Abstract
Reed canary grass (Phalaris arundinacea L.) is known for its tolerance to drought, heavy metals, and waterlogging, making it a popular choice for forage production and wetland restoration in the Qinghai-Tibet Plateau (QTP). To accurately assess gene expression in reed canary grass under different abiotic stresses, suitable reference genes need to be identified and validated. Thirteen candidate reference gene sequences were selected and screened using RT-qPCR to detect their expression levels in reed canary grass leaves under drought, salt, cadmium, and waterlogging stresses. Four algorithms were used to assess the stability of the expression levels of the candidate reference genes. The most stably expressed genes were UBC and H3 under drought Cd, ETF and CYT under salt stress, and ETF and TUB under waterlogging stress. GAPDH was found to be less stable under abiotic stresses. PIP-1, PAL, NAC 90, and WRKY 72A were selected as response genes for quantitative expression assessment under drought, salt, Cd, and waterlogging stresses to confirm the accuracy of the selected stable reference genes. These results provide a theoretical reference for assessing gene expression in reed canary grass under abiotic stresses.
1. Introduction
Reed canary grass (RCG, P. arundinacea L.) is a cool-season perennial grass valued for its use in forage, bioenergy, and wetland restoration [1]. There is a wide distribution across the temperate areas of North America, Europe, and Asia [2]. Due to its rapid growth, high yield, and tall height, Reed canary grass is an exceptional forage species, serving as a vital source of hay and silage on the Qinghai-Tibet Plateau (QTP) [3]. Additionally, reed canary grass is often used to filter pollutants from aquatic environments due to its tolerance to waterlogging. It can absorb pollutants such as Al3+ and is a popular choice for wetland restoration [4,5]. As an energy plant, reed canary grass provides clean energy and is drought-tolerant compared to most annual plants. It requires less tillage and maintenance, and its deep-rooted nature improves nutrient use efficiency [6]. However, drought, soil salinity, heavy metals, and waterlogging are among the abiotic stresses that can limit its growth and development. Studying the transcriptional response of plants during abiotic stress can help identify genes important for critical plant pathways, leading to the development of better and more resilient canary grass varieties [7,8,9]. The study of reed canary grass is crucial for understanding its resistance to abiotic stress and gene exploitation. However, limited genomic sequence information and the lack of stable reference genes in reed canary grass hinder gene mining, gene function validation, and gene expression analysis. This lack of precision in key gene expression analysis limits the study of abiotic stress in reed canary grass. Therefore, it is crucial to further research and development in this field to identify stable reference genes in reed canary grass.
For quantifying transcript expression, real-time quantitative PCR (RT-qPCR) is widely used because it is highly sensitive, specific, reproducible, and high throughput [10,11]. The accuracy of RT-qPCR depends on expression levels of reference genes, and its results are affected by RNA quality, stable reference gene expression, cDNA synthesis, and other factors; therefore, to calibrate for experimental variability, stable reference genes must be selected for amplification with the target gene [12]. Stable reference genes have been reported to include cytoskeletal proteins and essential components in basic cellular biochemical pathways, such as ubiquitin-binding enzymes, 50S ribosomal protein L2, tubulin, etc. [13]. Although reference genes are generally considered to be stable under all conditions, it is important to note that their expression levels may not always remain consistent across different conditions, growth stages, and physiological states. Therefore, to improve the accuracy of RT-qPCR results, stable reference genes must be tested under different conditions and physiological states [14]. GeNorm, NormFinder, BestKeeper, and RefFinder are four common algorithms to evaluate the stability of candidate reference genes [15,16]. Reference gene screens have widely used these algorithms, such as those for maize (Zea mays L.) and larch (Larix gmelinii (Rupr.) Kuzen.) [17,18]. However, there are still many species lacking reference genes, and few studies have detailed the screening of reed canary grass reference genes under abiotic stress, which significantly affects the accuracy of reed canary grass gene expression.
This study aimed to identify and validate reference genes for stable expression under abiotic stresses in reed canary grass using transcriptome sequencing data. The stability of 13 potential reference genes was examined under different abiotic stresses, including salt, drought, cadmium (Cd), and waterlogging stresses. Additionally, the expression levels of PIP-1, PAL, NAC 90, and WRKY 72A genes were determined to validate the best candidate reference genes. Overall, selected and identified reference genes are important for understanding the molecular mechanisms underlying abiotic stress responses in reed canary grass and related species.
2. Materials and Methods
2.1. Plant Materials and Abiotic Stress Treatments
The seeds of reed canary grass (cv. Chuanxi; seeds obtained from Sichuan Agricultural University, Chengdu, China) were soaked in 10% sodium hydroxide for 5 min to improve germination, then rinsed in sterile distilled water. Finally, each pot was evenly sown in plastic pots with quartz sand. Twelve pots were planted with a pot size of 20 cm × 15 cm × 5 cm, 1.3 g of seeds per pot, followed by 1/2 times the amount of Hoagland nutrient solution. After sowing, the pots were grown in an incubator for 90 days under the following conditions: (1) day and night temperatures of 23 °C and 19 °C respectively; (2) photoperiod of 12 h; (3) relative humidity of 75%; (4) light intensity of 250 μmol (m−2·s−1). When reed canary grass reached the three-leaf stage, the following treatments were applied: 20%PEG-6000 solution with drought stress, 300 mmol·L−1 NaCl with salt stress, 50 μmol·L−1 CdCl with Cd stress, and waterlogging of the whole plant with waterlogging stress, with three replicates used for each treatment group. Then, leaves were collected at 0 h, 0.5 h, 1.5 h, 3 h, 6 h, 12 h, 24 h, 48 h and 72 h of each treatment, respectively, and the collected samples were immediately frozen in liquid nitrogen and stored in a refrigerator at −80 °C for subsequent RNA extraction.
2.2. RNA Extraction and Reverse Transcription
A frozen leaf of 0.1 g of reed canary grass was selected and pulverized by a high-throughput tissue grinder, and total RNA was extracted using the M5 HiPer Plant Complex Mini Kit (Beijing, China). Analyzing the RNA concentration and quality with the NanoDrop1 ND-1000 Spectrophotometer (Nano Drop Technologies, Wilmington, DE, USA) and agarose gel electrophoresis, respectively, confirmed the quality of the extracted RNA. The quality of the extracted RNA was confirmed by analyzing the concentration and quality of the RNA using a NanoDrop1 ND-1000 spectrophotometer (Nano Drop Technologies, Wilmington, DE, USA) and agarose gel electrophoresis, respectively. For further analysis, the first strand of cDNA was reverse transcribed using the ABSCRIPT III RT Master Mix for qPCR with gDNA Remover (Wuhan, China).
2.3. Primer Design and Validation
Based on the RNA-seq of the reed canary grass, thirteen candidate reference genes were selected which included UBC (Ubiquitin-conjugating enzyme E2 34); GATP (Glyceraldehyde-3-phosphate dehydrogenase); ETF (Eukaryotic translation initiation factor 4E); TUB (Tubulin alpha-3 chain); CYT (Cytochrome P450); ADPF (ADP-ribosylation factor 1); H3 (Histone H3); 50Sr (50S ribosomal protein L2); MD (3-isopropylmalate dehydrogenase 2); Heats (Heat shock 70 kDa protein); Cul1 (Cullin-1); TATA (TATA-binding protein); ACT (Actin-7) [19,20]. The primers were then designed and screened for the selected candidate reference gene sequence (Table S1). The sequences of the selected candidate reference genes were used for the design and screening of the primers. Primer design was performed by Primer5 software, and primer synthesis was performed by You Kang Biotechnology (Chengdu, China) (Table S2). A melting curve analysis of the RT-qPCR reactions confirmed the specificity of the primers for candidate reference genes.
2.4. Quantitative RT-qPCR
For quantitative analysis, RT-qPCR was performed using the CFX96 RT-qPCR system (Bio-Rad, Singapore) and Power SYBR green PCR master mix (Wuhan, China). The temperature was maintained at zero degrees Celsius for better resolution, and the sample injection volume was kept at 10 µL. zero to one degree Celsius. The total reaction volumes were 10 µL containing 1 µL cDNA 0.2 µL forward primer. 0.2 µL reverse primer, 5 µL Genious 2 × SYBR Green Fast QPCR No ROX Mix (ABclonal, Wuhan, China) and 3.6 µL nuclease-free water. All reaction products were analyzed after 35 amplification cycles with the following steps: 10 min pre-denaturation at 95 °C, denaturation 15 s at 95 °C and annealing 1 min at 60 °C. In each abiotic stress sample, three biological replicates were performed, as well as three technical replicates.
2.5. Data Analysis
GeNorm [21], NormFinder [22], BestKeeper [23], and RefFinder (http://www.leonxie.com/referencegene.php, accessed on 17 January 2023) were used to determine the cycle threshold (Ct) for each reference gene. A Ct value must be converted into a relative quantification Q value as described above before NormFinder can analyze gene stability. This is mainly through the equation Q = 2−∆Ct, where ∆Ct = Ct sample − Ct min, t sample indicates the Ct value of the housekeeping gene, and Ct min indicates the lowest Ct value of the housekeeping gene under each abiotic stress. A coefficient of variation (CV) and standard deviation (SD) were calculated by BestKeeper using Ct values, and the expression stability measurement (M) values were calculated for each candidate reference gene via the GeNorm program (SD). Finally, RefFinder was used to combine the three software calculations mentioned above and to calculate a combined ranking index of each reference gene’s geometric mean and stability. Afterwards, the results of these four algorithms are correlated using the R language. Generally, low index values were considered reference genes of high stability. To determine the optimal number of reference genes, Vn/Vn + 1 is applied. In general, a value of Vn/Vn + 1 greater than 0.15 is required for the first V(n + 1) reference gene. Otherwise, no new reference genes need to be introduced.
2.6. Validation of Reference Genes
To validate the selected reference genes, reference genes, including the two most stable reference genes and the one least stable reference gene under each abiotic stress, were used to analyze the expression levels of target genes under four abiotic stresses. The target genes for each stress were selected as follows: the PIP-1 (Aquaporin PIP1-1) was selected for drought stress, the PAL (Phenylalanine ammonia-lyase) for salt stress, the WRKY 72A (WRKY transcription factor 72A) for waterlogging stress and the NAC 90 (NAC domain-containing protein 90) for cadmium stress(Tables S3 and S4) Then following by calculation using the 2−∆∆Ct method [24].
3. Results
3.1. The Effectiveness of the Primers of 13 Specific Reference Genes
The cDNA, reverse transcribed from the total RNA of reed canary grass leaves after exposure to drought, salt, Cd, and waterlogging stresses, was used as the reaction sample for the RT-qPCR reaction. The results showed a clear single peak in the solution curve for all reference genes, indicating that the primers selected had excellent specificity and amplified the target product of each gene generated (Figure 1). Additionally, the RT-qPCR results were reproducible, and no primer dimers were produced. These findings demonstrate that the RT-qPCR results are authentic and reliable, making them suitable for use in subsequent studies.
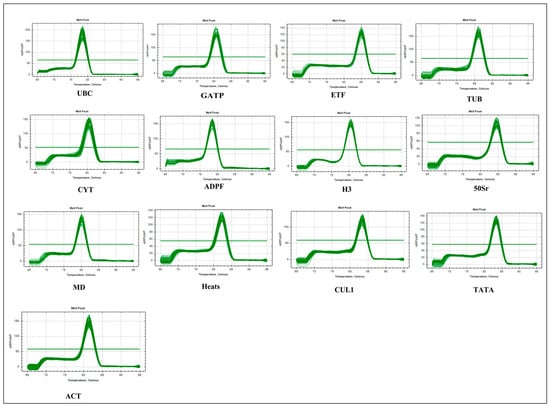
Figure 1.
Reed canary grass melt curves for 13 candidate reference genes.
3.2. Expression of Reference Gene
Ct values for reference genes are inversely related to target gene expression levels. Therefore, higher Ct values indicate a lower gene expression level in the sample. Ct values for different candidate reference genes were represented by box plots. The upper and lower box plot limits indicate the maximum and minimum Ct values, and the dispersion indicates gene stability. It is more likely that a sample with a lower dispersion has more stable gene expression. A variety of abiotic stresses led to Ct values ranging from 22.92 to 32.95 for the candidate reference genes. The UBC gene showed the highest expression abundance under drought stress, the CYT gene under salt stress, the ETF gene under waterlogging stress and the H3 gene under cadmium stress (Figure 2).
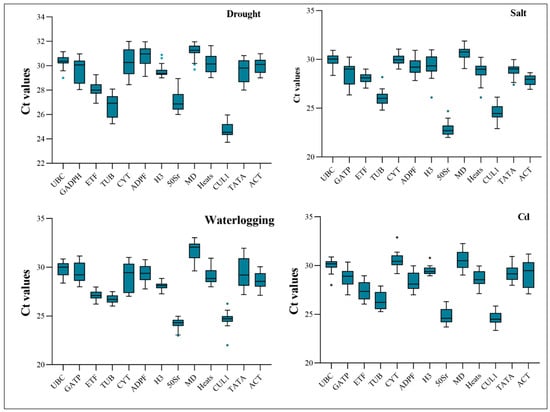
Figure 2.
Ct values of 13 candidate reference genes in reed canary grass leaves under abiotic stresses. The variation is displayed as medians values (lines across the box plot) and the circles indicates free value.
3.3. Candidate Reference Gene Stability Analysis
3.3.1. GeNorm Analysis
GeNorm software was used to calculate the M value, which represents the stability of the expression of the candidate reference gene. Generally, the lower the M value, the more stable the candidate gene is. The M value was used to rank the stability of the candidate reference genes. This research found that UBC and H3 were the most stably expressed candidate reference genes in drought and Cd stresses, with M values of 0.03 and 0.024, respectively (Figure 3). ETF and TUB ranked highest in stability in salt stress, with M values of 0.025. In waterlogging stress, TUB and H3 genes were the most stably expressed genes, with M values of 0.026. However, CYT was the least stable gene, with an M value of 0.057 (Figure 3).
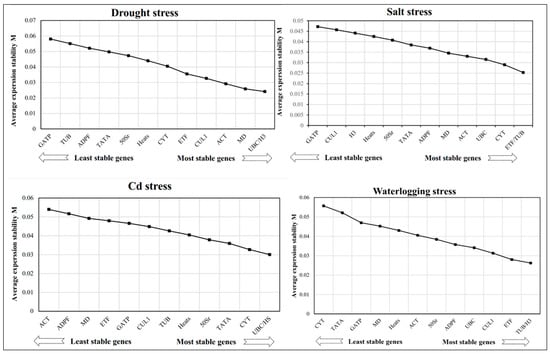
Figure 3.
Abiotic stress effects on the expression of 13 candidate reference genes in reed canary grass leaves.
A pairwise variation analysis can be useful for determining the optimal number of reference genes for accurate normalization. The pairwise variation values of standard factors are typically used, with a default threshold of 0.15 for V. When the Vn/Vn + 1 value is less than 0.15, n is considered the optimal number of reference genes, while n+1 reference genes are required when the value is greater. As Vn/Vn + 1 values fell below 0.15 under various abiotic stress, it was concluded that only two reference genes were required to normalize gene expression data (Figure 4).
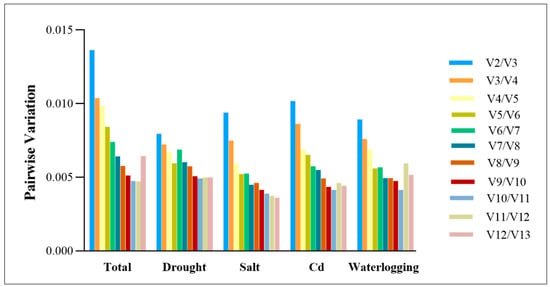
Figure 4.
Pairwise variation (V) measures were calculated for the candidate reference genes using GeNorm.
3.3.2. BestKeeper Analysis
The standard deviation (SD) and coefficient of variance (CV) of gene expression are calculated to evaluate the stability of reference gene expression. The reference gene is considered when SD is less than 1, with a higher SD indicating lower reference gene stability. In this study, H3 and UBC were found to have high stability of expression under drought stress. CYT and ETF were the most consistently expressed genes under salt stress, while H3 and UBC showed stable gene expression trends compared to other genes under Cd stress. UBC and GATP showed stable expression under waterlogging stress. However, it is important to note that these results are partially inconsistent with the results of the GeNorm algorithm and, therefore, require further analysis using other algorithms (Table 1).

Table 1.
BestKeeper expression stability values for reed canary grass reference genes.
3.3.3. NormFinder Analysis
In NormFinder V20, a variance method is used to calculate the stability of candidate reference genes (S-values), with lower S-values indicating higher gene stability. The S-values of ACT and UBC under drought stress were 0.353 and 0.372, respectively, lower than those of other candidate reference genes, indicating that ACT and UBC had higher stability under drought treatment (Table 2). ETF and CYT were more stable under salt stress, with S-values of 0.362 and 0.382, respectively. H3 and UBC were ranked as the top stability candidates under Cd stress, while H3 and ETF were more stable than other genes under waterlogging stress (Table 2).
Table 2.
Expression stability values for candidate reference genes were calculated by NormFinder.
3.3.4. RefFinder Analysis
ReferenceFinder V1.0 combines three algorithms, GeNorm, NormFinder, and BestKeeper, to rank candidate reference genes with respect to their stability. It is commonly used to analyze the combined stability of these genes. Under drought stress, UBC and H3 were the most stable genes, whereas ETF and CYT were the most stable genes under salt stress. Similarly, the best reference gene combinations were UBC and H3 under Cd stress, and ETF and TUB showed higher expression stability under waterlogging stress (Table 3).
Table 3.
The most stable and least stable reference gene combinations analyzed by RefFinder.
To compare the correlation between the four algorithms, we calculated the correlation coefficients of the results of the four algorithms. We found that the correlation between the four algorithms was greater than 0.75, of which the GeNorm and ReFfinder algorithms had the highest correlation, and the results of the calculated gene stability were similar (Figure S1).
3.4. Validation the Stability of Reference Genes
The stability of the candidate reference gene should be validated by selecting and normalizing the expression of the suitable target gene. In this study, target genes were chosen for each stress, and the two most and one least stable reference genes were selected for expression pattern analysis using the 2−∆∆Ct method (Table 3). The most stable genes (UBC and H3) were suitable for normalizing PIP-1 expression under drought stress, while the least stable gene (GATP) showed a different pattern (Figure 5A). Under salt stress, the selected most stable genes (ETF + CYP) normalized the expression of PAL genes compared to the least stable gene (GATP), and the normalization results showed different trends (Figure 5B). Similarly, under Cd stress, normalizing NAC 90 using the unstable reference gene ACT resulted in a different trend than the most stable genes (H3 and UBC) (Figure 5C). Under waterlogging stress, the most stable genes (ETF and TUB) exhibited similar trends, but the least stable reference gene (TUB) resulted in a different trend (Figure 5D). These results showed that the selected stable reference genes were reliable for this study, as they showed consistent expression patterns when used to normalize the expression of target genes under different abiotic stresses.
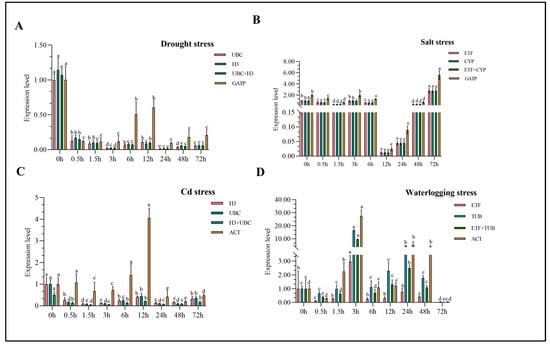
Figure 5.
Expression levels of target genes in reed canary grass leaves under drought, salt, waterlogging and heavy metal stress. Different letters indicate significant differences in the expression of the same gene among different treatments (P < 0.05). (A) represents the expression of PIP-1 in reed canary grass at different time points under drought conditions; (B) indicates the expression of PAL at different time points of salt stress in reed canary grass. (C,D) represent the expression of NAC 90 and WRKY 72A in reed canary grass at different time points under waterlogging and heavy metal stress, respectively.
4. Discussion
Despite its important forage, bioenergy and ecological restoration functions, reed canary grass faces various complex stresses, such as drought, salt, Cd and waterlogging, which hinder its production applications. Therefore, it is urgent to study the response mechanism to the abiotic stress of reed canary grass. In particular, the accuracy of gene expression analysis is the basis and key linkage of abiotic stress research and analysis [25]. Although several studies have been conducted on the abiotic stress mechanisms in reed canary grass, there has been a lack of systematic comparative analysis regarding the stability of reference gene expression under abiotic stress conditions, and it could result in potential biases of the quantification of target gene expression in previous studies. Therefore, it is crucial to investigate the stability of reference gene expression under abiotic stress in reed canary grass to ensure accurate and reliable quantification of target gene expression in future studies [26,27,28].
Reference genes are typically expressed in metabolically active cells and perform essential cellular functions. They are important in cell cycle regulation [29,30,31]. The choice of reference genes and primer design have been reported to have a significant impact on the reliability of RT-qPCR target gene results [32,33]. In this study, the 13 candidate reference genes showed differential expression, consistent with previous studies reporting that reference genes are differentially expressed under different abiotic stresses of the same species. However, it is noteworthy that both UBC and H3 performed well under drought and cadmium stress, which may indicate that UBC and H3 genes can maintain stable expression regardless of abiotic stresses. The ubiquitin-coupled enzyme (E2), a driver of ubiquitin signaling in plants, has been reported to be highly tolerant to osmotic stress and cadmium stress by overexpression of ubiquitin-coupled enzyme (E2) [34,35,36]. In addition, this study found that the expression of H3 varied with different treatments, and it suggested that H3 may be a suitable reference gene under various abiotic stresses. However, GATP, a commonly used reference gene, was less stable than the others, verified in okra (Abelmoschus esculentus) [37]. In contrast, TUB was a more stable gene for expression under waterlogging stress, which was also demonstrated in Betula alba [38], and then TUB was the least stable in Chrysoperla nipponensis [39], okra and Schima superba, which also suggests that specific reference genes exhibit different expression stability depending on the species and stress they are exposed to.
Four main algorithms are commonly used to assess the stability of reference genes, including GeNorm, NormFinder, BestKeeper and RefFinder. GeNorm has different roles and purposes. GeNorm ranking is based on the similarity of expression levels of each reference gene from different experimental samples [22]. The stability of the candidate reference gene is calculated directly by BestKeeper using a pairwise correlation analysis of the Ct values [23]. To combine the results of the above three algorithms, an overall ranking was performed using the online ranking software RefFinder, which calculates the average gene weights and obtains the final ranking results [40]. RefFinder has now been applied to Brassica juncea [41], okra [38] and cucumber (Cucumis sativus) [42] for reference gene selection. GeNorm, NormFinder, BestKeeper and RefFinder were used in combination for gene expression analysis of candidate reference genes, but the differences in the first three algorithms resulted in inconsistent ranking of the stability of the reference genes analyzed by them. In this study, H3 and UBC were the most stable reference genes when calculated by BestKeeper, and UBC and H3 were also ranked first in terms of gene expression stability when calculated using GeNorm, but UBC was ranked second when calculated using the NormFinder algorithm, and the top-ranked reference gene was ACT.
PIP-1, NAC 90, PAL and WRKY 72A were targeted genes to validate the most and least stable reference genes selected under drought, salt, Cd, and waterlogging stresses. PIP-1 is a water channel protein involved in drought stress in plants [43]. It has been reported that all 12 genes encoding water channel proteins (PIP-1, PIP-2 and NIP-2) were significantly up-regulated under drought stress in Miscanthus [44]. In this study, PIP-1 was selected as the target gene to validate a stable reference gene for drought stress, and the validation results showed that PIP-1 exhibited a general expression pattern under drought stress, demonstrating the significance of the selected stable reference gene. The PAL is the first and key enzyme in the phenylpropane pathway and is transcriptionally regulated by various environmental factors. The PAL has been reported to be effectively applied at the molecular breeding level to improve the resistance of Cyclocarya paliurus under salt stress [45]. PAL was selected as a target gene to validate the reference gene for the stabilization of reed canary grass under salt stress, and the validation results were consistent with PAL expression levels under salt stress. NAC 90 and WRKY 72A played different roles in different abiotic stresses and were selected as target genes to verify the stability of Cd stress and waterlogging stress, respectively [46]. This study selected NAC 90 and WRKY 72A as target genes to verify the stable and stable reference genes for Cd stress and waterlogging stress, respectively.
5. Conclusions
This study provides a useful guideline for screening suitable reference genes to study the expression of reed canary grass genes under abiotic stress. The results showed that UBC and H3 were more stable under drought and Cd stress; ETF and CYT were more desirable under salt stress; also, ETF and TUB were stable reference genes under waterlogging stress. These results provide an excellent functional genetic platform for studying reed molecular mechanisms under abiotic stress.
Supplementary Materials
The supporting information can be downloaded at https://www.mdpi.com/article/10.3390/genes14091790/s1, Table S1. Sequence of 13 candidate reference gene. Table S2. 13 Candidate Reference Gene Primer. Table S3. Sequences of target genes under four abiotic stresses. Table S4. Primers for target genes under four abiotic stresses. Figure S1. Pearson’s correlation heat map based on four algorithms.
Author Contributions
Conceptualization: X.M., D.L., S.B. and J.Z.; methodology: X.J., Y.X. (Yi Xiong) and Y.X. (Yanli Xiong); formal analysis: X.J., M.Y. and Q.Y.; writing—original draft preparation: X.J. and Y.X. (Yi Xiong); writing—review and editing: X.J., Y.X. (Yi Xiong), X.M. and J.Z.; visualization: X.L. and X.J.; supervision, M.Y. and X.M.; project administration: X.M. and J.Z.; funding acquisition: X.M. and J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Supported by the [Sichuan Science and Technology Program] grant number [2022YFN0035], [Sichuan Forestry and Grassland Science and Technology Innovation Team Special Funding of China] grant number [LCTD2023CZ01], [Sichuan Beef Innovation Team Project] grant number [sccxtd-2019-13], [Sichuan Forage Innovation Team Project] grant number [sccxtd-2020-16], [Sichuan Province “14th Five-Year Plan” Forage Breeding Research Project of China] grant number [2021YFYZ0013-2], and [National Forage Industry Technology System Aba Comprehensive Experimental Station of China] grant number [CARS-34].
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oram, R.N.; Ferreira, V.; Culvenor, R.A.; Hopkins, A.A.; Stewart., A. The first century of Phalaris aquatica L. cultivation and genetic improvement: A review. Crop Pasture Sci. 2009, 60, 1–15. [Google Scholar] [CrossRef]
- Rebecca, B.; Michelle, D.; Luke, P.; Sukhjiwan, K.; Richard, C.; Kevin, S.; German, S.; John, F.; Noel, C. Generation and Characterisation of a Reference Transcriptome for Phalaris (Phalaris aquatica L.). Agronomy 2017, 7, 14. [Google Scholar]
- Li, P.; Yan, J.J.; Bai, S.Q.; Chen, Z.H.; You, M.H.; Su, G.P.; Dao, Z.X. A preliminary study on the silage effect of Elymus sibiricus L. and Phalaris aquatica L. in alpine pastures of northwestern Sichuan. Acta Agrestia Sin. 2012, 20, 368–372. [Google Scholar]
- Polechońska, L.; Klink, A. Trace metal bioindication and phytoremediation potentialities of Phalaris arundinacea L. (reed canary grass). J. Geochem. Explor. 2014, 146, 27–33. [Google Scholar] [CrossRef]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J. The use of reed canary grass and giant miscanthus in the phytoremediation of municipal sewage sludge. Environ. Sci. Pollut. Res. 2016, 23, 9505–9517. [Google Scholar] [CrossRef] [PubMed]
- Melts, I.; Ivask, M.; Geetha, M.; Takeuchi, K.; Heinsoo, K. Combining bioenergy and nature conservation: An example in wetlands. Renew. Sustain. Energy Rev. 2019, 111, 293–302. [Google Scholar] [CrossRef]
- Song, J.Z.; Ling, P.P.; Fu, W.G. Effect of water stress and rewatering on the physiological and biochemical characteristics of Phalaris arundinacea. Acta Prataculturae Sin. 2012, 21, 62–69. [Google Scholar]
- Zhang, Y.L.; Nie, W.W.; Ren, X.Z.; Zhu, T.X. Effects of Mixed Saline Stress on the Absorption and Transportation of K+ and Na+ of Two Reed Canary grass. Chin. J. Grassl. 2010, 32, 28–33. [Google Scholar]
- Manfred, K.; Niina, H.; Jim, G.; Paul, C.; John, F.; Krishna, A.S.; Filippo, U.; Tia, V.; Laxmi, P.; Susanne, B. Transcriptome characterization and differentially expressed genes under waterlogging and drought stress in the biomass grasses Phalaris arundinacea and Dactylis glomerata. Ann. Bot.-London 2019, 4, 14. [Google Scholar]
- Kulcheski, F.R.; Marcelino-Guimaraes, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Margis, R. The use of microRNAs as reference genes for quantitative polymerase chain reaction in soybean. Anal. Biochem. 2010, 406, 185–192. [Google Scholar] [CrossRef]
- Wen, L.; Tan, B.; Guo, W.W. Estimating transgene copy number in precocious trifoliate orange by TaqMan real-time PCR. Plant Cell Tissue Organ Cult. 2012, 109, 363–371. [Google Scholar] [CrossRef]
- Bustin, S.A.; Vladimir, B.; Garson, J.A.; Jan, H.; Jim, H.; Mikael, K.; Reinhold, M.; Tania, N.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Qu, R.; Miao, Y.; Cui, Y.; Cao, Y.; Zhou, Y.; Tang, X.; Yang, J.; Wang, F. Selection of reference genes for the quantitative real-time PCR normalization of gene expression in Isatis indigotica fortune. BMC Mol. Biol. 2019, 20, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sun, H.; Zhang, R.; Yang, Q.; Zhang, X. Selection of reference gene from Gracilaria lemaneiformis under temperature stress. J. Appl. Phycol. 2014, 27, 1365–1372. [Google Scholar] [CrossRef]
- Yang, Q.; Yin, J.; Li, G.; Qi, L.; Yang, F.; Wang, R.; Li, G. Reference gene selection for qRT-PCR in Caragana korshinskii Kom. under different stress conditions. Mol. Biol. Rep. 2014, 41, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Legal Med. 2012, 126, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, C.; Lan, H.; Gao, S.; Liu, H.; Jian, L.; Cao, M.; Pan, G.; Rong, T.; Zhang, S. Validation of Potential Reference Genes for qPCR in Maize across Abiotic Stresses, Hormone Treatments, and Tissue Types. PLoS ONE 2014, 9, e95445. [Google Scholar] [CrossRef]
- Wu, T.; Li, W.F.; Zhang, J.H.; Han, S.Y.; Qi, L.W. Stability analysis of reference genes for larch gene expression studies by quantitative real-time PCR. For. Res. 2013, 26, 1–8. [Google Scholar]
- Schmer, M.R.; Vogel, E.K.; Mitchell, R.; Perrin, M.R. Net energy of cellulosic ethanol from switchgrass. Proc. Natl. Acad. Sci. USA 2008, 105, 464–469. [Google Scholar] [CrossRef]
- Jia, J.X.; Xiong, Y.; Xiong, Y.L.; Ji, X.F.; Li, D.X.; Bai, S.Q.; Yan, L.J.; You, M.H.; Ma, X.; Zhang, J.B. Transcriptomic sequencing analysis, development, and validation of EST-SSR markers in reed canary grass. Grass Res. 2023, 3, 17. [Google Scholar] [CrossRef]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 341–3411. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2018, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-Based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Clark, O.; Thomsen, M. Restoring wetlands dominated by Phalaris arundinacea (Reed Canary Grass) with multiple treatments: Haying, spraying, and establishing aggressive competitors. Ecol. Restor. 2020, 38, 6–9. [Google Scholar] [CrossRef]
- Maeda, Y.; Hirano, S.; Yoshiba, M.; Tadano, T. Variations in salt tolerance of reed canarygrass (Phalaris arundinacea L.) plants grown at sites with different degrees of cattle urine contamination (Plant Nutrition). Soil Sci. Plant Nutr. 2006, 52, 13–20. [Google Scholar] [CrossRef]
- Jones, T.A.; Carlson, I.T. Inheritance of the Virescent Trait in Reed Canarygrass. J. Hered. 1988, 79, 218–221. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Zhang, C.; Tian, Y.A.; Lin, H. Physiological and transcriptional responses of Phalaris arundinacea under waterlogging conditions. J. Plant Physiol. 2021, 261, 153428. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Jiang, Y.; Li, Y.; Zhang, H.; Li, R. Reference genes identification for normalization of qPCR under multiple stresses in Hordeum brevisubulatum. Plant Methods 2018, 14, 110. [Google Scholar] [CrossRef]
- Boava, L.P.; Laia, M.L.; Jacob, T.R.; Dabbas, K.M.; Alves, J.F.G.; Ferro, J.A.; Ferro, M.I.; Furtado, E.L. Selection of endogenous genes for gene expression studies in Eucalyptus under biotic (Puccinia psidii) and abiotic (acibenzolar-S-methyl) stresses using RT-qPCR. BMC Res. Notes 2010, 3, 43–52. [Google Scholar] [CrossRef]
- Andrade, L.M.D.; Brito, M.D.S. Reference genes for normalization of qPCR assays in sugarcane plants under water deficit. Plant Methods 2017, 13, 28. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Scheible, C. Eleven golden rules of quantitative RT-PCR. Plant Cell Online 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, K.P.; Lledias, F.; Kisselev, A.F.; Scaglione, K.M.; Skowyra, D.; Gygi, S.P.; Goldberg, A.L. Certain Pairs of Ubiquitin-conjugating Enzymes (E2s) and Ubiquitin-Protein Ligases (E3s) Synthesize Nondegradable Forked Ubiquitin Chains Containing All Possible Isopeptide Linkages. J. Biol. Chem. 2007, 282, 17375. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, R.; Kim, D.G.; Lee, B.D.; Hwang, S. Over-expression of tobacco UBC1 encoding a ubiquitin-conjugating enzyme increases cadmium tolerance by activating the 20S/26S proteasome and by decreasing Cd accumulation and oxidative stress in tobacco (Nicotiana tabacum). Plant Mol. Biol. 2017, 94, 433–451. [Google Scholar] [CrossRef]
- Chung, E.; Cho, C.W.; So, H.A.; Kang, J.S.; Chung, Y.S.; Lee, J.H. Overexpression of vrubc1, a mung bean e2 ubiquitin-conjugating enzyme, enhances osmotic stress tolerance in Arabidopsis. PLoS ONE 2013, 8, e66056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.R.; Feng, Y.Y.; Yang, M.J.; Xiao, Y.; Liu, Y.S.; Yuan, Y.; Li, Z.; Zhang, Y.; Zhuo, M.; Zhang, J. Systematic screening, and validation of reliable reference genes for qRT-PCR analysis in Okra (Abelmoschus esculentus). Sci. Rep. 2022, 12, 12913. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, H.; He, Z.; Wang, C.; Wang, Y.; Ji, X. Selection of appropriate reference genes for quantitative real-time reverse transcription PCR in Betula platyphylla under salt and osmotic stress conditions. PLoS ONE 2019, 14, e225926. [Google Scholar] [CrossRef]
- Wang, X.; Kong, X.; Liu, S.; Huang, H.; Xu, Y. Selection of Reference Genes for Quantitative Real-Time PCR in Chrysoperla nipponensis (Neuroptera: Chrysopidae) Under Tissues in Reproduction and Diapause. J. Insect Sci. 2020, 20, 20. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, N.; Si, H.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef]
- Li, Y.; Deng, Y.; Li, Z.; Liu, Z.; Cui, X. Composition, physicochemical properties, and anti-fatigue activity of water-soluble okra (Abelmoschus esculentus) stem pectins. Int. J. Biol. Macromol. 2020, 165, 2630–2639. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Du, H.; Jin, Q.; Lin, Y.; Zhong, Y.; Yao, C.; Xu, X.; Wu, T. Stability determination of candidate reference genes in cucumber plants subjected to stresses from Phytophthora melonis. Height. Sci. Publ. Corp. 2019, 3, 28–35. [Google Scholar]
- Jiang, C.X.H.L. Genome-wide identification of plasma membrane aquaporin gene family in Populus and functional identification of PIP-1;1 involved in osmotic stress. Environ. Exp. Bot. 2020, 179, 10. [Google Scholar] [CrossRef]
- Vega, J.; Teshome, A.; Klaas, M.; Grant, J.; Barth, S. Physiological and transcriptional response to drought stress among bioenergy grass Miscanthus species. Biotechnol. Biofuels 2021, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Z.; Fang, S.; Liu, Y.; Shang, X. Integrative analysis of metabolome and transcriptome reveals molecular regulatory mechanism of flavonoid biosynthesis in Cyclocarya paliurus under salt stress. Ind. Crop. Prod. 2021, 170, 113823. [Google Scholar] [CrossRef]
- Yu, R.; Tang, Y.; Liu, C.; Du, X.; Miao, C.; Shi, G. Comparative transcriptomic analysis reveals the roles of ROS scavenging genes in response to cadmium in two pak choi cultivars. Sci. Rep. 2017, 7, 9217. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).