | 0.000 | 0.931 | 0.069 | 0.401 | 0.937 | 100.7 |
| TLR4 | g.5087A>G | 73 | 0.329 | 0.493 | 0.178 | 0.575 | 0.425 | 0.006 | 0.746 | 129.7 |
| TLR4 | g.5134G>A | 76 | 0.724 | 0.250 | 0.026 | 0.849 | 0.151 | 0.054 | 0.694 | 81.8 |
| TLR4 | g.7999A>G | 80 | 0.163 | 0.600 | 0.238 | 0.463 | 0.538 | 3.421 | 0.754 | 163.0 |
| TLR4 | g.9422C>T | 66 | 0.182 | 0.348 | 0.470 | 0.356 | 0.644 | 3.803 | 0.231 | 64.9 |
| TLR4 | g.10310T>G | 84 | 0.679 | 0.321 | 0.000 | 0.839 | 0.161 | 3.080 | 0.625 | 74.5 |
| TLR5 | g.488C>G | 81 | 0.185 | 0.395 | 0.420 | 0.383 | 0.617 | 2.175 | 0.385 | 100.6 |
| TLR5 | g.545T>C | 70 | 0.414 | 0.357 | 0.229 | 0.593 | 0.407 | 4.739 b | 0.500 | 84.3 |
| TLR5 | g.619T>G | 85 | 0.706 | 0.294 | 0.000 | 0.853 | 0.147 | 2.527 | 0.180 | 21.1 |
| TLR5 | g.1736C>T | 60 | 0.417 | 0.400 | 0.183 | 0.617 | 0.383 | 1.422 | 0.940 | 152.4 |
| TLR5 | g.3714C>T | 77 | 0.273 | 0.494 | 0.234 | 0.519 | 0.481 | 0.010 | 0.500 | 96.3 |
| TLR5 | g.3891C>T | 86 | 0.814 | 0.186 | 0.000 | 0.907 | 0.093 | 0.905 | 0.950 | 104.7 |
| TLR5 | g.4626T>C | 63 | 0.095 | 0.429 | 0.476 | 0.310 | 0.690 | 0.000 | 0.700 | 226.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novák, K.; Valčíková, T.; Samaké, K.; Bjelka, M. Association of Variants in Innate Immune Genes TLR4 and TLR5 with Reproductive and Milk Production Traits in Czech Simmental Cattle. Genes 2024, 15, 24. https://doi.org/10.3390/genes15010024
Novák K, Valčíková T, Samaké K, Bjelka M. Association of Variants in Innate Immune Genes TLR4 and TLR5 with Reproductive and Milk Production Traits in Czech Simmental Cattle. Genes. 2024; 15(1):24. https://doi.org/10.3390/genes15010024
Chicago/Turabian StyleNovák, Karel, Terezie Valčíková, Kalifa Samaké, and Marek Bjelka. 2024. "Association of Variants in Innate Immune Genes TLR4 and TLR5 with Reproductive and Milk Production Traits in Czech Simmental Cattle" Genes 15, no. 1: 24. https://doi.org/10.3390/genes15010024
APA StyleNovák, K., Valčíková, T., Samaké, K., & Bjelka, M. (2024). Association of Variants in Innate Immune Genes TLR4 and TLR5 with Reproductive and Milk Production Traits in Czech Simmental Cattle. Genes, 15(1), 24. https://doi.org/10.3390/genes15010024